

1 курс / Психология / Диагностика_и_лечение_психических_и_наркологических_расстройств (4)
.pdfполушария (поле 18, зона V2), медиальной височной корой (МТ), зоной V4 с подкорковыми центрами, в т.ч. системой заднего продольного пучка, а также с областямикорыдругогополушариячерезмозолистое тело, авнекоторых случаях через его переднюю комиссуру [1].
Стриарная кора (поле 17 или V1) ретинотопически связана многими выходными волокнами со следующей ассоциированной зоной V2 (поле 18), она представлена в мозгу небольшой полоской коры в 6–8 см (шпорная борозда), которая почтиполностьюогибаетполе17.Анатомическионасостоитизверхнейгубы (cuneus) и нижней губы (gyrus lingvalis), между которыми расположена глубокая борозда. На структуре шпорной борозды, расположенной на медиальной и латеральной поверхности затылочной доли мозга каждого полушария, проецируется изображение внешнегомира.Онопредставлено ввидепроекцийполейзрения по типу 4 квадрантов (дорзального и вентрального). Вероятнее всего, каждое изображение в клетках правой и левой шпорных борозд анализируется, и информация передается вболеевысокиеструктуры коры.Ноеслинасетчаткеправого и левого глаза создаются четко различающиеся изображения, то одно из них перестает восприниматься, т.е. нервная система не может воспринимать очень разные изображения (таким образом, возникает борьба изображений). Не исключено, что именно с этим связаны так называемые «неправильные ответы мозга»
вопытах по зрительным иллюзиям Эббингауза.
Вклинике расположение проекций полей зрения сетчатки в виде 4 квадрантов подтверждается при очаговых заболеваниях этой области: после инсультов, раненийкоры18-гополя,опухолейвозникаютразличныевариантыгомонимных гемианопсий (односторонних, верхнеили нижнеквадрантных) [9].
Следует предполагать, что подобно полю 17 зона V2 состоит минимум из 3 совокупностей,мозаичныйхарактерееполяобразовантремятипамипараллельных полосок, в клетках которых происходят удивительные распределения функций.
В совокупности толстых полосок большинство клеток чувствительно к горизонтальному расположению стимулов, подаваемых на оба глаза, отчасти это может быть связано со стереоскопическим зрением. У второй совокупности
втонких полосках клетки лишены ориентировочной избирательности, и они демонстрируют специфические цветовые реакции. Клетки третьей совокупности
вбледных полосках ориентационно избирательныибольшереагируют наконцы линий, т. е. в целом поле 18 связано со стереоскопическим зрением, цветом и формой.
Проведенные в последней четверти ХХ века исследования способом окрашивания клеток зрительных зон с помощью фермента цитохромоксидаза выявило неизвестныеранееклетки,похожиена«пузырьки»,которыетакибылиназваны.
Оказалось,чтоониимелицветовыепредпочтенияибылиполностьюлишены ориентировочнойизбирательности,номоглиполучатьееотокружающихклеток
свыполнением специфических реакций на стимул (линии, отрезки или концы линий и др.). Все эти клетки имеют рецептивные поля с центром и периферией
иучаствуютвкодированиицвета.Возможно,чтовзонеV2ивышемогутсуще-
399
ствовать обособленные участки пузырьковых и непузырьковых клеток, но неизвестно, где происходит их объединение. Д. Х. Хьюбел высказал даже мнение о присутствии в поле 18 (V2) клеток, отвечающих на стимулы «дальнего и ближнего», а также на равномерное расположение объекта (т. е. нулевую диспарантность). Возможно, что эти клетки биполярного типа и сложные, но к тому же обладаютчувствительностьюкглубине,реагируютнадвижущиесястимулы,иногда на концы линий.
Это утверждение нобелевского лауреата подтвердилось работой Д. О’Кифа, М.-Б.МозериЭ.И.Мозера,получившихв2014годуНобелевскуюпремиюзаот- крытие клеток, составляющих систему позиционирования в мозге. В аннотации к работе авторы назвали эту систему «системой GPS» в мозгу, которая делает возможнойориентацию впространстве идемонстрирует клеточнуюоснову для высших когнитивных функций [10].
