

Сжатая ZIP-папка / ЛК_ген_10
.docЛекция 10
ЛЕКЦИЯ 10
ИЗМЕНЧИВОСТЬ, ЕЁ ПРИЧИНЫ И МЕТОДЫ ИЗУЧЕНИЯ
МНОЖЕСТВЕННЫЙ АЛЛЕЛИЗМ
Проявление одного и того же признака у представителей популяции может носить разный характер. Так, окраска шерсти у животных одного вида может быть белой, чёрной, коричневой, серой и т.д. В этом случае говорят, что один и тот же ген, определяющий признак, может находиться в нескольких состояниях, т.е. в виде разных аллелей. Поэтому, напр., ген А может находиться в виде таких аллелей, как а1, а2, а3 и т.д. Следовательно, рассмотрение любого признака в виде только двух альтернативных проявлений (доминантного и рецессивного, напр., А и а) является несколько упрощённым.
Фактически многие признаки могут иметь несколько аллелей (проявлений). В таком случае говорят о множественном аллелизме. В то же время имеется в виду, что признак должен определяться только одной парой генов.
Поскольку каждый рассматриваемый признак единичен то его обозначают одной и той же буквой алфавита, а отдельные проявления (аллели) обозначают индексами: а1, а2, а3 и т.д.
В целом множественный аллелизм характеризуется следующими положениями:
-
Возникновение таких аллелей возможно при мутациях, в результате чего наблюдаемый признак обладает разнообразным проявлением. Такие взаимопревращения можно изобразить в виде нескольких схем:
b


 1 c 1 c 2
1 c 1 c 2
А

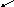

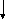

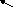
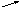
 a, B , c
a, B , c


b 2 c 3
-
Каждый из членов серии может полностью или не полностью доминировать над другим её членом, в результате чего образуются ряды типа A > а1 > а2 > а3 , в которых каждый левый член ряда доминирует над любым членом, находящимся справа от него.
-
Серия множественных аллелей у каждого диплоидного организма может быть одновременно представлена только двумя её членами, т.е. в виде Аа1, Аа2, а1а3 и т.п. Сочетание двух разных или одинаковых членов серии называют КОМПАУНДОМ.
-
Наследование членов серии множественных аллелей подчиняется менделеевским закономерностям.
Одним из практических примеров служит существование и наследование окраски шерсти у кроликов: чёрная, шиншилла, гималайская, неполный альбинос, полный альбинос. Напр., при скрещивании чёрных и гималайских кроликов в первом поколении все будут иметь чёрную окраску, а во втором наблюдаем расщепление 3 : 1. При скрещивании гималайских кроликов с альбиносами в первом поколении все будут иметь окраску «гималайский», а во втором – расщепление 3 : 1. Следовательно, каждая пара указанной серии аллелей ведёт себя как одна аллельная пара. Серия этих аллелей обозначается буквами: С – чёрная, сch сch – шиншилловая окраска, сh сh - гималайская, сa сa - полный альбинос.
Число мутантных аллелей данного гена может быть весьма большим. Напр., по гену окраски глаз у мушки дрозофилы серия аллелей представлена двенадцатью членами, которые в компаунде дают промежуточную окраску от белого до красного.
Множественный аллелизм увеличивает резерв мутационной изменчивости, что важно с эволюционной точки зрения, и обычно имеет приспособительное значение. Однако до сих пор не известно, все ли локусы представлены в виде таких серий множественных аллелей.
Таким образом, изложенный материал свидетельствует о гораздо большей сложности явлений наследственности и изменчивости, чем кажется на первый взгляд.
ВИДЫ ИЗМЕНЧИВОСТИ
Кроме наличия разных качественных проявлений признаков, они всегда характеризуются своей изменчивостью, которая возникает под влиянием различных причин. Изменчивость проявляется как в появлении новых качественных, так и количественных показателей признака.
Различают ненаследственную и наследственную изменчивость.
МОДИФИКАЦИИ
Модификации составляют основу ненаследственной изменчивости, поэтому здесь гораздо труднее выявить какие-либо закономерности. Чарльз Дарвин говорил о неопределённой и определённой изменчивости, что в нынешних терминах соответствует мутационной и модификационной изменчивости, соответственно.
