

Fatty_acids_Synthesis
.pdfЛекция № 21.
Тема: «Синтез жирных кислот на синтетазе. Структура и функции полиферментного комплекса синтетазы жирных кислот. Биосинтез триацилглицеринов»
План.
1.Синтез жирных кислот. Строение синтазы жирных кислот.
2.Синтез насыщенных и мононенасыщенных жирных кислот.
3.Синтез триацилглицеридов.
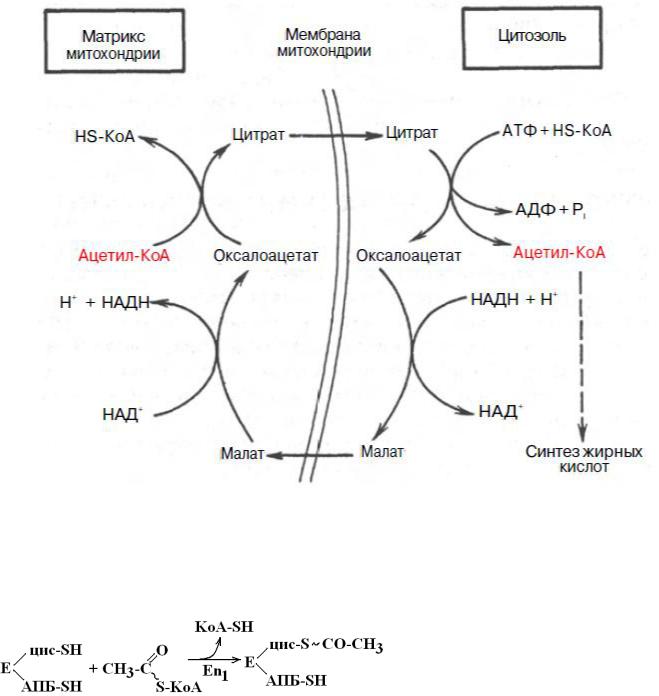
1. Синтез жирных кислот. Строение синтазы жирных кислот. Жирные кислоты, синтезируемые в организме человека, называются заменимыми. Незаменимые жирные кислоты должны проступать в организм с пищей (растительные масла). К незаменимым жирным кислотам относятся полиненасыщенные жирные кислоты: линолевая, линоленовая и арахидоновая. Жирные кислоты синтезируются в цитозоле клеток печени, жировой ткани и лактирующей молочной железе. «Строительным материалом» для синтеза жирных кислот является ацетил-КоА, который может образовываться при окислении углеводов в абсорбтивный период пищеварения. АцетилКоА образуется в матриксе митохондрий в результате окисления жирных кислот и глюкозы. В абсорбтивный период пищеварения в матриксе митохондрий высока концентрация цитрата, АТФ и НАДН и реакции цикла Кребса тормозятся избытком этих продуктов. Для вступления в реакции синтеза жирных кислот ацетил-КоА выходит из митохондрий в цитозоль при участии цитратного целночного механизма (см. наглядный материал).
Первая реакция синтеза жирной кислоты происходит в цитозоле – это реакции карбоксилирования ацетил-КоА под действием фермента ацетил-КоА-карбоксилазы, при участии СО2, биотина и АТФ:
Ацетил-КоА-карбоксилаза – это регуляторный фермент, который активируется цитратом.
Все последующие реакции синтеза жирной кислоты происходят на мультиферментоном комплексе синтазы жирных кислот или пальмитатсинтазе (т.к. этот фермент синтезирует пальмитиновую кислоту). Мультиферментный комплекс построен из центрального ацилпереносящего белка (АПБ) и семи ферментов: 1. Ацетил-трансацилаза; 2. Малонилтрансацилаза; 3. -кетоацил-АПБ-синтетаза; 4. -кетоацил-АПБ-редуктаза; 5. -оксиацил-АПБ- дегидратаза (еноил-АПБ- дегидратаза); 6. Еноил-АПБ-редуктаза; 7. Тиоэстераза (деацилаза).
В молекуле пальмататсинтазы есть 2 функционально важные сульфгидрильные группы (SH): одна SH группа цистеина (расположена в 3-ем ферменте), вторая – SH группа остатка фосфопантотеновой кислоты (производное витамина В3) на АПБ. В состав комплекса входят редуктазы (4-й и 6-й ферменты), в активном центре которых расположен кофермент восстановленный НАДФН. Главным источником НАДФН являются реакции окислительной стадии пентозофосфатного цикла, а дополнительным источником НАДФН – «яблочная» реакция (см. схему цитратного челнока).
Химизм реакции синтеза жирных кислот включает три основных этапа (см. наглядный материал):
1.Загрузку пальматсинтазы ацетильной и малонильной группами;
2.Наращивание цепи жирной кислоты из С2-фрагментов;
3.Повторение этих этапов до образования 16-ти углеродного скелета пальмитиновой кислоты.
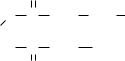
Образовавшийся в результате первого цикла реакций бутирил затем переносится на периферическую SН-группу (образуется бутирил-АПБ), а центральная SH-группа снова загружается малонилом:
O
S C CH2 CH2 CH3
Å
 S C CH2 COOH O
S C CH2 COOH O
Цикл наращивания С2-фрагментов повторяется таким образом семь раз и завершается образованием пальмитиновой кислоты (С15Н31СООН), которая под действием гидролитического фермента деацилазы отщепляется от АПБ-синтетазы.
Энергетика синтеза пальмитиновой кислоты:
Для образования пальмитиновой кислоты на синтазе необходимо: 1. гидролиз семи молекул АТФ для образования семи молекул активного малонила из ацетила-КоА в карбоксилазной реакции.
2. Восстановленный НАДФ·Н для редуктазных реакций на синтазе.
Суммарная реакция синтеза пальмитата на синтетазе:
Ацетил-КоА + 7 Малонил-КоА + 7 АТФ + 14 НАДФН  С15Н31СООН + 7 СО2 + 14 НАДФ
С15Н31СООН + 7 СО2 + 14 НАДФ
Регуляция синтеза жирных кислот
Синтез кислот активируется цитратом, АТФ, малонилом-КоА, а также избытком углеводов, способствующих накоплению в клетке всех этих продуктов. Напротив, конечный продукт синтеза пальмитат ингибирует процесс по принципу обратной связи. Низкая концентрация ЩУК, возникающая при недостатке углеводов, тормозит синтез цитрата и пальмитата, а также усиливает окисление жирных кислот.
2. Синтез насыщенных и мононенасыщенных жирных кислот. Пальмитат является предшественником других длинноцепочечных кислот (стеариновая, лигноцериновая, и др.). Удлинение цепи происходит вне синтетазы путем присоединения С2-фрагментов за счет малонилКоА. Это осуществляют ферменты ЭПС и МХ (жирная кислота из ЦЗ проникает в МХ с помощью карнитина). В свою очередь, пальмитиновая и стеариновая кислоты подвергаются окислению НАДФ-зависимой оксигеназой (фермент ЭПС) с образованием мононенасыщенных пальмитолеиновой и олеиновой кислот (см. наглядный материал). Синтезированные жирные кислоты используются для синтеза триацилглицеридов, глицерофосфолипидов, сфингофосфолипидов и пр. Полиненасыщенные жирные кислоты, а именно арахидоновая (20:4, ω- 6), используются для синтеза эйкозаноидов (простангландинов, тромбоксанов и лейкотриенов) (см. наглядный материал).
3. Синтез триацилглицеридов. (см. наглядный материал). Синтез жиров или триацилглицеринов (ТАГ) происходит в абсорбтивный период в печени и жировой ткани из активных жирных кислот (ацил-КоА) глицеролфосфата. В печени глицеролфосфат образуется в реакции фосфорилирования глицерина под действием фермента глицерокиназы, а в жировой ткани вследствие отсутствия этого фермента глицеролфосфат образуется их метаболита гликолиза диоксиацетонфосфата (см. наглядный материал). Затем к фосфатидной кислоте последовательно присоединяются два ацил-КоА с образованием фосфатидной кислоты. Под действием гидролазы отщепляется остаток фосфорной кислоты и образуется диацилглицерид (ДАГ). В последней реакции синтеза к ДАГ присоединяется еще один остаток ацил-КоА и образуется ТАГ. Синтезированные в печени ТАГ упаковываются в липопротеины очень низкой плотности (ЛОНП) и транспортируются к периферическим тканям, в первую очередь к жировой ткани. Жиры в составе ЛОНП гидролизуются ферментом эндотелия капилляров липопротеинлипазой до глицерина и жирных кислот. Жирные кислоты поступают в адипоциты и используются там для синтеза жира, а глицерин транспортируется в печень, где снова участвует в синтезе ТАГ.
Наглядный материал по теме:
«Синтез жирных кислот на синтетазе. Структура и функции полиферментного комплекса синтетазы жирных кислот. Биосинтез триацилглицеринов»
Транспорт ацетил-КоА из матрикса митохондрий в цитозоль
Биосинтез жирных кислот начинается с транспорта ацетил-КоА из матрикса МХ в
ЦЗ клетки.
Из матрикса МХ цитрат с помощью специального переносчика переносится в ЦЗ. Там под действием цитратлиазы (ЦЗ) происходит обратная реакция.
Синтез жирных кислот
1-ая реакция:

2-я реакция:
3-я реакция:
4-я реакция:
5-я реакция:
6-я реакция:

Синтез заменимых жирных кислот из пальмитиновой кислоты
Характеристика эйкозаноидов
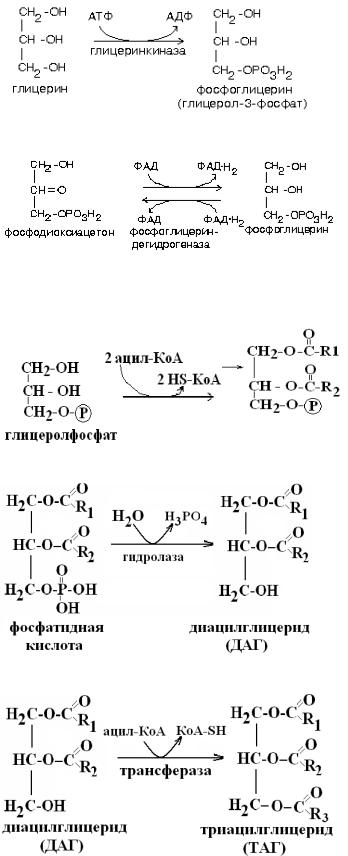
СИНТЕЗ ТАГ
Образование глицеролфосфата В клетках печени, почек, кишечника:
В клетках жировой ткани:
СИНТЕЗ ТАГ
1. Образование фосфатидной кислоты:
2. Синтез диацилглицерида (ДАГ)
3. Синтез ТАГ