В60-х годах прошлого века Д. О. Киф, применив методику вживления электродов в гиппокамп крыс, свободно передвигавшихся по клетке, обнаружил так называемые «клетки места», или «пространственные клетки». Они активировалисьлишьтогда,когдаживотные находилисьвопределенном участкеклетки,получившем название «поле клетки».
В2000 году в лаборатории начали работать супруги Мозер, которые расширили диапазон передвижения крыс в клетке и в 2005 году обнаружили в энториальной коре новый элемент ориентации животных — «клетки координатной сетки». В совокупности эти клетки образовывали шестиугольник, а вместе с точками активности других нейронов целую так называемую «гексагональную сетку», которая покрывает всеокружающее пространство мозга иреализует еговозможности, определяетрасстояние иположениеживотных впространстве. Позже былиоткрытыидругиекомпонентысистемынавигации—«клеткинаправления»
всубикулюме и «краевые клетки» в гиппокампе и гиппокампальной формации. Первыеуказывают,вкакую сторонуповернута головаживотного, а«краевые клетки» —границы территории. Обнаружены также клетки сосмешанной активностью. В других исследованиях подобные клетки были найдены в мозге новорожденных крысят, что подтверждает точку зрения о том, что компоненты пространственной ориентации у млекопитающих закладываются еще до рождения потомства. При этом у мышей и крыс система навигации оказалась двухмерной, а у летучих мышей — трехмерной. При лечении пациентов с эпилепсией
всоответствующих зонах были найдены нейроны, похожие на «клетки места» и «клетки координатной сетки», что было подтверждено экспериментами с маг- нитно-резонансной томографией (МРТ).
Таким образом, приведенные данные свидетельствуют о новом, достаточно оригинальном типе работы нейронов, которые формируют многокомпонентную разветвленную сеть, осуществляющую сложную когнитивную деятельность. Не исключено, что нарушение элементов этой сети является основой пространственных и ориентационных дефектов памяти при ряде заболеваний мозга чело-
века [11].
400
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
Наоснованиипроведенных вначале текущеговекаисследованийпопрограмме функциональной МРТ правая и левая зоны V1 передают визуальную информациюпопервичнымдорзальнымивентральнымзрительнымпутям.Дорзальный путь, начинаясь в зоне V1 (17-е поле), проходит через зону V2 (18-е поле), направляется в зону V6 и в задние отделы теменной доли коры. Он ассоциирован с движением, локализацией и возможностью оценки досягаемости объектов, управлением глаз (возможно, саккад?) и др. Вентральный путь из зоны V1 через V2 направляется в зону V4, далее в зону V8 к вентральной (нижней) части височной доли (зона МТ). Он предполагает возможность распознавания формы и характеристик объекта, а также связан с долговременной памятью. Зона V4 отвечает за восприятиецветовыххарактеристикстимулов,высоты,ширины,общегоразмера и ориентации объекта. Зона V8 активируется только при восприятии цвета. Анатомически вышеуказанные зоны вентрального пути примыкают к медиальной поверхности полушария и структурам гиппокампа и так называемой «гиппокампальной формации», которые имеют разветвленные связи с ядрами таламуса, гипоталамуса, ядрами шва и голубым пятном в стволе головного мозга и направляют аксоны в энторинальную кору. По данным В. Ф. Шефера, изучавшего гистологическиеизменениякоры,лимбическойобластиидругихучастковмозга,отвечающих за процессы хранения и воспроизведения различных событий, именно вышеупомянутые структуры демонстрируют самое значительное уменьшение числа корковых клеток, а в мозолистом теле — нервных волокон при прогрессирующихдементирующихзаболеванияхубольныхпожилоговозраста[12].
1.2. Теории цветового зрения
Что такое цвет? Это результат неодинаковой стимуляции фоторецепторов разного типа — палочек и колбочек, в которых содержатся особые вещества — цветовые пигменты.