Из повседневной жизни известно, что при пересадке растения в лучшие условия оно увеличивает биомассу, обильнее цветёт и плодоносит. Вспомним также эксперименты по минеральному питанию из курса физиологии растений. Или влияние освещённости на содержание пигментов и фотосинтез у растений.
В природе существует множество примеров модификационных изменений организмов. В зависимости от того, в каких условиях растёт и развивается организм, его фенотипические характеристики могут меняться. Это касается, напр., изменений формы листьев у водяного лютика в зависимости от того, оказываются ли они полностью или нет погруженными в воду, или находятся над водой в воздушной среде. Или наблюдаемое изменение окраски цветков у одного из видов примулы в зависимости от температуры окружающей среды: при 15-20 о С она образует розовые цветки, а при температуре 30-35 о С – белые.
Аналогичные изменения можно наблюдать и у животных. Напр., под воздействием пониженной температуры у млекопитающих волосяной покров становится более густым и пышным.
Такие наблюдения приводят у выводу о том, что любой организм наследует не конкретные по величине и качеству признаки, а только способность формировать определённый фенотип в конкретных условиях среды или эксперимента.
ВАРЬИРОВАНИЕ ПРИЗНАКА И НОРМА РЕАКЦИИ
Для количественной характеристики модификационной изменчивости используют ряд показателей: среднюю арифметическую признака, коэффициент вариации, дисперсию, размах признака и т.д. С некоторыми из них вы познакомитесь на полевой практике.
Здесь же уместно сказать о норме реакции (пределах развития) признака в данных условиях среды или эксперимента. Эти пределы оказываются вовсе не безграничными. Напр., значительная нехватка воды или избыток тепла могут погубить растение и не дать проявиться даже минимальным характеристикам признаков (размеров листьев, корней и т.п.). Напротив, излишние удобрения (особенно азотные) способны не только привести к развитию максимальных показателей признаков, но и также погубить растительный организм. Поэтому норма реакции подразумевает некоторые пределы в развитии признаков, которые позволяют сохранить жизнеспособность организма в заданных условиях.
В то же время модификационной изменчивости придаётся важное эволюционное значение, поскольку модификации позволяют формировать фенотип, максимально соответствующий внешним условиям, и, с другой стороны, соответствующий генетическим возможностям организма.
Впервые строгий количественный учёт к изучению модификационной изменчивости применил В.Иогансен. Семена фасоли массой 150-750 мг (со средней арифметической в 500 мг) он разделил на две группы с массой 250-350 мг и 550-650 мг и высеял отдельно. Из группы «лёгких» семян он получил семена со средней арифметической массы в 440 мг, а из группы «тяжёлых» семян получил новые семена с показателем в 520 мг. В обеих группах наблюдали значительное варьирование массы отдельных семян. Последующие самоопыления и отбор самых тяжёлых и самых лёгких семян в этих линиях в течение шести поколений не позволил вывести линии, которые достоверно различались бы по этому показателю.
Таким образом, вначале сорт был разложен на две «чистые» линии, которые, как оказалось, далее нельзя разделить на какие-либо чётко разграниченные группы.
Из истории науки также известны опыты Вейсмана, который в экспериментах обрубал белым мышам хвосты и скрещивал их между собой в надежде получить бесхвостых или хотя бы короткохвостых мышей.
Подобные опыты дали возможность сформулировать центральную догму генетики о том, что перенос информации возможен только от генетического материала к генным продуктам – белкам, но не в обратном направлении.
Таким образом, ненаследственная (модификационная) изменчивость не сохраняется при половом размножении, связана с изменением фенотипа и обусловлена функционированием генов в процессе формирования признаков в определённых условиях среды. В связи с этим модификационной (ненаследственной) изменчивости в эволюции отведена ограниченная роль, поскольку она отражает вмешательство условий среды в формирование и развитие признака, но не меняет наследственность.
ПРИМЕР: Один и тот же вид, произрастая в разных условиях, даёт разный фенотип. Экстремальные факторы приводят к ещё более резким внешним изменения. И мы говорим о модификационной изменчивости. Однако семена такого растения в нормальных условиях дадут организмы обычно наблюдаемого фенотипа.
