

Царство прокариоты
К прокариотам относятся организмы, в цитоплазме которых нет обособленного ядра, а имеется один или несколько участков с высокой концентрацией ДНК. Эти участки называются нуклеотидами или нуклеоплазмой. Нуклеотид более простая форма организации ядерного аппарата. У прокариот он качественно отличается от ядерного аппарата эукариот. У прокариот нет ядрышка, нет высокоорганизованных хромосом, не обнаружены белки гистоны, нет митохондрий, пластид,
ряда других клеточных включений, известных у эукариот. У прокариот в клетках имеются лизосомы - сложные мембранные образования, формирующиеся из цитоплазматических мембран. У некоторых прокариот известны газовые вакуоли, позволяющие им свободно плавать в воде, не
погружаясь на дно.

Рисунок 1. Прокариоты
Основу клеточных стенок у прокариот составляет гликопептид (мукопептид) муреин. У подвижных прокариот более просто, чем у эукариот, устроены жгутики. Клетки прокариот всегда гаплоидны, смены ядерных фаз не наблюдается, у них неизвестны митоз и мейоз. Половой процесс
или отсутствует, или проходит по типу конъюгации. Вследствие удвоения нити ДНК нуклеотида происходит деление клетки на две путем образования поперечной перегородки. Среди представителей встречаются почкующиеся формы, когда от материнской образуется дочерняя,
меньшая чем материнская.
Прокариоты, вероятно, первые организмы Земли. В настоящее время царство прокариот насчитывает около 3000 видов. В их состав включены 2 отдела: дробянки (бактерии, микоплазмы, актиномицеты, спирохеты)и сине-зеленые водоросли (цианобактерии). Большинство прокариот
гетеротрофы, часть из них - паразиты человека, животных, растений и человека. К фототрофным и хемоавтотрофным прокариотам принадлежат пурпурные и зеленые бактерии и сине-зеленые водоросли.
Благодаря ничтожным размерам прокариоты встречаются повсеместно. Они принимают самое активное участие в круговороте веществ в природе. Все органические соединения и значительная часть неорганических подвергаются при этом существенным изменениям. Благодаря их деятельности происходит разложение и минерализация отмерших организмов и их выделений, животных и растительных остатков. В процессе жизнедеятельности прокариот образуется ряд соединений, которые широко используются человеком: антибиотики, аминокислоты, витамины, ферменты и другие соединения. Необычайно велико значение
прокариот в патогенезе заболеваний человека, животных и растений.
Отдел Eubarteriophyta
Cистематика прокариот еще несовершенна. Условно их относят к царству растений (Plantae), однако их связи с высшими растениями или водорослями не доказаны. Среди прокариот наиболее широко распространенная группа микроорганизмов в природе - бактерии. Бактерии - самый большой и чрезвычайно разнообразный мир микроскопических существ. Основные признаки Eubacteria совпадают с характеристикой прокариот.
Название бактерий произошло от греческого слова «бактерион» - палочка. В зависимости от формы клетки бактерии носят разные названия: шаровидные «кокки», палочковидные «бациллы», имеющие форму запятой - вибрионы; спирально закрученные - спириллы, кокки, располагающиеся попарно, - диплококки; цепочкой - стрептококки, собранные в гроздь - стафиллококки.

Рисунок 2
Клетки бактерий имеют плотную
оболочку. У многих вокруг клеток
образуются слизистые капсулы. Многие
бактерии подвижны благодаря очень
тонким жгутикам. Некоторые бактерии
перемещаются, выбрасывая слизь.
Большинство бактерий бесцветно, но
некоторые выделяют пигмент в окружающую
среду, окрашивая ее в желтый, оранжевый,
красный, зеленый, оливково-зеленый цвет.
Некоторые способны образовывать внутри
клетки эндогенные споры и микроцисты
образующиеся из целой клетки, способствующие
перенесению неблагоприятных условий.

Физиология обеспечивает им возможность существования практически в любых местообитаниях. Питание разнообразное, среди бактерий встречаются автотрофы - хемосинтетики и гетеротрофы. Хемосинтетики питаются без участия солнечного света за счет энергии окисления различных неорганических соединений и за счет кислорода воздуха (например, серобактерии окисляют сероводород до серы, а серу до серной кислоты). Гетеротрофные бактерии питаются как сапробионты, т.е. обитатели гниющего органического материала; паразиты или симбиоты. Сапробионтное питание известно двух типов. Брожение - распад органических веществ, который не приводит к полному их окислению часто сопровождается выделением газов (спиртовое, молочнокислое, маслянокислое брожение). Гниение - разложение азотистых веществ белковых соединений. Вызывается гниение многими бактериями, причем деятельность одних может сменяться другими, третьими и т.д. Дыхание большинства бактерий - это окисления кислорода различных органических веществ внутри тела. В результате дыхания аэробных
бактерий образуется углекислота. У некоторых реакция окисления не доходит до конечного предела (уксуснокислые бактерии окисляют этиловый спирт только лишь до уксусной кислоты). Анаэробные бактерии расщепляют сложные органические соединения до более простых без
участия свободного кислорода.
Бактерии являются редуцентами. Благодаря их жизнедеятельности в природе происходит минерализация органического вещества до неорганического. Неорганическое вещество оказывается доступным для питания растений. Бактерии играют огромную роль в круговороте веществ в природе и в создании плодородия почвы. Жизнедеятельность бактерий человек использует в промышленном и сельском хозяйстве.
Лекция 2. Общая характеристика цианобактерий (Cyanophyta)
Сегодняшняя лекция будет посвящена общей характеристике цианобактерий и водорослей.
Цианобактерии ранее называли сине-зелеными водорослями, но поскольку это прокариоты, то сейчас их рассматривают в системе бактерий. Поскольку они являются оксигенными фотосинтезирующими организмами, рассматривать мы их будем вместе с водорослями.
Сюда относятся одноклеточные, колониальные и нитчатые водоросли, окрашенные в синевато-зеленый, черно-зеленый или оливково-зеленый цвета. Окраска зависит от пигментов: хлорофилла а, некоторых каротиноидов, фикобилинов (синего фикоцианина и красного фикоэритрина). Пигментация, а также полное отсутствие жгутиковых стадий сближает сине-зеленые водоросли с красными. Оболочки клеток пектиновые, часто ослизняющиеся. Цитоплазма не содержит вакуолей с клеточным соком, в периферической части она темнее - хроматоплазма, бесцветна в центре - центроплазма. В центроплазме локализована ДНК, хотя ядерная оболочка и ядрышко отсутствуют. Запасной продукт - гликоген, волютин, зерна белка цианофицина. У многих, преимущественно планктонных, в протоплазме в виде черных точек или кружков присутствуют полости, наполненные газом - газовые вакуоли. Предположительно, они содержат N и способствуют парению в толще воды. У многих нитчатых видов в разных местах на протяжении нити или в основании ее образуются т.н. гетероцисты - особые округлые клетки с сильно утолщенными стенками и гомогенным клеточным содержимым.
Класс хроококовые (Chrooccophyceae)
Порядок хроококовые (Chrooccales)
Порядок энтофизалиевые (Entophysalidales)
Класс хамесифоновые (Chamesiphonophyceae)
Класс гормогониевые (Hormogoniophyceae)
Порядок стигонемовые (Stigonematales)
Порядок мастигокладовые (Mastigokladales)
Порядок ностоковые (Nostocales)
Порядок осцилаториевые (Oscilatoriales)
Фикохромовые дробянки (Schirophyceae), слизевые водоросли (Myxophyceae).
Строение клетки.
В природе встречаются:
1) виды с более или менее шаровидными клетками, широкоэлипсоидные, груше - и яйцевидные.