Теорийцветовоговосприятиякнастоящемувременинасчитываетсяболее30: это и религиозные, и древнегреческие теории; вопросами происхождения цвета в разное время занимались Леонардо да Винчи, поэт Гете и другие выдающиеся деятели.
Одним из первых ученых, заинтересовавшихся цветом, стал Исаак Ньютон, который в 1672 году получил доказательства сложности белого света, расщепив солнечныйлучсистемойпризмиполучивврезультате«всецветарадуги».
Однако первым высказал предположение о современной трехкомпонентной теории цвета М.В.Ломоносов, который на основе своих опытов на публичном собрании Императорской Академии наук 1 июля 1756 года произнес «Слово опроисхождениисвета, новыетеории о цветах представляющее». Ондоказал,что воздействиесветанаглазразлично,ноединопоприроде(«коловратноедвижение эфира»), число основных цветов — три: красный, желтый, голубой, а для создания цветовой гаммы достаточно смешения этих цветов.
Томас Юнг, знакомый с докладом М. В. Ломоносова, в 1802 году объяснил основные положения теории трихроматичности, а Герман Гельмгольц принял
401
и отстаивал теорию Юнга. С тех пор эта теория, носящая название Юнга–Гельм- гольца, является одной из основных теорий о цветовом восприятии мира человеком. В своем «Руководстве по физиологической оптике» Г.Гельмгольцсформулировалгипотезувосприятияитеориюбессознательногоумозаключения.Онписал: «Психическая деятельность, в результате которой мы воспринимаем определенный объект,находящийсяпереднамивопределенномместеиобладающийопределеннымипризнаками,естьвобщемнесознательная,абессознательнаядеятельность. Ее результат эквивалентен заключению, поскольку зарегистрированное воздействие на наши органы чувств позволяет нам сформулировать вывод о возможной причине этого воздействия: ведь фактически воспринимаются только нервные стимулы, а никоим образом не внешние объекты. Заключения астрономов о различных небесных объектах основаны на сознательном применении законов оптики. В обычном акте зрения знание этих законов не фигурирует. И все же позволительно говорить о психических актах обычного восприятия как о бессознательныхзаключениях.Безусловно,были,естьи,можетбыть,навсегдаостанутсянекоторыесомненияотносительностепенисходства междуэтимидвумявидами психической деятельности, но нет никаких сомнений в наличии сходства между результатами, к которым можно прийти с помощью сознательного и бессознательного заключения» (цит. по Р.Л. Грегори)[13].
Вместе с тем, учитывая вклад различных ученых мира в разработку трихроматической теории цвета, следует, вероятно, считать ее творцами М. В. Ломоносова, Т. Юнга, Г. Гельмгольца, Д. Максвела.
В 1870 году была опубликована так называемая оппонентная теория цвета Э. Геринга, известная также как теория обратного процесса. Автор предположил, что в глазу осуществляются оппонентные процессы между парами цветов крас- ный-зеленый, желтый-синий и белый-черный. Он не понимал, почему в теории Юнга–Гельмгольца отсутствуют такие смешанные цвета как желтовато-синий, красновато-зеленый, а также взаимное уничтожение синего и желтого, красного и зеленого, при этом возникает ощущение белого цвета. Автор считал, что красный, зеленый, желтый и синий могут считаться основными цветами, а чернобелый процесс предполагает пространственные сравнения или вычитания отраженных способностей. Его теория позволяет объединить все цвета, в том числе коричневый; при этом работают еще две системы: черно-белая и желто-синяя. Именно восприятие черного, белого и серого цветов осуществляют клетки сетчатки и НКТ с помощью своих рецептивных полей с центрами и периферией. Это подтверждаетсяэкспериментамисоветскогофизиологаА.Л. Ярбуса в1962 году, доказавшего, что если смотреть на синее пятно, окруженное красным фоном, тостабилизацияграницпятнанасетчаткеприэтомприводиткегоисчезновению. Это указывает на то, что для восприятия цвета нужны различия в спектральном составе света из различных участков сетчатки, т. е. это подтверждает гипотезу оналичиивсетчатке клеток,чувствительных кцветовымграницам.Ав1956 году шведско-финско-венесуэльский физиолог Г. Светихин осуществил регистрацию активности нейронов горизонтальных клеток у костистых рыб. Они отвечали
