

3 курс / Общая хирургия и оперативная хирургия / Хирургическая_анатомия_венечных_артерий
.pdfПо R. Grant (1926), вены впервые обнаруживаются у эмбрионов длиной 7-8 мм. Н. Bennett (1936), изучавший развитие кровоснабжения сердца у эмбрионов свиньи, первое возникновение вен сердца наблюдал у 6-7-миллиметровых эмбрионов. У эмб риона величиной 2 мм вены распространяются уже у луковицы сердца. Первое появ ление зачатка сердечной вены, по данным ряда авторов (R. Hirakow, 1983; G. Conte and A. Pellegrini, 1984; G. Hutchins et al., 1988 и др.), наблюдается на XVI-XVIII гори зонтах. Он обнаруживается в виде эндотелиального выроста эндокарда, выстилающе
го левый рог |
венозного |
синуса |
(И. И. Новиков, |
1975; Е. Б. Лейтан |
и Л. М. Непомня |
щих, 1979; |
Л.Г.Шелия, |
1985; |
R.Licata, 1955, |
1956; J.Wilson, |
1961; R.Hirakow, |
1983). У 8-миллиметрового эмбриона могут наблюдаться 3-4 таких выроста. Вначале закладываются правая, задняя и средняя вены, затем - левая вена сердца. Правая и левая вены располагаются по предсердно-желудочковой борозде, а средняя растет к верхушке в дорсальной межжелудочковой борозде. Таким образом, эндотелиальные зачатки вен быстро растут и в виде сплетения распространяются в соединительной ткани предсердно-желудочковой борозды, откуда затем проникают в заднюю межже лудочковую и переднюю предсердно-желудочковую борозды. Одновременно в базальной части сердца и в области межжелудочковых борозд появляются протокапилляры с расплывчатыми контурами (Л.Г.Шелия; 1980). Сформированное венозное сплете ние устанавливает связь с синусоидами. На этом этапе замыкается чисто венозный круг кровообращения миокарда. Z. Voboryl и Т. Schiebler (1969) выделяют этот корот кий период в виде отдельной фазы формирования венечного кровообращения - фазы синусоидо-венознои циркуляции. Питание миокарда на этом этапе осуществляется синусоидами, а дренаж - венами и синусоидами.
В дальнейшем венозное сплетение постепенно обособляется из общей венозной се ти и начинает дренировать кровь из синусоид через коронарный синус непосредствен но в правое предсердие.
Как отмечалось, наряду с крупными синусоидами возникают первые эмбриональ ные капилляры, а точнее - протокапилляры (В. В. Куприянов, 1969). В литературе имеются разногласия относительно сроков возникновения капилляров, но то, что их развитие предшествует появлению венечных артерий, поддерживается большинством авторов (R.Grant, 1926; R.Licata, 1956; J.Dbaly, 1964; Z. Voboryl and T. Schiebler, 1969). По поводу источников возникновения протокапилляров в эмбриональном серд це в литературе также существуют разные точки зрения. Так, одни авторы (Б. Н. Клосовский и В.А.Ермакова, 1966; Г.Г.Павлов, 1973; S.Levis, 1904; H.Bennett, 1936; J. Goldsmith and H. Buttler, 1937) считают, что капилляры возникают из венечных со
судов и внедряются в миокард, другие, и |
их большинство (R.Grant, 1926; Z.Voboryl |
and Т. Schiebler, 1969; S. Viragh and E. Challice, |
1981), - придерживаются мнения, что |
капилляры возникают из синусоид, эндотелий которых прорастает в миокард и в субэпикардиальной соединительной ткани образует капилляры. S. Viragh и Е. Challice (1981) помимо этого описывают субэпикардиальные эндотелиальные трубочки, кото рые могут играть роль в становлении капиллярного русла. М. Obrucnik и соавт. (1972), В. Lichnovsky и соавт. (1978) на основании изучения ультраструктуры примордиальных капилляров сердца человека пришли к выводу, что они формируются из трех основных источников: 1) из кровяных островков эпикарда, располагающихся преимущественно в бороздах сердца; 2) из зачатков мезенхимной интерстициальной ткани компактного миокарда; 3) из оставшейся части межтрабекулярных прост ранств губчатого миокарда.
Согласно R. Grant (1926), венечно-артериальное сплетение возникает сначала в виде эндотелиальных тяжей, а затем в них появляется просвет. По данным Е. Б. Лей тан и Л. М. Непомнящих (1979), первые кровеносные сосуды просматриваются под эпикардом в виде скоплений теснолежащих клеток, в центре которых намечается про свет с клеточными элементами крови в них. По мнению W. Hirakow (1983), самые ранние эквиваленты сосудистого сплетения появляются в виде скопления эритробластов, схожих с кровяными островками. Базируются эти скопления, как правило, в бо-
19
роздах сердца. Согласно данным G. Conte и A. Pellegrini (1984), первые признаки со судистой системы сердца зарождаются в виде кровяных островков или капилляров. Эти островки представляют собой эндотелиальные мешки, заполненные ядросодержащими эритроцитами. На первых порах (приблизительно XIV горизонта развития эмб риона) эти островки, или сосудистые тяжи, разобщены, но быстро растут и вследствие бурной пролиферации сосудов соединяются друг с другом, образуя систему каналов, быстро увеличивающуюся в размерах (G.Hutchins et al., 1988). Первичная локализа ция этих островков зафиксирована в межжелудочковых бороздах в области верхушки сердца и по времени совпадает с исчезновением кардиального геля и вследствие этого уменьшением расстояния от эпикарда до эндокарда, что, по мнению G. Hutchins и соавт. (1988), и является причиной образования эпикардиальных кровяных островков.