2) виды с клетками, сильно вытянутые в одном направлении (удлинённо эллипсоидные, веретеновидные, цилиндрические от коротко цилиндрических и бочонковидных до удлинённо-цилиндрических). Клетки живут отдельно, иногда соединены в колонии или образуют нити. Клетки имеют толстые стенки. Протопласт окружен четырьмя оболочковыми слоями (L1-L4 слои): двухслойная клеточная оболочка покрыта сверху внешней волнистой мембраной, а между протопластом и оболочкой находится ещё и внутренняя клеточная мембрана. В образовании поперечной перегородки между клеток в нитях участвуют только внутренний слой оболочки и внутренняя мембрана; внешняя мембрана и внешний слой оболочки туда не заходят. Основную роль в клеточной оболочке играют пектиновые вещества и слизевые полисахариды, хотя и содержится целлюлоза. У одних видов клеточные оболочки хорошо ослизняются и содержат даже пигменты; у других вокруг клеток образуется специальный слизистый чехол, иногда самостоятельный вокруг каждой клетки, но чаще сливающийся в общий чехол, окружающий группу или весь ряд клеток. У многих сине-зеленых водорослей трихомы окружены настоящими чехлами - влагалищами. Как клеточные, так и настоящие чехлы состоят из тонких переплетающихся волокон. Они могут быть гомогенными или слоистыми: слоистость у нитей с обособленными основаниями и верхушкой бывает параллельной или косой, иногда даже воронкообразной. Настоящие чехлы растут путем наложения новых слоев слизи друг на друга или внедрения новых слоев между старыми. У некоторых ностоковых (Nostoc, Anabaena) клеточные чехлы образуются путем выделения слизи через поры в оболочках. Протопласт сине-зеленых водорослей лишен оформленного ядра и ранее считался диффузным, разделенным лишь на окрашенную периферическую часть - хроматоплазму - и лишенную окраски центральную часть - центроплазму. Однако различными методами микроскопии и цитохимии, а также ультрацентрифугировапием было доказано, что такое разделение может быть только условным. Некоторые авторы выделяют теперь в протопласте сине-зеленых водорослей три составные части: 1) нуклеоплазму; 2) фотосиптетические пластины (ламеллы); 3) рибосомы и другие цитоплазматические гранулы. Но так как нуклеоплазма занимает район центроплазмы, а ламеллы и другие составные части расположены в районе хроматоплазмы, содержащей пигменты, то нельзя считать ошибкой и старое, классическое разграничение (рибосомы встречаются в обеих частях протопласта). Пигменты, сосредоточенные в периферической части протопласта, локализованы в пластинча-тых образованиях - ламеллах, которые располагаются в хроматоплазме по-разному: хаотично, бывают упакованы в гранулы или ориентированы радиально. Подобные системы ламелл теперь нередко называют парахроматофорами. В хроматоплазме, кроме ламелл и рибосом, встречаются еще эктопласты (цианофициновые зерна, состоящие из липопротеидов) и различного рода кристаллы. В зависимости от физиологического состояния и возраста клеток все эти структурные элементы могут сильно изменяться вплоть до полного исчезновения. Центроплазма клеток сине-зеленых водорослей состоит из гиалоплазмы и разнообразных палочек, фибрилл и гранул. Последние представляют собой хроматиновые элементы, которые окрашиваются ядерными красителями. Гиалоплазму и хроматиновые элементы вообще можно считать аналогом ядра, поскольку в этих элементах содержится ДНК; они при делении клеток делятся продольно, и половинки поровну распределяются по дочерним клеткам. Но, в отличие от типичного ядра, в клетках сине-зеленых водорослей вокруг хроматиновых элементов никогда не удается обнаружить ядерной оболочки и ядрышек. Это - ядроподобное образование в клетке, и называют его нуклеоидом. В нем встречаются и рибосомы, содержащие РНК, вакуоли и полифосфатные гранулы. Установлено, что у нитчатых форм между клетками имеются плазмодесмы. Иногда соединены между собой также системы ламелл соседних клеток. Поперечные перегородки в трихоме ни в коем случае нельзя считать кусками мертвого вещества. Это живая составная часть клетки, которая постоянно участвует в ее жизненных процессах подобно перипласту жгутиковых организмов. Протоплазма сине-зеленых водорослей более густая, чем у других групп растений; она неподвижна и очень редко содержит вакуоли, наполненные клеточным соком. Вакуоли появляются только в старых клетках, и возникновение их всегда приводит к гибели клетки. Зато в клетках сине-зеленых водорослей часто встречаются газовые вакуоли (псевдовакуоли). Это полости в протоплазме, наполненные азотом и придающие клетке в проходящем свете микроскопа черно-бурый или почти черный цвет. Встречаются они у некоторых видов почти постоянно, но есть и такие виды, у которых их не обнаруживают. Присутствие или отсутствие их часто считается таксономически важным признаком, но, конечно, еще далеко не все о газовых вакуолях нам известно. Чаще всего встречаются они в клетках у таких видов, которые ведут планктонный образ жизни (представители родов Anabaena, Aphanizomenon, Rivularia, Microcystis и др., рис. 50, 58,1). Нет сомнения в том, что газовые вакуоли у этих водорослей служат своеобразным приспособлением к уменьшению удельного веса, т. е. к улучшению «парения» в толще воды. И все же их наличие совсем необязательно, и даже у таких типичных планктеров, как Microcystis aeruginosa можно наблюдать (особенно осенью) почти полное исчезновение газовых вакуолей. У некоторых видов они появляются и исчезают внезапно, часто по неизвестным причинам. У ностока сливовидного (Nosloc pruniforme), крупные колонии которого всегда живут на дне водоемов, они появляются в природных условиях весной, вскоре после таяния льда. Обычно зеленовато-коричневые колонии приобретают тогда сероватый, иногда даже молочный оттенок и в течение нескольких дней полностью расплываются. Микроскопирование водоросли в этой стадии показывает, что все клетки ностока набиты газовыми вакуолями и стали черновато-коричневыми, похожими на клетки планктонных анабен. В зависимости от условий газовые вакуоли сохраняются до десяти дней, но в конце концов исчезают; начинается образование слизистого чехла вокруг клеток и их интенсивное деление. Каждая нить или даже кусок нити дает начало новому организму (колонии). Подобную картину можно наблюдать и при прорастании спор эпифитных или планктонных видов глеотрихии. Иногда газовые вакуоли появляются только в некоторых клетках трихома, например в меристемальной зоне, где происходит интенсивное деление клеток и могут возникать гормогонии, выходу которых газовые вакуоли каким-то образом помогают. Газовые вакуоли образуются на границе хромато и центроплазмы и по очертаниям совсем неправильны. У некоторых видов, живущих в верхних слоях придонного ила (в сапропеле), в частности у видов осциллатории, крупные газовые вакуоли располагаются в клетках по сторонам поперечных перегородок. Экспериментально установлено, что появление таких вакуолей бывает вызвано уменьшением в среде количества растворенного кислорода, с прибавлением в среду продуктов сероводородного брожения. Можно предполагать, что такие вакуоли возникают в качестве хранилищ или мест отложения газов, которые выделяются при ферментативных процессах, происходящих в клетке. Состав пигментного аппарата у сине-зеленых водорослей очень пестрый, у них найдено около 30 различных внутриклеточных пигментов. Они относятся к четырем группам - к хлорофиллам, каротинам, ксантофиллам и билипротеинам. Из хлорофиллов пока достоверно доказано наличие хлорофилла а; из каротиноидов, и каротинов; из ксантофиллов - эхинеона, зеаксантина, криптоксантина, миксоксантофилла и др., а из билинротеинов - с-фикоцианина, с-фикоэритрина и алло-фикоцианина. Весьма характерно для сине-зеленых водорослей наличие последней группы пигментов (встречающейся еще у багрянок и некоторых криптомонад) и отсутствие хлорофилла b. Последнее еще раз свидетельствует о том, что сине-зеленые водоросли являются древней группой, отделившейся и пошедшей по самостоятельному пути развития еще до возникновения в ходе эволюции хлорофилла b, участие которого в фотохимических реакциях фотосинтеза дает наиболее высокий коэффициент полезного действия. Разнообразием и своеобразным составом фотоассимилирующих пигментных систем объясняется устойчивость сине-зеленых водорослей к воздействию продолжительного затемнения и анаэробиоза. Этим же частично объясняется и существование их в крайних условиях обитания - в пещерах, богатых сероводородом слоях придонного ила, в минеральных источниках. Продуктом фотосинтеза в клетках сине-зеленых водорослей является гликопротеид, который возникает в хроматоплазме и там же отлагается. Гликопротеид похож на гликоген - от раствора иода в йодистом калии он приобретает коричневый цвет. Между фотосинтетическими ламеллами обнаружены полисахаридные зернышки. Цианофициновые зерна во внешнем слое хроматоплазмы состоят из липопротеидов. Волютиновые зерна в центроплазме представляют собой запасные вещества белкового происхождения. В плазме обитателей серных во-доемов появляются зернышки серы. Пестротой пигментного состава можно объяс-нить и разнообразие цвета клеток и трихомов сине-зеленых водорослей. Окраска их варьи-рует от чисто-сине-зеленой до фиолетовой или красноватой, иногда до пурпурной или коричне-вато-красной, от желтой до бледно-голубой или почти черной. Цвет протопласта зависит от систематического положения вида, а также от возраста клеток и условий существования. Очень часто он маскируется цветом слизистых влагалищ или колониальной слизи. Пигменты встречаются и в слизи и придают нитям пли колониям желтый, коричневый, красноватый, фиолетовый или синий оттенок. Цвет слизи, в свою очередь, зависит от экологических условий - от света, химизма и рН среды, от количества влаги в воздухе (у аэрофитов).