Значительный вклад в развитие идей в этом направлении внесли Владимирский, Кирпичников, Шмальгаузен.
Дальнейшее развитие идей в этом направлении привело к формированию такого понятия как «чистая линия», под которым подразумевают серию поколений, полученных от одного самоопыляющегося растения. В более широком понимании – это группа особей, характеризующаяся гомозиготностью по ряду признаков. Такие чистые линии некоторое время остаются генотипически однородными.
Чистые линии могут быть поучены и у животных. Они формируют породы, напр., холмогорскую у коров, орловского рысака – у лошадей и т.п.
Тем не менее, в целом в пределах каждой чистой линии модификации признаков ограничены более узкими характерными пределами изменчивости, которые в индивидуальном конкретном генотипе реализуются в виде нормы реакции на данные условия среды или эксперимента.
В то же время, учёт вариаций признака в разных условиях среды позволяет оценить роль факторов наследственности в формировании признака. Мерой, характеризующей роль факторов наследственности, является наследуемость, которая обозначается буквой h и численно равна квадрату коэффициента корреляции между показателем организма и изменением характеристики внешних условий среды (r 2 ).
Модификационные изменения делят на ряд типов.
Наиболее известны адаптивные модификации, полезные для выживания организма в изменившихся условиях. Напр., увеличение листовых пластинок у растения и более высокое содержание пигментов в листьях в условиях затенения, что позволяет поддерживать фотосинтетические процессы на необходимом уровне. Или появление загара в ответ на облучение солнечным светом кожи человека. Или смена сезонной окраски шерсти у животных или оперения у птиц.
Можно вспомнить и исторические факты, когда монархи употребляли в пищу небольшие количества ядов, и тем самым повышали устойчивость своих организмов к воздействию относительно высоких концентраций ядов, использованных противниками, пытавшимися свергнуть правящую фигуру.
Встречаются модификационные изменения, которые называют МОРФОЗАМИ – изменения фенотипа под воздействием неспецифических внешних факторов. Напр., радиоморфозы – гигантизм у растений, наблюдаемый после облучения (вспомним Чернобыльскую аварию и её последствия). Морфозы не нарушают нормальной жизнедеятельности организма и его взаимодействия со средой, но меняют привычные фенотипические характеристики объекта.
Также выделяют ФЕНОКОПИИ – модификационные изменения, похожие на фенотипическое проявление известных мутаций, но вызванные у нормального в генетическом отношении организма условиями внешней среды.
Обычно модификации нестойки и исчезают, как только прекращается действие фактора, вызвавшего их появление. (Растение затемнение хлорофилла больше яркий свет хлорофилла меньше). НО морфозы и фенокопии отражают вмешательство внешних факторов в развитие признаков на определённой (критической) стадии онтогенеза. Поэтому многие изменения сохраняются на протяжении всей жизни особи, поскольку индивидуальное развитие необратимо (растение удобрения размеры увеличиваются снижение количества удобрений фенотип (размеры растения) не меняется).
Изучение модификаций требует соблюдения ряда правил, главным из которых является генотипическая однородность исследуемого материала, т.е. необходимо использовать в эксперименте только «чистые линии» организмов. Использование же методов статистического анализа позволит говорить о достоверности выявленных закономерностей.
В то же время адаптивная модификация играет важную роль в эволюционном процессе, поскольку позволяет организму выжить и оставить потомство. А возникающие мутантные формы, фенотипически копирующие такие модификации, позволяют увеличить приспособленность организма к изменившимся условиям при участии естественного отбора.
ПРИЧИНЫ МОДИФИКАЦИЙ
Причины модификаций обусловлены изменениями механизмов регуляции действия генов, вызванных влиянием условий внешней среды. Напр., выращивание мух дрозофил при повышенной температуре сопровождается изменением уровня транскрипции генетического материала. Важно отметить, что важнейшая роль в том процессе принадлежит цитоплазме клеток.