402
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
на стимуляцию сетчатки только медленными потенциалами. Их оказалось три типа: L-клетки гиперполяризовались независимо от спектра световой стимуляции, r-g клетки гиперполяризовались волнами малой длины с максимумом ответа на зеленый цвет и деполяризовались волнами большой длины с максимумом отве- танакрасный;третийтип—y-bклеткиотвечали,какипредыдущие,нос максимумом гиперполяризации на синий цвет и максимумом деполяризации на желтый цвет. Белый цвет у клеток второго и третьего типов вызывал лишь слабые и быстрозатухающие ответы, а свет с промежуточной длиной волны не вызывалникакихреакций,т.е.онисвязанысцветовымиощущениями.В1958 году Р. де Валуа с коллегами обнаружили ответы, сходные со светихинскими, от НКТ макаки. Между тем известно, что цветовое зрение человека и макаки почти одинаковое. Таким образом, де Валуа выявилоппонентно-цветовыеклетки двух типов: красно-зеленые и желто-красные. Д. Хьюбел и Т. Визель считали, что теория Юнга–Гельмгольца верна для рецепторного уровня, а Геринга — для более высокого нейронного уровня. Они же доработали в 70-х годах ХХ века теорию Э. Геринга и высказали предположение, что в мозг поступают сигналы из сетчатки не о трех основных цветах, а только разница их яркости: разница яркости зеленогоикрасного,синегоижелтого,белогоичерного.Приэтомавторыподчеркивали, что яркость зависит не только от полученного, но и от отраженного света [1].
К трихроматической и другим теориям, использующим данные о трех основных цветах и наличии трех типов колбочек, в разное время было высказано много претензий, в частности: в сетчатке всеми научными методами до настоящего времени не обнаружены три типа колбочек, а пигменты, обнаруженные в них, не дают тех смешанных цветов, которые окрашивают объекты полей зрения, до сих пор в колбочках не обнаружен пигмент синего цвета и т.д.
Кроме того, надо учитывать имеющуюся у хрусталика и роговицы особенность:«хроматическаяаберрация»,котораяпредполагаетразложениебелогосвета в спектр таким образом, что синие лучи проецируются ближе к линзе хрусталика, желтого — дальше, а красного — на максимальном удалении от линзы хрусталика.
В 1975 году советский ученый С. Д. Ременко создал нелинейную двухкомпонентную теорию цветового зрения. По его данным в сетчатке находятся два типа фоторецепторов—1типколбочеки1типпалочек.Воднойколбочкеодновре- меннонаходятсякатализаторзеленогоцветахлоралабииндикаторкрасногоцвета эритролаб (они широкополосные и чувствительны ко всей видимой части спектра), а в палочках находится катализатор сине-зеленого цвета — родопсин. Таким образом, существует один тип колбочек, а не три разноцветных, как предполагает трихроматическая теория. При этом прозрачный слой нервных клеток сетчатки позволяет окрашивать красный цвет в пурпурные тона.
Модель цветового восприятия на линейной основе представлена так: свет от источника или отраженный от объекта, или прошедший сквозь него, попадает на роговицу и хрусталик, которые отфильтровывают губительные для сетчатки
403
ультрафиолетовые лучи. Затем свет приходит через стекловидное тело, которое задерживает инфракрасные лучи, содержащие много помех и мало информации. Поэтому фокус для разных участков спектра на различных расстояниях от хрусталикапопадаетнарасположенныенасетчаткеодинаковыеколбочкиипалочки. В результате воздействия света каждая колбочка выдает два сигнала: один являетсяиндикаторомотношенияинтенсивностикрасногоизеленогофронтовсвета, другой — усредненный сигнал, соответствующий желтому фронту. Усредненный сигнал смешиваетсяссиним сигналомотпалочек,создавая вторую цветоразностную пару.Совокупность разностей яркости сигналов красный-зеленый, жел- тый-синий определяет цветовосприятие глаза.