Строение нитей. Немногие сине-зеленые водоросли растут в виде отдельных клеток, большинству свойственно образование колоний пли многоклеточных нитей. В свою очередь, нити могут или образовывать ложнопаренхимные колонии, в которых они тесно сомкнуты, а клетки сохраняют физиологическую самостоятельность, или иметь гормогониальное строение, при котором клетки соединены в ряд, слагая так называемый трихом. В трихоме протопласты соседних клеток соединены плазмодесмами. Трихом, окруженный слизистым влагалищем, получил название нити. Нитчатые формы могут быть простые и разветвленные. Ветвление у сине-зеленых водорослей бывает двояким - настоящим и ложным. Настоящим называют такое ветвление, когда боковая ветвь возникает в результате деления одной клетки перпендикулярно к основной нити (порядок Stigonematales). Ложным ветвлением называют образование боковой ветви путем разрыва трихома и прорыва его через влагалище в сторону одним или обоими концами. В первом случае говорят об одиночном, во втором -о двойном (или парном) ложном ветвлении. Ложным ветвлением можно считать и петлеобразное ветвление, характерное для семейства Scytonemataceae, и редко встречающееся V-образное ветвление - результат повторного деления и роста двух соседних клеток трихома в двух взаимно противоположных направлениях по отношению к длинной оси нити. У очень многих нитчатых сине-зеленых водорослей имеются своеобразные клетки, получившие название гетероцист. У них хорошо выражена двухслойная оболочка, а содержимое всегда без ассимиляционных пигментов (бесцветное, голубоватое или желтоватое), газовых вакуолей и зерен запасных веществ. Они образуются из вегетативных клеток в разных местах трихома, в зависимости от систематического положения водоросли: на одном (Rivularia, Calothrix, Gloeotrichia) и обоих (Anabaenopsis, Cylmdrospermum) концах трихома - базалыю и терминально; в трихоме между вегетативными клетками, т. е. интеркалярно (Nostoc, Anabacna, Nodularia) или сбоку трихома - латералыю (у некоторых Sligone-matales). Гетероцисты встречаются поодиночке или по нескольку (2 -10) в ряд. В зависимости от расположения в каждой гетероцисте возникают одна (у терминальных и латеральных гетероцист) или две, изредка даже три (у интеркалярных) пробки, которые с внутренней стороны закупоривают поры между гетероцистой и соседними вегетативными клетками.

Рисунок 3. Гетероцисты
Гетероцисты называют ботанической загадкой. В световом микроскопе они выглядят как будто пустыми, но иногда, к большому удивлению исследователей, они вдруг прорастали, давая начало новым трихомам. При ложном ветвлении и во время разделения нитей трихомы чаще всего разрываются возле гетероцист, как будто ими ограничивается рост трихомов. Благодаря этому их раньше называли пограничными клетками. Нити с базальными и терминальными гетероцистами прикрепляются к субстрату при помощи гетероцист. У некоторых видов с гетероцистами связано образование покоящихся клеток - спор: они располагаются рядом с гетероцистой по одну (у Anabaenopsis raciborskii, СуImdrospermum, Gloeotrichia) или по обе ее стороны (у некоторых Anabaena). He исключено, что гетероцисты являются хранилищами каких-то запасных веществ или энзимов. Все виды сине-зеленых водорослей, способные фиксировать атмосферный азот, имеют гетероцисты.
Размножение. Самым обычным типом размножения у сине-зеленых водорослей является деление клеток надвое. Для одноклеточных форм этот способ единственный; в колониях в нитях он приводит к росту нити или колонии. Трихом образуется тогда, когда делящиеся в одном направлении клетки не отходят друг от друга. При нарушении линейного расположения возникает колония с беспорядочно расположенными клетками. При делении в двух перпендикулярных направлениях в одной плоскости образуется пластинчатая колония с правильным расположением клеток в виде тетрад (Merismopedia). Объемные скопления в виде пакетов возникают в том случае, когда клетки делятся в трех плоскостях (Eucapsis). Представителям некоторых родов (Gloeocapsa, Microcystis) свойственно также быстрое деление с образованием в материнской клетке множества мелких клеток - нанноцитов. Сине-зеленые водоросли размножаются и другими способами - образованием спор (покоящихся клеток), экзо- и эндоспор, гормогониев, гормоспор, гонидиев, кокков и планококков. Одним из самых распространенных видов размножения нитчатых форм является образование гормогониев. Этот способ послужил названием целому классу гормогониевых (Hormogoniophyсеае). Гормогониями принято называть фрагменты трихома, на которые последний распадается. Гормогонии обособляются благодаря отмиранию некоторых некроидальных клеток, затем с помощью выделения слизи они выскальзывают из влагалища (если оно имеется) и, совершая колебательные движения, перемещаются в воде или по субстрату. Каждый гормогоний может дать начало новой особи. Если группа клеток, похожая на гормогоний, одета толстой оболочкой, ее называют гормоспорой (гормоцистои), которая одновременно выполняет функции и размножения, и перенесения неблагоприятных условий. У некоторых видов от таллома отделяются одноклеточные фрагменты, которые названы гон и дням и, кокками или планококками. Гонидии сохраняют слизистую оболочку; кокки лишены ясно выраженных оболочек; планококки тоже голые, но, подобно гормогониям, обладают способностью к актив-ному движению.Причины движения гормогониев, планококков и целых трихомов (у Oscillatoriaceae) далеко еще не выяснены. Они скользят вдоль продольной оси, колеблясь из стороны в сторону, или вращаются вокруг нее. Движущей силой считают выделение слизи, сокращение трихо-мов по направлению продольной оси, сокращения внешней волнистой мембраны, а также электрокинетические явления. Распространенными органами размножения являются споры, особенно у водорослей из порядка Nostocales. Они одноклеточные, обычно крупнее вегетативных клеток и возникают из них, чаще из одной. У представителей некоторых родов (Gloeotrichia, Anabaena) они образуются в результате слияния нескольких вегетативных клеток, и длина таких спор может достигать 0,5 мм. Не исключено, что в процессе такого слияния происходит и рекомбинация, но пока точных данных об этом нет. Споры покрыты толстой, двухслойной оболочкой, внутренний слой которой называют эндоспорием, а наружный - экзоспорием. Оболочки гладкие или усеяны сосочками, бесцветные, желтые или коричневатые. Благодаря толстым оболочкам и физиологическим изменениям в протопласте (накопление запасных веществ, исчезновение ассимиляционных пигментов, иногда увеличение количества цианофнцпповых зерен) споры могут длительное время сохранять жизнеспособность в неблагоприятных условиях п при разнообразных сильных воздействиях (при низких и высоких температурах, при высыхании и сильном облучении). В благоприятных условиях спора прорастает, ее содержимое делится на клетки -- образуются спорогормогонип, оболочка ослизняется, разрывается или открывается крышкой и гормогоний выходит. Эндо- и экзоспоры встречаются главным образом у представителей класса хамесифоновых (Chamaesiphonophyceae). Эндоспоры образуются в увеличенных материнских клетках в большом количестве (свыше ста). Образование их происходит с у к ц е д а н н о (в результате ряда последовательных делений протопласта материнской клетки) или с и м у л ь т а н н о (путем одновременного распадения материнской клетки на многие мелкие клетки). Экзоспоры по мере своего образования отчлеияются от протопласта материнской клетки и выходят наружу. Иногда они не отделяются от материнской клетки, а образуют на ней цепочки (например, у некоторых видов Chamaesiphon). Половое размножение у сине-зеленых водорослей полностью отсутствует.