Интересно, что воздействие совершенно различных внешних факторов (напр., повышенной температуры, химических веществ и др.) активирует одни и те же гены, что приводит к появлению в клетках аналогичных белковых молекул определённых иРНК. Вначале это явление было показано при воздействии на организм повышенных температур. Поэтому синтезируемые белки стали называть белками теплового шока (БТШ). При этом через несколько часов после начала воздействия внешнего фактора БТШ могут составлять до 10% всех растворимых белков клетки. К настоящему времени выделено несколько групп БТШ, различающихся по молекулярной массе, и которые являются предметом изучения как биохимиков, так и генетиков, поскольку эти белки связаны с регуляцией активности генетического материала.
Иной механизм возникновения модификаций – это временные ненаследуемые изменения генетического материала, которые затем устраняются системами репарации.
НАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ
Наследственная изменчивость связана с изменениями признаков организма, определяется генотипом и сохраняется в ряду поколений. В основе лежат изменения генотипа – мутационные изменения отдельных генов.
Однако, гораздо более широкое проявление наследственной изменчивости обусловлено комбинацией генов и их взаимодействием, что осуществляется двумя путями:
-
независимым распределением хромосом в мейозе и их случайным сочетанием при оплодотворении,
-
перекрёстом хромосом и рекомбинацией (кроссинговером).
В таких случаях говорят о комбинативной изменчивости, при которой меняются не гены, а их сочетания и характер взаимодействия.
Ещё раз подчеркнём, что комбинативный тип наследственной изменчивости – явление вторичное, поскольку обусловлено лишь изменением комбинаций существующих генов.
Однако объектом действия естественного отбора служат, как мутации генов, так и их комбинации и рекомбинации.
Мутационные изменения обязательно связаны с реорганизацией воспроизводящих структур половых и соматических клеток. Они обязательно воспроизводятся в длительном ряду поколений вне зависимости от условий окружающей среды, поскольку связаны с физическим изменением участков хромосом.
В истории развития науки – наследование «благоприобретённых» признаков (Ламарк, частично – Ч.Дарвин). Однако ТОЛЬКО отбор оценивает приспособительное значение мутаций, направление и темп эволюции. Сам естественный отбор обусловлен многими факторами внутренней и внешней среды.
Термин «мутация» предложен Г. де Фризом. Классическая работа – «Мутационная теория». Основные положения его взглядов:
-
мутация возникает внезапно без всяких переходов;
-
новые формы константны в ряду поколений;
-
мутации не образуют непрерывных рядов изменчивости и не группируются вокруг статистического показателя, называемого «модой» (наиболее часто появляющейся величины характеристики в данной выборке). Они являются качественными изменениями;
-
мутации идут в разных направлениях и могут быть полезными или вредными для организма;
-
выявить мутации можно при анализе определённого (часто – весьма значительного) числа особей;
-
одни и те же мутации могут возникать повторно.
Однако Г. де Фриз противопоставил теорию мутаций теории естественного отбора, т.к. считал, что мутации сразу приводят к образованию новых видов, приспособленных к внешней среде без участия отбора.
Фундаментом для теории мутаций послужили эксперименты, доказавшие дискретность хромосом. Открытия Менделя и Моргана способствовали развитию теории мутаций.
ГОМОЛОГИЧЕСКИЕ РЯДЫ НАСЛЕДСТВЕННОЙ ИЗМЕНЧИВОСТИ
Изучение наследственной изменчивости у разных систематических групп растений позволило Н.И.Вавилову сформулировать закон гомологических рядов, характеризующихся двумя основными положениями:
-
Генетически близкие виды и роды характеризуются сходными рядами наследственной изменчивости с такой правильностью, что, зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем генетически ближе друг к другу роды и виды, тем полнее сходство в рядах их изменчивости.
-
Целые семейства растений в общем характеризуются определённым циклом изменчивости, проходящим через все роды и виды, составляющие семейство.
Этот закон был установлен для таких признаков как окраска зерна, форма стеблей, экологический тип и т.п.
Накопленный экспериментальный материал позволяет теперь говорить и о том, что у родственных видов, имеющих общее происхождение, возникают и сходные мутации, как спонтанные, так и индуцированные. Даже у весьма далёких форм наблюдаются параллельные гомологические ряды мутаций по морфологическим, физиологическим и биохимическим признакам и свойствам. Напр., бесшёрстность и альбинизм у разных млекопитающих, альбинизм и отсутствие перьев у птиц, отсутствие чешуи у рыб и т.п.