С помощью разработанной модели цветовосприятия С. Д. Ременко объясняет причины всех трех видов дальтонизма; также оказалось технически просто построить прибор колориметр, с помощью которого можно диагностировать многие болезни органа зрения у человека.
Итак, система из одного типа колбочек и одного типа палочек в условиях нелинейности процессов анализа цвета обладает всеми биофизическими свойствами для переработки световой информации и формирования сигналов яркости цветности, которые достаточны для опознания цвета мозгом без дополнительной обработки. Следовательно, можно предположить, что значительная часть цветового восприятия объектов окружающего мира осуществляется уже в клеточных структурах сетчатки. Сам мозг, вероятно, участвует в этом процессе косвенным образом (в какой-то степени это подтверждается различными скоростями прохождения фронтов света и скоростью передачи нервного импульса)
[14].
Весьма интересно предположение Т.Х. Хьюбелла и Т. Визеля о возможном рецепторном и нейронном механизме работы зрительной хиазмы.
Действительно, можно представить, что при однообразной, размеренной работе человека достаточен рецепторный механизм цветового зрения, так как
вэтомслучаефункциимозгазаключаютсявконтроле заработойклетокглазного яблока (сетчатки) и состоянием внешней среды. Но при изменении обстановки для оценки степени опасности необходимо подключение высших аналитических зон, сенсомоторной коры и чувствительных центров. Поэтому задействование нейронногоканала, обеспечивающего, возможно, безопасность живого существа,
вт. ч. человека, является неизбежным.
Не исключено, что в этом процессе важное значение приобретают изменения параметров траектории и скорости так называемой «огибающей» движущейся волны ЭЭГ, связанные с сенсомоторными нейронами [15].
Гипотеза Ременко подтверждается многими отечественными и зарубежными учеными,нотрихроматическаятеорияЮнга–Гельмгольцаостаетсявостребован- ной. Для принятия решения по этому вопросу необходимы, возможно, новые исследования с участием специалистов различных научных школ.
Вероятнее всего, по нервным путям передается яркость цветности объекта поля зрения, а не длина световых волн. Также вероятно, что анализ объектов
404
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
окружающего мира начинается уже в сетчатке и НКТ, а более сложные решения могут приниматься по системе нейронного канала уже в высших ассоциативных зонах. Учитывая многогранный механизм взаимодействия ассоциативных зрительных зон с другими мозговыми центрами, особенно слуховым, речевым, когнитивным и другими, так называемых «зрительных нейронных каналов» может быть достаточно много. Таким образом, из имеющихся к настоящему времени данных о работе зрительного анализатора можно сделать следующие выводы.
1.Природа заложила основы структуры и механизма центрального зрения во взаимодействии с другими органами чувств, и к моменту рождения ребенка ониготовы к функционированию. Последующее увеличение объемазнанийиразвитиеспособностейчеловекаявляетсяоднойизглобальных задачвоспитания иобразования, ответственность закоторыевозлагается на родителей,семью,общество и его просветительские и образовательные структуры.
2.В сетчатке, НКТ и стриарной коре обнаружены клетки с восприятием цвета, поэтому в результате нескольких синаптических переключений анализцветовойгаммыипредметовокружающегомираможетпроисходить уже в сетчатке и НКТ с последующей трансформацией в высших зрительных зонах.
3.Обнаружено, что входные синапсы подкорковых, корковых и более высоких зрительных зон расположены ретинотопически, т. е. соответствуют расположению клеток сетчатки.
4.До настоящего времени неизвестна роль системы нейронной обратной связи, соединяющей клеточные образования высших зрительных зон с клетками сетчатки.