Способы питания и экология.
Большинство сине-зеленых водорослей способно синтезировать все вещества своей клетки за счет энергии света. Фотосинтетические процессы, происходящие в клетках сине-зеленых водорослей, в своей принципиальной схеме близки процессам, которые совершаются в других хлорофиллсодержащих организмах. Фотоавтотрофный тип питания является для них основным, но не единственным. Кроме настоящего фотосинтеза, сине-зеленые водоросли способны к фоторедукции, фотогетеротрофии, автогетеротрофии, гетероавтотрофии и даже полной гетеротрофии. При наличии в среде органических веществ они используют и их в качестве дополнительных источников энергии. Благодаря способности к смешанному (миксотрофному) питанию они могут быть активными и в крайних для фотоавтотрофной жизни условиях. В подобных местообитаниях почти полностью отсутствует конкуренция, и сине-зеленые водоросли занимают доминирующее место.В условиях плохой освещенности (в пещерах, в глубинных горизонтах водоемов) в клетках сине-зеленых водорослей изменяется пигментный состав. Это явление, получившее название хроматической адаптации, представляет собой приспособительное изменение окраски водорослей под влиянием изменения спектрального состава света за счет увеличения количества пигментов, имеющих окраску, дополнительную к цвету падающих лучей. Изменения окраски клеток (хлорозы) происходят в случае недостатка в среде некоторых компонентов, в присутствии токсических веществ, а также при переходе к гетеротрофному типу питания. Есть среди сине-зеленых водорослей и такая группа видов, подобной которой среди других организмов вообще мало. Эти водоросли способны фиксировать атмосферный азот, и это свойство сочетается у них с фотосинтезом. Сейчас известно уже около ста таких видов. Как уже указывалось, эта способность свойственна только водорослям, имеющим гетероцисты, да и им не всем. Большинство сине-зеленых водорослей азотфиксаторов приурочено к наземным местообитаниям. Их относительная пищевая независимость как фиксаторов атмосферного азота позволяет им заселять необитаемые, без малейших следов почвы, скалы, как это наблюдалось на острове Кракатау в 1883 г.: через три года после извержения вулкана на пепле и туфах были найдены слизистые скопления, состоящие из представителей родов Anabaena, Gloeocapsa, Nostoc, Calothrix, Phormidium и др. Первыми поселенцами острова Сурцей, возникшего в результате извержения подводного вулкана в 1963 г. около южного берега Исландии, были тоже азотфиксаторы. Среди них оказались некоторые широко распространенные планктонные виды, вызывающие «цветение» воды (Anabaena circinalis, A. cylindrica, A. flosaguae, A. lemmermannii, A. scheremetievii, A. spiroides, Anabaenopsis cir-cularis, Gloeotrichia echinulata). Максимальной температурой для существования живой и ассимилирующей клетки считают +65°С, но это не предел для сине-зеленых водорослей (см. очерк о водорослях горячих источников). Такую высокую температуру термофильные сине-зеленые водоросли переносят благодаря своеобразному коллоидному состоянию протоплазмы, которая при высокой температуре очень медленно коагулирует. Самыми распространенными термофилами являются космополиты Mastigocladus laminosus, Phormidium laminosum. Сине-зеленые водоросли способны выдерживать и низкую температуру. Некоторые виды без повреждения хранились в течение недели при температуре жидкого воздуха (-190°С). В природе такой температуры нет, но в Антарктиде при температуре - 83°С были в большом количестве найдены сине-зеленые водоросли (ностоки). В Антарктиде и в высокогорьях, кроме низкой температуры, на водоросли влияет еще и высокая солнечная радиация. Для снижения вредного влияния коротковолнового радиационного излучения сине-зеленые водоросли в ходе эволюции приобрели ряд приспособлений. Важнейшим из них является выделение слизи вокруг клеток. Слизь колоний и слизистые влагалища нитчатых форм являются хорошей защитной обверткой, предохраняющей клетки от высыхания и одновременно действующей как фильтр, устраняющий вредное влияние радиации. В зависимости от интенсивности света в слизи отлагается больше или меньше пигмента, и она окрашивается по всей толще или по слоям. Способность слизи быстро поглощать и длительно удерживать воду позволяет сине-зеленым водорослям нормально вегетировать и в пустынных районах. Слизь поглощает максимальное количество ночной или утренней влаги, колонии набухают, и в клетках начинается ассимиляция. К полудню студенистые колонии или скопления клеток высыхают и превращаются в черные хрустящие корочки. В таком состоянии они держатся до следующей ночи, когда снова начинается поглощение влаги. Для активной жизни им достаточно парообразной воды. Сине-зеленые водоросли встречаются в почве и в напочвенных сообществах, встречаются они и в сырых местообитаниях, а также на коре деревьев, на камнях и т. п. Все эти местообитания часто не постоянно обеспечены влагой и неравномерно освещены (подробнее см. в очерках о наземных и почвенных водорослях). Они водоросли встречаются также в криофильных сообществах - на льдах и на снегу. Фотосинтез возможен только в том случае, когда клетки окружены прослойкой жидкой воды, что и происходит здесь при ярком солнечном освещении снега и льда. Солнечная радиация на ледниках и снежниках очень интенсивна, значительную часть ее составляет коротковолновое излучение, что вызывает у водорослей защитные приспособления. В группу криобионтов входит ряд видов сине-зеленых водорослей, по все же в целом представители этого отдела предпочитают местообитания с повышенной температурой. Сине-зеленые водоросли преобладают в планктоне эвтрофных (богатых питательными веществами) водоемов, где их массовое развитие часто вызывает «цветение» воды. Планктонному образу жизни этих водорослей способствуют газовые вакуоли в клетках. Прижизненные выделения и продукты посмертного разложения у некоторых из этих сине-зеленых водорослей ядовиты. Массовое развитие большинства планктонных сине-зеленых водорослей начинается при высокой температуре, т. е. во второй половине весны, летом и в начале осени. Установлено, что для большинства пресноводных сине-зеленых водорослей температурный оптимум находится около ЗО°С. Есть и исключения. Некоторые виды осциллатории вызывают «цветение» воды подо льдом, т. е. при температуре около 0°С. Бесцветные и сероводородолюбивые виды развиваются в массовом количестве в глубинных слоях озер. Некоторые возбудители «цветения» явно выходят за границы своего ареала благодаря человеческой деятельности. Так, виды рода Anabaenopsis за пределами тропических и субтропических областей долгое время совсем не встречались, но потом были найдены в южных районах умеренного пояса, а несколько лет назад развились уже в Хельсинкской бухте. Подходящая температура и повышенная эвтрофизация (органическое загрязнение) позволили этому организму развиваться в больших количествах и севернее 60-й параллели. «Цветение» воды вообще, а вызванное сине-зелеными водорослями особенно считается стихийным бедствием, так как вода