При этом гомологические ряды мутационной изменчивости биохимических признаков встречаются и у высших организмов, и у микроорганизмов. В основе этого явления лежит биохимическая общность всей живой природы.
МЕТОДЫ ИЗУЧЕНИЯ МУТАЦИЙ
В связи со значительной генетической гетерогенностью представителей даже одного биологического вида или подвида в любой современной области биологической науки любую экспериментальную работу важно проводить с так называемыми «чистыми линиями». К ним относят специально создаваемые группы организмов, характеризующиеся гомозиготным состоянием по генам, функционирование которых изучает исследователь. Только в таком случае можно следить за мутационным процессом и определять его частоту в зависимости от каких-либо внешних или внутренних условий.
Работа с такими линиями предусматривает изучение значительного количества экспериментальных объектов (клеток или организмов), использование наиболее удобных генетических методов, анализ любых изменений наследуемого признака в связи с его наследуемостью.
В основе работы лежит деление исследуемого материала на контрольные образцы и опытные. Последние подвергают, напр., воздействию какого-либо фактора (действию ультрафиолета, химического вещества и т.п.). По разнице в количестве появившихся мутаций через какой-либо промежуток времени судят о мутагенном действии такого фактора.
УЧЁТ МУТАЦИЙ У МИКРООРГАНИЗМОВ
Наиболее удобными в работе оказались гаплоидные микроорганизмы, у которых понятие гена и признака – синонимы. Каждая клетка при росте и развитии образует отдельную колонию (клон). Поэтому можно следить практически за каждой клеткой. Этот метод позволил Бидлу и Татуму внести значительный вклад в изучение процесса мутагенеза.
Многие микроорганизмы дикого типа являются прототрофами, т.е. способны синтезировать все необходимые им органические соединения, поэтому они способны расти на минимальной питательной среде, содержащей только глюкозу, как источник энергии, и минеральные соли. При воздействии какого-либо фактора в группе клеток могут произойти мутации, и такие клетки будут не способны синтезировать, напр., какую-либо аминокислоту или витамин. Поэтому такие мутантные формы (ауксотрофы) будут расти только на минимальной питательной среде, в которую добавлена такая аминокислота или витамин.
Дж.Ледербергом был разработан «метод отпечатков», который позволил значительно упростить методы выявления мутаций. Суть метода состоит в том, что в чашку Петри с минимальной питательной средой заливают раствор микроорганизмов определённой концентрации и обрабатывают её каким-либо веществом, радиоактивным излучением, ультрафиолетом. Через некоторое время с помощью печатки из бархата делают пересевы таких колоний на новые чашки Петри с минимальной средой и выявляют пятна, где микроорганизмы не развиваются.
Затем из этих участков отбирают микроорганизмы и пересевают их на питательные среды, куда добавлен необходимый метаболит, и так выявляют суть возникшей мутации. Подсчёт числа развившихся колоний и числа клеток в них позволяет определить частоту возникшей мутации в расчёте на одну клетку за одно клеточное деление.
Таким образом, были получены разнообразные линии микроорганизмов, характеризующихся, напр., устойчивостью к тем или иным антибиотикам или ядовитым веществам. У кишечной палочки так были выявлены мутантные формы, нуждающиеся для своего роста и развития в биотине, метионине, лейцине, на устойчивость к азиду натрия, фагу Т-1, на неспособность к ферментации лактозы, галактозы, маннита и т.д.
УЧЁТ МУТАЦИЙ У ДРОЗОФИЛЫ
Учет мутаций у высших организмов затруднен рядом факторов.
-
Главная трудность в определении мутаций состоит в ограниченном числе особей, которые можно исследовать.
-
Многие животные – одноплодные особи, т.е. приносят по одному потомку.
-
Большая продолжительность цикла развития до наступления половозрелости.
-
Большое число хромосом.
-
Часто мутации невозможно отличить от изменений фенотипа, вызванных рекомбинацией участков хромосом или отдельных генов. Особенно это относится к тесно сцепленным между собой генам, которые редко разделяются в процессе кроссинговера. В последнем случае на помощь приходят методы биохимии, позволяющие выделить и провести секвенирование (исследование) структуры самого гена или прямого его продукта – белка.