5.Клетки стриарной коры, имеющие огромное число синаптических связей, формируютсявтакназываемые «модули»,реагирующие нацветовые стимулы в виде линий различной длины, конфигурации, ориентации, концы линий, контуры и текстуру различных объектов, обеспечивают бинокулярноезрение,пространственныевзаимодействияидр.Поэтомуструктура зрительной коры может проводить только «локальный» анализ видимой картины окружающего мира.
6.Обобщающий анализ начинается, вероятно, в поле 18 (зона V2) с окончательнымоформлением ввышерасположенных такназываемых «ассоциативных зонах», что подтверждается клиническими данными.
7.Учитывая множественные связи аппарата зрения с другими ассоциативными корковыми зонами, можно предполагать наличие в головном мозге развития сети нейронных зрительных каналов.
8.Обнаружение в гиппокампе, субикулюме, энторинальной коре и др. нейронной сети, определяющей пространственные, ориентационные способности млекопитающих, может свидетельствовать о природе и локализации некоторых когнитивных заболеваний мозгачеловека.
405
II. СОВРЕМЕННЫЕ НЕЙРОВИЗУАЛИЗАЦИОННЫЕ ИССЛЕДОВАНИЯ ЗРИТЕЛЬНОГО ПУТИ
В настоящее время современные методы лучевой диагностики позволяют оценить морфологические изменения практически всех отделов зрительного анализатора. Однако чувствительность и специфичность нейровизуализационных методов отличаются между собой.
2.1. Структурная нейровизуализация
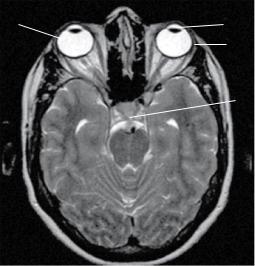
Наиболее информативным методом структурной визуализации зрительного анализатора является МРТ. Так, при исследовании орбит МРТ позволяет с высокой точностью определять патологию их содержимого, включая оболочки зрительного нерва и периневральные ликворные пространства (рис. 1), выявлять процессы, приводящие к компрессии структур хиазмально-селлярной области и являющиеся причиной зрительных расстройств.
2 |
1 |
3
 4 5
4 5
1.Хрусталик
2.Стекловидное тело
3.Слезная железа
4.Зрительный нерв
5.Хиазма
Рис. 1. МРТ головного мозга, Т2-ВИ, аксиальная проекция. Нормальная анатомия зрительного анализатора
2.2. Функциональная нейровизуализация
СпециализированныеметодикиМРТ,такиекакдиффузионно-тензорнаявизу- ализация, позволяют определить микроструктурные изменения в веществе головного мозга, а перфузионная МРТ — установить нарушения мозгового кровотока.
406
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
2.2.1. МР-трактография
Одним из наиболее современных методов оценки состояния зрительного пути являетсяМР-трактография. Этаметодика позволяет оценить диффузию молекул воды вдоль миелиновой оболочки нервных волокон и получить информацию освязях междуразличнымиотделами головногомозга ицелостностипроводящих путей (нервных трактов, пучков нервных волокон).
Данныйметод обеспечивает получение информации о нарушении структуры проводящихволоконбелоговеществапривоспалительных идеструктивныхпроцессах, в том числе в зрительном нерве и зрительном тракте.
При патологических процессах, например при ишемии, воспалении, нейродегенеративных заболеваниях, травме,опухоли,происходитнарушение структуры проводящих путей (отек, дислокация, обрыв тракта).
Одним из достоинств МР-трактографии является также возможность визуализировать нервные структуры в трехмерном изображении.
Сложность задачи состоит в том, что зрительные пути представляют собой сложную и комплексную нейроанатомическую структуру, состоящую как из идущих параллельно, так и пересекающихся пучков нервных волокон, что является критичным при операциях на височной и затылочной долях мозга (БаевА.А.,
2021).
Предлагаемые в настоящее время алгоритмы реконструкции трактов, в том числе при анатомически сложном пересечении трактов, позволяют хорошо визуализировать центральный отрезок зрительного анализатора — от перекреста зрительных нервов до шпорной борозды.