становится почти ни к чему уже не пригодной. При этом значительно увеличиваются вторичное загрязнение и заиление водоема, так как биомасса водорослей в «цветущем» водоеме достигает значительных величин (средняя биомасса- до 200 г/м3, максимальная - до 450-500 г/ж3), а среди сине-зеленых очень мало таких видов, которые употреблялись бы другими организмами в пищу. Отношения между сине-зелеными водорослями и другими организмами. Виды из родов Gloeocapsa, Nostoc, Scytonema, Stigonema, Rivularia и Calothrix являются фикобионтами в лишайниках. Некоторые сине-зеленые водоросли живут в других организмах в качестве ассимиляторов. В воздушных камерах мхов Anthoceros, Blasia живут виды Anabaena и Nostoc. В листьях водяного папоротника Azolla americana обитает Anabaena azollae, в межклетниках Cycas и Zamia - Nostoc punctiforme. Сине-зеленые водоросли встречаются на всех континентах и во всевозможных местообитаниях - в воде и на суше, в пресных и соленых водах, везде и всюду. Многие авторы придерживаются мнения, что все сине-зеленые водоросли убиквисты и космополиты, но это далеко не так. Выше уже говорилось о географическом распространении рода Anabaenopsis. Подробными исследованиями доказано, что даже такой распространенный вид, как Nostoc pruniforme, не космополит. Некоторые роды (например, Nostochopsis, Camptyloncmopsis, Raphidiopsis) целиком приурочены к поясам жаркого или теплого климата, Nostoc flagelliformo - к аридным районам, многие виды рода Chamaesiphon - к холодным и чисто-водным рекам и ручьям горных стран. Отдел сине-зеленых водорослей считают древнейшей группой автотрофных растений на Земле. Примитивное строение клетки, отсутствие полового размножения и жгутиковых стадий - все это серьезные доказательства их древности. По цитологии сине-зеленые сходны с бактериями, а некоторые их пигменты (билипротеины) встречаются и у красных водорослей. Говоря о хозяйственном значении сине-зеленых, на первое место нужно поставить их роль в качестве возбудителей «цветения» воды. Это отрицательная роль. Положительное значение их заключается, прежде всего, в способности усваивать свободный азот. В восточных странах сине-зеленые водоросли используют даже в пищу, а в последние годы некоторые из них нашли дорогу в бассейны массовых культур для индустриального производства органического вещества. Систематика сине-зеленых водорослей еще далека от совершенства. Сравнительная простота морфологии, относительно малое количество ценных с точки зрения систематики признаков и широкая изменчивость некоторых из них, а также разная трактовка одних и тех же признаков привели к тому, что почти все имеющиеся системы в той или иной мере субъективны и далеки от естественной. Нет хорошего, обоснованного разграничения вида как целого и объем вида в разных системах понимается по-разному. Общее количество видов в отделе определяется в 1500-2000. По принятой нами системе отдел сине-зеленых водорослей делится на 3 класса, несколько порядков и много семейств.
Рассмотрим некоторых представителей:
Род хроококк. Клетки довольно крупные, шаровидные, одиночные, или в малоклеточных неправильной формы колониях. Распространены преимущественно на почве и в пресных водоемах. Род носток характеризуется слизистыми или студенистыми колониями разных размеров и формы: от микроскопических мелких до крупных, достигающих величины сливы. Слизь содержит массу известных нитей беспорядочных или радиально расходящихся из центра колонии. Размножение - посредством гормогониев. Ностоки способны фиксировать атмосферный азот. В водоемах северных и умеренных широт распространен самый крупный носток - сливовидный (Nostoc pruniforme).
![]()
Рисунок 4. Носток
Род анабена (Anabaena) представлен обычно одиночными или собранными в неправильные скопления нитями. Нити симметричные, одинаковой ширины на всем протяжении, состоят из округлых или бочонкообразных вегетативных клеток с промежуточные гетероцистами. Нити прямые или изогнутые. Виды анабены встречаются как в бентосе (придонные), так и в планктоне.
Род осциллятория (Oscillatoria). Его виды часто образуют сине-зеленые пленки, покрывающие влажную землю, подводные предметы, или плавают в виде толстых кожистых лепешек на поверхности стоячих водоемов. Длинные нити осциллатории сложены из цилиндрических клеток, совершенно одинаковых, за исключением верхушечных, которые по форме могут несколько отличаться от остальных. Нити обнаруживают своеобразное колебательное движение, сопровождающееся вращением
нити вокруг собственной оси и ее поступательным движением.

Рисунок 5
Лекция 3. Общая характеристика водорослей
Общим признаком водорослей является отсутствие вегетативных органов (корней, стеблей, листьев), проводящей сосудистой системы, типичных для высших растений. (У подавляющего большинства многоклеточных водорослей, кроме того нет тканей). То есть их тело не расчленено на вегетативные органы, его называют таллом или слоевище. Кроме того, репродуктивные органы без защитного слоя стерильных клеток женские половые органы (если они есть) почти всегда одноклеточные, в отличие от многоклеточных половых органов высших растений.
Водоросли – это низшие преимущественно водные оксигенные фотоавтотрофные, т.е. фотосинтезирующие и образующие при этом процессе кислород, организмы, не имеющие истинных тканей и листостебельной организации. Это искусственная гетерогенная группа живых организмов, включающая много эволюционных линий. Сюда также относят большое количество гетеротрофных представителей, близкородственных организмам с фотосинтетическими пигментами.
Общая характеристика водорослей
Существует много различных классификационных систем водорослей. В нашем курсе мы рассмотрим следующие группы низших оксигенных фотосинтезирующих организмов (слайд):
Цианобактерии или сине-зеленые водоросли – Cyanobacteria seu Cyanophyta иногда также Cyanoprokaryota;
Наиболее крупные отделы водорослей
криптофитовые водоросли – Cryptophyta;
динофитовые водоросли – Dinophyta, ранее эти две группы рассматривали как отдел пирофитовые водоросли – Pyrrophyta;
золотистые водоросли – Chrysophyta;
диатомовые водоросли – Bacillariophyta;
желто-зеленые водоросли – Xanthophyta;
бурые водоросли – Phaeophyta;
красные водоросли – Rhodophyta;
эвгленовые водоросли – Euglenophyta;
9) зеленые водоросли – Chlorophyta.
Как видите, названия разных отделов водорослей связаны прежде всего с их цветом. Цвет в значительной степени обусловлен особенностями химического состава, а именно, различным составом и соотношением пигментов. Считают, что разные отделы водорослей произошли от различно окрашенных жгутиковых форм. В ходе эволюции этих отделов можно наблюдать параллелизм в образовании аналогичных типов морфологической организации талломов. Отделы водорослей существенно различаются по химическому составу, в первую очередь составу фотосинтетических пигментов, запасных веществ, веществ, формирующих клеточную стенку. Значительны также различия по ультраструктурной организации клетки, особенно строению клеточных покровов, хроматофоров, жгутиков.