Наиболее удобный метод разработан для мушки дрозофилы. При изучении воздействия рентгеновских лучей на мутационный процесс у дрозофилы Г.Мёллером были получены летальные мутации, сцепленные с полом, что и определило успех экспериментов для развития теоретических представлений.
Легче всего обнаруживаются доминантные мутации, поскольку они проявляются уже в первом поколении. Рецессивные мутации выявляются только при генетическом анализе ряда поколений. Такую мутацию необходимо перевести в гомозиготное состояние. Эту мутантную линию далее скрещивают с линией-анализатором, у которой маркирована одна или несколько групп сцепления, что позволяет выявить место расположения в хромосоме новой мутации.
Описанный путь позволяет относительно просто изучать мутации только у хорошо изученных в генетическом плане организмов – дрозофилы, кукурузы, некоторых простейших особей и микроорганизмов. Для каждого объекта разработаны специальные методы учёта частоты мутаций, которые нашли отражение в учебниках и специальной литературе.
КЛАССИФИКАЦИЯ МУТАЦИЙ
Ген характеризуется определённой структурой и действием. Поэтому мутации в целом можно классифицировать как по фенотипу, так и по характеру изменений в хромосомах.
Каждый организм характеризуется определённой структурой генотипа, которая сформирована в процессе отбора. Аллели генов, типичные для диких (исходных) форм вида, называют генами дикого типа, а изменённые – мутантными. Принципиальной разницы – нет, т.к. гены дикого типа созданы в процессе эволюции и первоначально также являлись мутантными.
Большинство возникающих мутаций – рецессивно, и их появление приводит к гетерозиготности организмов, которую можно обнаружить посредством генетического анализа. Это важно для существования вида, т.к. в своём большинстве мутации вредны для организма в данных условиях. Рецессивный характер мутантных аллелей позволяет им долгое время сохраняться у особей вида в гетерозиготном состоянии и проявляется в результате комбинативной изменчивости.
В ходе эволюции рецессивный аллель может перейти в доминантное состояние, но такой процесс является длительным и сложным. При этом могут происходить изменения генов и в обратную сторону, в результате чего восстанавливается доминантная аллель дикого (наиболее распространённого) типа. Аллели дикого типа – доминируют.
Поскольку гены определяют последовательную цепь таких процессов как морфологическая, физиологическая и биохимическая дифференциация тканей и клеток, постольку можно выделить соответствующие виды мутаций: морфологические, физиологические или биохимические. При этом напр., морфологические мутации изменяют характер роста и формирования органов у животных или растений, и мы наблюдаем коротконогость у сельскохозяйственных животных, отсутствие крыльев или глаз у насекомых, бесшерстность у млекопитающих, гигантизм, карликовость и т.п.. Иногда происходит не только изменение морфологии органа, но и место его развития (на месте антенн у насекомых образуется нога – мутация aristopedia).
Названия мутаций можно давать и по изменению функции гена(ов), напр., мутации стерильности и фертильности, пигментные мутации, мутации групп крови и т.п.
Условность такой классификации очевидна ввиду того, что в основе любого проявления генов и их мутаций лежат биохимические процессы, которые определяют формирование признаков более высоких уровней. При этом изменение функционирования даже одного гена часто приводит к значительным изменениям в организме, что связано с плейотропным характером действия большинства генов.
В связи с выше изложенным, в научной литературе можно встретиться с самыми разными видами классификаций мутаций в зависимости от того, с изменением чего они связаны:
-
В зависимости от характера изменений генома выделяют геномные (связанные с изменением набора гаплоидного числа хромосом в клетке) и хромосомные (связанные с изменением отдельных хромосом) мутации.
-
По проявлению в гетерозиготе – доминантные и рецессивные.
-
По уклонению от нормы (дикого типа) – прямые и обратные.
-
В зависимости от причины возникновения – спонтанные и индуцированные.
-
По локализации в клетке – ядерные и цитоплазматические.
-
По отношению к возможности наследования – соматические и генеративные.
-
По фенотипическому проявлению – летальные, морфологические, биохимические, поведенческие, по устойчивости к фактору (напр., у микроорганизмов – к антибиотику).