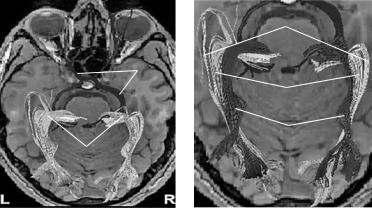
ВыполнениеМР-трактографиизрительныхтрактовпозволяетчетковизуали- зировать все сегменты зрительного тракта (рис. 2).
в
а
г
д
б
А Б Рис. 2. А. Двухполушарные реконструкции зрительного нерва и тракта (а), а также лучистого
венца (б). Б. Разделение лучистого венца на петлю Мейера (в), центральный пучок (г) и спинальный пучок (д).
HoferS.,KarausA.,FrahmJ.Reconstruction and dissection of the entire human visualpathway using diffusion tensor MRI. Front. Neuroanat. 2010; 4: 15. doi: 10.3389/fnana.2010.00015
407
Зрительный тракт представляет собой пучок нервных волокон, который соединяет зрительные нервы и хиазму с латеральными коленчатыми телами. Последние расположенывнижнелатеральных отделахталамусовисодержат третичные нейроны, образующие зрительную лучистость, волокна которой проецируются
всерое вещество шпорной борозды затылочных долей — в область первичной зрительной коры, центральную часть зрительного анализатора. Зрительная лучистость имеет трисегмента:тело(центральный сегмент), вентральный идорсальныйсегменты.Вентральныйсегментсостоитизволокон,располагающихсявглубине средней височной извилины и образующих петлю над крышей височного рога, называемую петлей Мейера, которая, проходя над треугольником и затылочным рогом бокового желудочка, оканчивается внижней шпорной коре. Петля Мейера расположена кзади от передней комиссуры и крючкообразного пучка, на 5 мм кпереди от переднего края височного рога и в среднем на 31 мм кзади от переднего края полюса височной доли. Центральный сегмент зрительной лучистости (тело) проходиткнизуотнижнего продольного пучка и продолжается
впередне-заднемнаправлениидопересечениясослуховойлучистостью,выходя- щей из медиального коленчатого тела. Затем вместе они проходят медиальнее тапетума(частиволоконмозолистоготела),гдеделаютповоротна90.Дорсальный сегмент зрительной лучистости направляется к коре затылочной доли выше или ниже шпорнойборозды (первичная зрительная кора). За долгийпериод, прошедший с момента первой работы, опубликованной A. Meyer в 1907 году, было написано множество статей об анатомии зрительной лучистости, ее переднего, или вентрального сегмента (петли Мейера), а также о зрительных нарушениях после операций на височных долях. Из трех трактов — зрительная лучистость, крючко- образныйтракт,нижнийпродольныйпучок—образуетсяструктура,называемая сагиттальным пластом (stratum). Сагиттальный пласт — конгломерат переплетенных между собой волокон белого вещества, ориентированных в сагиттальной плоскости, гдеволокна идут от височной доли кзатылочной.E.H.Sincoff etal.[17],
U. Türe et al. установили, что волокна задней таламической ножки и нижнего лоб- но-затылочного тракта смешиваются между собой в сагиттальном пласте, что приводит к сложностиих разделения иидентификации, втомчисле определения точного хода волокон зрительной лучистости в их составе. В анатомических работахU.EbelingиH.J.Reulenпоказано,чтопетляМейеравбольшинствеслучаев достигала переднего края височного рога, но в нескольких из 50 препаратов она недоходиладопереднегокраявисочногорога5мм.P.A.Rubinoetal.,используя методКлинглераиприменяяоперационныймикроскопи3D-моделирование фотографий20замороженныхсрезовмозговоговещества,установили,чтоволокна вентральной порции зрительной лучистости, то есть волокна петли Мейера, достигали края височного рога во всех 20 препаратах нормального мозга человека. В каждом из них с помощью операционного микроскопа был выявлен очень тонкий слой передней части петли Мейера, которая во всех наблюдениях распространялась до края височного рога. Во всех исследованных образцах зрительная лучистость полностью покрывала крышу височного рога и была отделена от нее
408
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/