Можно различить несколько хорошо обособленных типов морфологической структуры таллома водорослей. Это главные ступени морфологической дифференциации таллома водорослей при эволюции. Эти ступени повторяются в разных отделах водорослей, что отражает параллелизм эволюции этих групп. До широкого использования данных об ультраструктуре клетки, данных молекулярной биологии разные уровни организации талломов одного отдела служили основным признаком их классов.
Рассмотрим следующие типы морфологической структуры талломов (слайд):
Монадная.
Ризоподиальная (или амебоидная).
Коккоидная.
Пальмеллоидная (или капсальная).
Нитчатая.
Разнонитчатая (или гетеротрихальная).
Пластинчатая.
9. Сифонокладальная.
Харофитная.
Сифональная (или сифоновая).
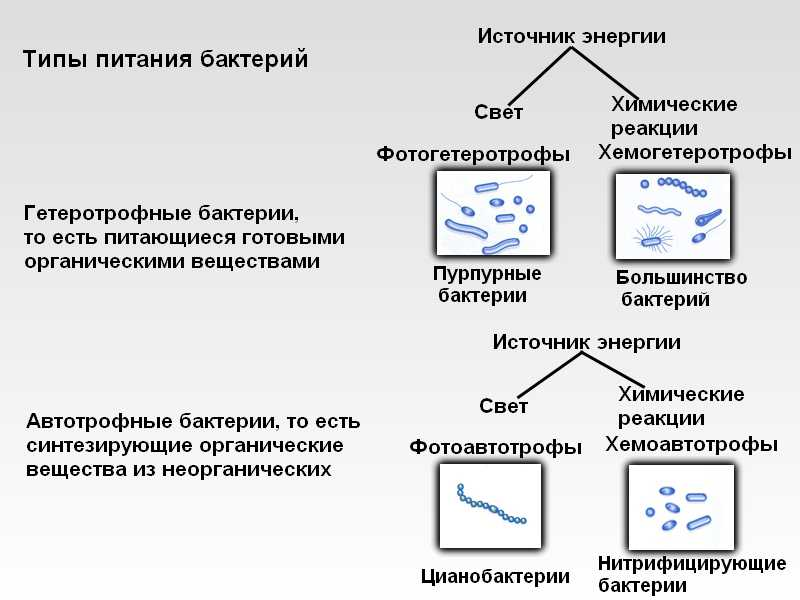
Монадная структура свойственна одноклеточным организмам. Она характеризуется подвижностью с помощью жгутиков (слайд). Эту организацию имеют низшие эволюционные звенья многих отделов водорослей. У вышеорганизованных водорослей монадную форму имеют лишь специализированные клетки, служащие для размножения. Монадная структура свойственна представителям криптофитовых, динофитовых, золотистых, желто-зеленых, эвгленовых и зеленых водорослей. Причем, у эвгленовых, золотистых, криптофитовых и динофитовых она преобладающая.
1 – одиночная клетка Chlamydomonas;
2 – ценобий Pyrobotrys, образованный срастанием клеток;
3 – ценобий Eudorina, образованный слизью.
Ризоподиальная (или амебоидная) структура свойственна одноклеточным организмам (слайд) характеризуется отсутствием твердых оболочек и развитием выростов – псевдоподиев, при помощи которых клетки способны передвигаться. Часто псевдоподии бывают очень длинными и тонкими. Тогда их называют ризоподиями (отсюда и название этого типа организации). Иногда клетки подобного типа соединяются своими ризоподиями, либо формируют плазмодии. Плазмодий – крупное многоядерное вегетативное тело. Амебоидная структура присуща ряду представителей динофитовых, золотистых и желто-зеленых водорослей.

1 – одиночные клетки Chrysamoeba;
2 – колония Chrysidiastrum;
3 – колония Rhizochrysis.
Коккоидная организация представлена неподвижными, окруженными оболочками клетками (слайд 7). Клетки бывают одиночные, либо они объединены в колонии и ценобии (см. ниже). Коккоидная форма – одна из наиболее распространенных, свойственна всем отделам, а у диатомовых она является единственной.

1 – одиночная клетка Chlorococcum;
2 – одиночная клетка Characium;
3 – колония Sphaerocystis, образованная слизью;
4 – ценобий Coelastrum, образованный срастанием клеток.
Пальмеллоидная – Этот тип организации представлен неподвижными клетками, погруженными в общую слизь (слайд). О пальмеллоидной структуре говорят тогда, когда водоросль большую часть жизненного цикла имеет такую форму. Если же водоросли временно образуют подобные слизистые тела, то говорят о пальмеллевидном состоянии. Пальмеллоидный тип структуры таллома распространен у зеленых, желто-зеленых, золотистых водорослей.
Это четыре типа одноклеточных форм водорослей. На основе монадного, амебоидного и коккоидного типов формируются разнообразные колонии, в том числе пальмеллоидный тип морфологической структуры. Клетки в колониях объединены только механически и прямой связи цитоплазмы между отдельными клетками нет.
Если количество клеток колонии определяется на ранних стадиях развития и является постоянным до следующей репродуктивной фазы их называют ценобиями. Ценобий – колония, в которой количество клеток определяется на ранних стадиях развития и не меняется до следующей репродуктивной фазы.

1 – пальмеллоидная структура у золотистой водоросли Hydrurus (часть таллома);
2 – пальмеллевидное состояние у зеленой водоросли Chlamydomonas.
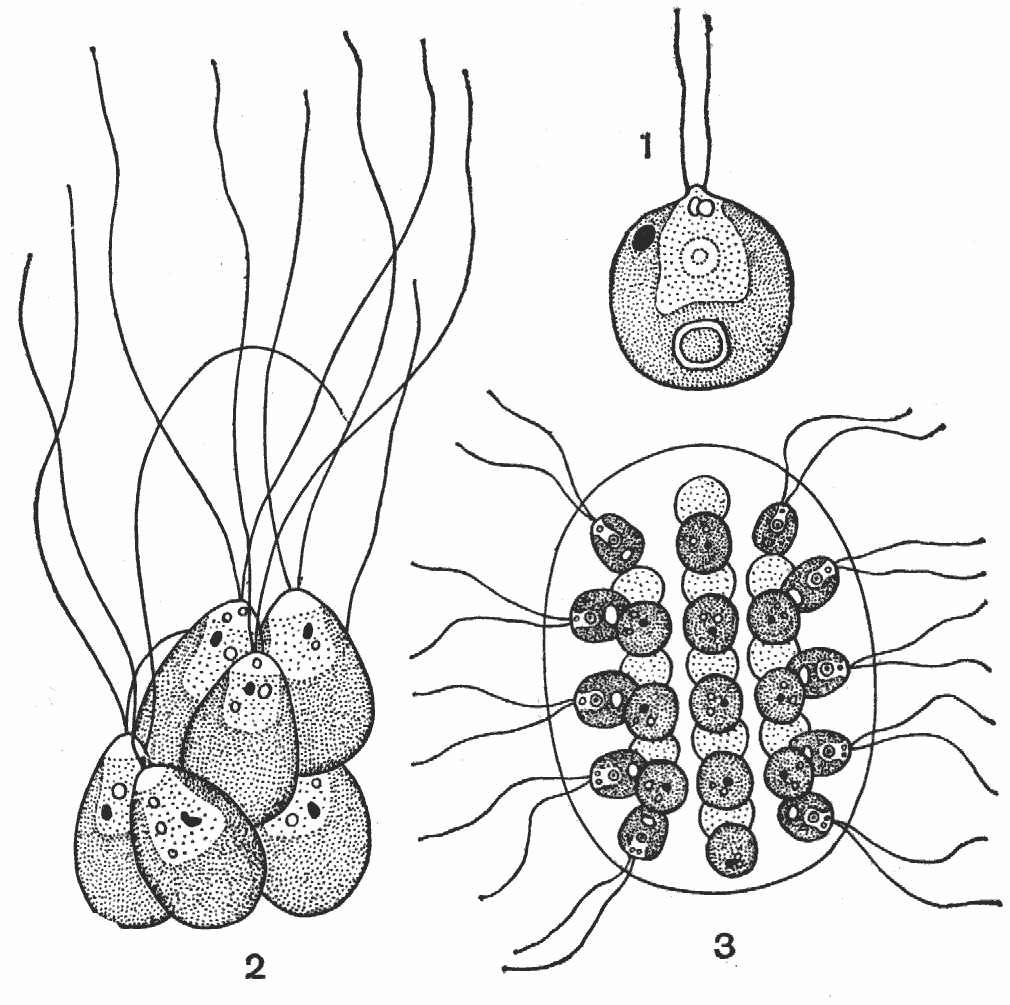
О нитчатой структуре говорит само название (слайд). Нитчатые талломы состоят из нескольких или многих клеток, образуются в результате деления клеток преимущественно в одной плоскости. Нити бывают простые и разветвленные. Клетки нитчатых форм связаны друг с другом порами и плазмодесмами, проходящими через поперечные перегородки. Клетки могут быть полностью одинаковыми на протяжении всей длины нити, так и различаться (морфологически и функционально). Различия наиболее сильно проявляются в прикрепленных организмах, у которых нижняя клетка выполняет прикрепительную функцию и превращается в ризоид. Ризоид – нитевидное корнеподобное образование, прикрепляющее таллом водорослей к субстрату. Ризоиды образуются у прикрепленных водорослей с нитчатым, разнонитчатым, пластинчатым, сифонокладальным, харофитным и сифоновым типом морфологической структуры таллома.
Рост нитчатых талломов может происходить по-разному. Различают диффузный рост (делятся все клетки), апикальный (делятся преимущественно конечные клетки), интеркалярный (зона роста расположена в средней части нити, обычно ближе к вершине) и базальный рост (делятся клетки в основании нити).
Возникновение нитчатого типа структуры было качественно новым этапом в развитии растительного мира. Он послужил отправным пунктом для развития других, более сложных типов структуры.

1 – нить с диффузным ростом у Oscillatoria;
2 – нить с дифференцированным основанием у Endonema;
3, 4 – верхушка нити у Rivularia, вытянутая в волосок;
5 – апикальный рост на концах ветвей у Scytonema;
6 – базальный рост у Calothrix.
Разнонитчатая (или Гетеротрихальная) организация представляет собой нитчатую структуру с морфологически и функционально дифференцированными многоклеточными участками. Чаще всего разнонитчатый таллом состоит из двух частей – стелющейся по субстрату и прямостоящей (слайд). Горизонтальная часть таллома состоит или из различимых нитей, или они полностью смыкаются в сплошную пластинку. В этом случае она нарастает по периферии и уже не имеет следов нитчатого строения. Развитие вертикальной и горизонтальной частей бывает разным, вплоть до редукции какой-либо части. Если редуцируется горизонтальная часть, то таллом становится сходным с нитчатым. Если исчезают вертикальные части, то остается лишь простой однослойный диск, целиком прикрепленный к субстрату, имеющий сходство со следующим, пластинчатым типом строения, но отличающимся по способу формирования.
У некоторых водорослей вертикальные нити дифференцированы на междоузлия и узлы, от которых отходят мутовки боковых ветвей. Этот тип структуры таллома распространен у зеленых, бурых и красных водорослей.
Пластинчатая (или тканевая) структура характеризуется многоклеточными слоевищами в форме пластинок (слайд). Состоят они из одного или нескольких слоев клеток. Такие талломы могут иметь сложную форму.
Образуются такие пластинки из нити в результате продольного деления клеток. Если клетки делятся только в одной плоскости, получается однослойное слоевище, если в двух – 2-многослойное. Разновидность пластинчатой структуры – трубчатое слоевище. Образуется оно из двухслойных талломов в результате расхождения слоев в середине при сохранении связи по краям. При этом внутри образуется полость, окруженная однослойными стенками.
Пластинчатый тип слоевища может ветвиться. Он растет либо распростертый по субстрату, или прикрепленный краем пластинки. Пластинчатые слоевища хорошо представлены в отделах зеленых, бурых и красных водорослей.

Пластинчатая структура у зеленой водоросли Prasiola:
Однорядная нить, разрастающаяся в однослойную пластинку
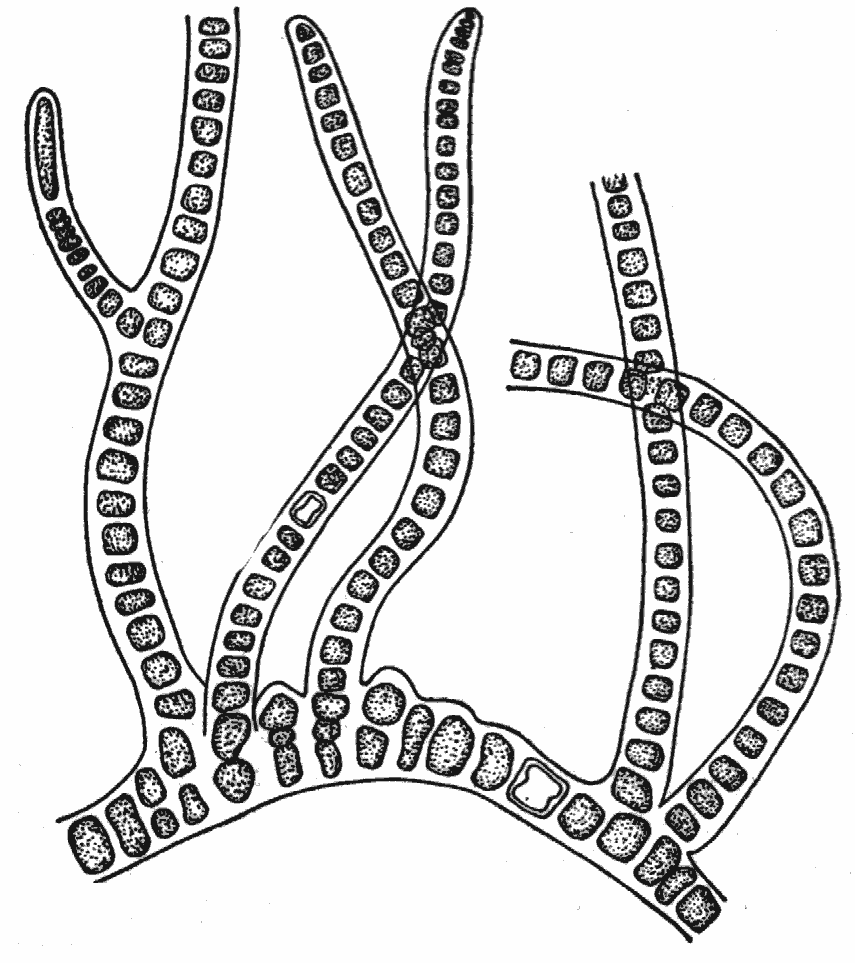
Харофитнаяструктура свойственна только харовым водорослям. Их структура характеризуется наличием главной оси, мутовками членистых боковых осей и отходящими к низу ризоидами. Места расположения мутовок называют узлами, части главной оси между ними – междоузлиями. Междоузлия и членики боковых осей образованы одиночной вытянутой клеткой, но у ряда видов рода хара эти клетки снаружи покрыты слоем других клеток, образующих кору. Такой принцип членисто-мутовчатого строения растительного организма повторяется на более высоком уровне организации у хвощей и некоторых цветковых растений (род казуарина).
Молодой экземпляр Chara
Сифонокладальная форма представлена талломами с многоядерными клетками.
Сифональную (или сифоновую) структуру часто называют неклеточной. Организмы отличаются отсутствием клеточных перегородок и талломы как бы представляют собой одну клетку часто с большим количеством ядер. При этом размеры организма бывают довольно большими и сильно расчлененными внешне. Сифональная структура хорошо выражена в отделах зеленых и желто-зеленых водорослей.
Сифональная структура у морской зеленой водоросли Caulerpa (часть таллома)
Перечисленные ступени организации талломов в разных отделах представлены неодинаково. Наиболее полным спектром форм характеризуются зеленые, золотистые, желтозеленые и динофитовые водоросли. Крайне интересно, что цианобактерии, которые являются прокариотами, также демонстрируют разные типы морфологической структуры таллома. Следует отметить, что у многоклеточных форм, гаметы и споры имеют монадный и коккоидный тип структуры. У высокоорганизованных форм разные стадии развития могут иметь разный тип структуры таллома.
Если расположить некоторые роды водорослей в таблицу, в которой вертикальные ряды отличаются по окраске талломов, а горизонтальные – по их морфологии, то получим сеть наподобие периодической системы элементов Д.И. Менделеева. Этот морфологический параллелизм отражает близкие пути эволюции отдельных групп водорослей. Это фактически классификация водорослей, сложившаяся в 19 - первой половине 20 века. В качестве основных признаков использовали как морфологию (выделяли классы и отделы по горизонтальным рядам), так и пигментацию, проводя разделение по вертикальным столбцам.

Главная причина этого параллелизма – сходство образа жизни и способов питания. Сходный образ жизни обуславливает сходные каналы эволюции. Значительную роль в формировании разнообразия живых организмов сыграл также первичный и вторичный эндосимбиоз, который привел к возникновению качественно иных организмов.
Симбиоз – это различные формы совместного существования разных организмов. Здесь я использую термин симбиоз в узком смысле, соответствующем понятию «мутуализм». Мутуализм – это форма симбиоза, при которой два различных организма возлагают друг на друга регуляцию своих отношений с внешней средой. При этом отношения между партнерами взаимно выгодны и один из них не может существовать без другого. «Далеко зашедший» симбиоз можно рассматривать как особый способ эволюции, при котором сложные организмы образуются из нескольких простых. Теория о происхождении организмов путем симбиоза называется симбиогенез.
Теория эндосимбиоза описывает происхождение клеточных органелл от различных групп прокариот, в частности, пластид от подобных цианобактериям прокариот, живущих в эукариотных клетках. Эндосимбионты в значительной степени редуцированы, но морфологические, биохимические данные, данные молекулярной биологии во многом указывают на прокариотов как на предков пластид. Первичный эндосимбиоз дал начало пластидам с двумя ограничивающими мембранами. Это хлоропласты зеленых водорослей, высших растений, красных водорослей.
В следующем витке эндосимбиоза принимали участие только эукариоты. Красные или зеленые водоросли были проглочены и сохранены другими эукариотами. Таким образом был приобретена возможность фотосинтеза у других эволюционных групп. Это вторичный эндосимбиоз, в результате которого получились пластиды, имеющие три или четыре ограничивающие мембраны. В результате у зеленых и красных водорослей двухслойная оболочка хлоропласта, у динофитовых и эвгленовых – 3-х слойная, у золотистых, желтозеленых, диатомовых, криптофитовых и бурых – четырехмембранная. При вторичном эндосимбиозе захваченная клетка значительно редуцировалась до хроматофора и окружающих мембран, которые возникли как результат захвата. Для динофитовых есть основания предполагать третичный эндосимбиоз. Получается, что клетки некоторых организмов, подобно матрешкам, содержат вставленные друг в друга мелкие клетки.
Вегетативное, бесполое и половое размножение водорослей и их формы вы изучите самостоятельно. Эти вопросы хорошо рассмотрены в «Жизни растений» и «Курсе низших растений».
ЦИКЛЫ РАЗВИТИЯ ВОДОРОСЛЕЙ.
Изменения, претерпеваемые особями вида между одноименными стадиями, составляют его цикл развития. Жизненные циклы в наиболее полном виде включают следующие фазы развития: вегетативный рост, бесполые спороношения, половой процесс, покоящиеся стадии.
В зависимости от места мейоза в жизненном цикле различают следующие его виды:
Типы мейоза:
1. гаметический – гаплоидные гаметы формируются на диплоидном талломе (Fucus и некоторые другие бурые водоросли, диатомовые); характерна для животных. Вся вегетативная фаза диплоидна
2.спорический – гаплоидные споры бесполого размножения формируются на диплоидном талломе (бурая водоросль Laminaria);
3. зиготический – при прорастании зиготы (зеленые водоросли: вольвоксовые, хлорококковые);
4. соматический – гаплоидные вегетативные клетки формируются на диплоидном талломе; впоследствии из них образуются гаметы (зеленая водоросль Prasiola).
Типы жизненных циклов водорослей:
1. Бесполый. Половой процесс отсутствует. Размножение осуществляется гаплоидными спорами Таллом гаплоидный, диплоидной стадии нет (зеленые водоросли: хлорококковые, эвгленовые, криптофитовые и др.)
2. Гаплоидный. На гаплоидном талломе формируются споры и гаметы. После слияния гамет образуются покоящиеся зиготы. Вегетативное тело гаплоидно, диплоидна лишь зигота. Мейоз зиготический. (многие зеленые водоросли).
3. Диплоидный. На диплоидном вегетативном талломе после мейоза формируются гаметы (т.е. это гаметический мейоз). Зигота прорастает в новый диплоидный таллом без периода покоя. Гаплоидны только гаметы (диатомовые водоросли, некоторые бурые водоросли).
4. Гаплоидно-диплоидный. Чередование гаплоидных и диплоидных талломов. На гаплоидном талломе формируются споры и гаметы. Первые прорастают в такой же таллом, вторые, сливаясь дают начало диплоидному таллому. На нем формируются диплоидные споры. Они прорастают в такой же таллом. Кроме того формируются гаплоидные споры после мейоза (спорическая редукция), прорастающие в гаплоидный таллом.
Гаплоидный таллом, формирующий гаметы, называют гаметофитом, диплоидный таллом, образующий споры – спорофит.
В зависимости от от морфологии гаметофита и спорофита может быть несколько типов гаплоидно-диплоидного цикла:
4.1. Изомофный. Гаметофит и спорофит морфологически неразличимы. Наиболее распространен.
4.2.. Гетероморфный. Гаметофит и спорофит морфологически различаются, один из них может быть микроскопическим, другой макроскопическим, либо они оба микроскопические.(бурые водоросли Laminaria, Cutleria; красная водоросль Porphyra).
Смена генераций водорослей позволяет гибко реагировать на изменения условий среды. Гаплоидные особи быстрее размножаются, поэтому для них благоприятны условия, способствующие r-отбору. Диплоидные организмы более устойчивы к неблагоприятным условиям среды, способствующим отбору к-стратегов.
Водоросли благодаря образованию спор и гамет могут наряду с чередованием гаплоидных и диплоидных генераций длительное время существовать только в одной какой-либо фазе. Переход от одной фазы цикла к другой у многих водорослей обусловлен адаптациями к изменившимся условиям среды.
Лекция 4.
Красные водоросли – Rhodophyta
Золотистые водоросли – Chrysophyta
Желто-зеленые водоросли – Xanthophyta