Так формируется эндотелиальная сосудистая сеть, развивающаяся самостоятель но в субэпикардиальном слое.
После установления сообщений между венами и синусоидами от эпикарда прорас тают капилляры, представляющие собой сначала тяжи, позже в них появляется про свет. На данном этапе развития внутренний слой миокарда состоит из крупных мышеч ных трабекул и синусоид, которые проникают в средний слой и устанавливают связь с капиллярами. Капилляры миокарда, первоначально сравнительно крупные, не имеют определенной направленности и располагаются в компактном миокарде хаотично. В наружном продольном слое капилляры также начинают ориентироваться хаотично.
По мере установления связи с венозной системой сердца (XVI-XVII горизонты) размеры капиллярного сплетения уменьшаются, что, по-видимому, обусловлено дре нированием их содержимого через венечный синус в камеры сердца.
На этом первый этап васкуляризации сердца заканчивается. В этом периоде фор мируются синусоиды. Затем закладывается венозное сплетение и налаживается связь между синусоидами и венами. Данный этап завершается формированием эндотелиальных трубочек, образующих капиллярную сеть.
В норме в растущем сердце по мере увеличения толщины компактного слоя миокар да для него создаются все менее благоприятные условия питания, поскольку диффузия кислорода и питательных веществ через эндотелий синусоид не может удовлетворять потребности плотного миокарда. Согласно данным большинства исследователей
(И.И.Новиков, 1975; Л.Г.Шелия, 1985; J.Dbaly et al., 1968; G.Hutchins et al, 1988),
именно этот фактор и является одной из главных причин появления венечных артерий. По мнению Л. Г. Шелия (1985), второй период васкуляризации сердца начинается с появления венечных артерий. Причем по первоначальному строению стенок отли
чить артерии, вены и капилляры друг от друга трудно. Л. Г. Шелия считает, что арте рии, растущие от центра к периферии, в дальнейшем устанавливают контакт с капил лярным руслом. При этом по мере установления кровотока и развития мышечной обо лочки артерии легко отличимы от вен и капилляров. Коронарная циркуляция на пер вых этапах данного периода функционирует наряду с синусоидной. По мере компактизации миокарда функция кровоснабжения миокарда переходит к венечным артериям.
Третий период характеризуется окончательным формированием терминального русла. В ходе слияния мышечных трабекул (то есть по мере образования компактного миокарда) большая часть синусоид подвергается редукции. Их эндотелиальная вы стилка превращается в эндотелиальный тяж с исчезнувшим просветом. Часть синусо ид сохраняется в виде тонких сосудов, открывающихся в полости желудочков, и, ве роятно, они являются источниками формирования сосудов Вьессена-Тебезия
(И.И.Новиков, 1975; А. К. Габченко, 1980; R.Grant, 1926; H.Bennett, 1936; J. Goldsmith and H. Buttler, 1937; Z. Voboryl and T. Schiebler, 1969 и др.).
Функция кровоснабжения миокарда на данном этапе полностью переходит к ве нечным артериям. По данным А. К. Габченко (1973, 1978, 1980), на ранних этапах эм брионального развития в кровоснабжении сердца человека наблюдается два периода: 1) синусоидный (до 7-10-й недели внутриутробного развития), когда венечные арте рии не развиты и не могут еще питать мышцы сердца, и 2) смешанный, в котором сле-
20
дует различать: а) ранний смешанный (с 9-10-й недели), когда наряду с синусоидами в кровоснабжении сердца принимают участие венечные артерии и притоки венечного синуса с передними венами, и б) поздний смешанный (с 2,5-3-го лунного месяца), во время которого к сердцу подрастают экстракардиальные артерии и вены. При этом, согласно А. К. Габченко, рост и развитие сосудов сердца в ходе эмбриогенеза происхо дит в трех направлениях: 1) со стороны эндокарда растут первичные синусоиды, ко торые образуют интракардиальную (в основном субэндокардиальную) синусоидаль ную сеть; 2) субэпикардиалъно к верхушке сердца растут стволы и ветви венечных ар терий; вены, сопровождающие их, открываются в венечный синус, часть вен открыва ется непосредственно в правое предсердие; 3) в области перикарда по магистральным сосудам сердца к миокарду подрастают экстракардиальные артерии и вены.
Иногда в постнатальной жизни возможно сохранение связи между экстракардиальными сосудами и венечными артериями2 , а также сохранение губчатого миокарда с эмбриональным (синусоидным) типом кровоснабжения3 , что свидетельствует об этапном развитии сердечной стенки и сосудов сердца.
3.Сущность и роль субэпикардиального сплетения
вформировании венечно-артериальной системы
дефинитивного сердца
Как следует из приведенных данных, субэпикардиальная система венечных ар терий образуется из капиллярной сети. Относительно локализации первых зачат ков венечно-артериального сплетения серьезных разногласий в литературе нет. Еще в 1926 г. R. Grant отмечал, что сначала (у эмбрионов длиной 10-11,5 мм) воз никает перитрункальное кольцо с ветвью к передней межжелудочковой борозде. Затем (у эмбрионов длиной 17-20 мм) к ним присоединяются правая и левая огиба ющие ветви. По Н. Bennett (1936), венечные артерии возникают как левая венеч ная артерия с предсердно-желудочковой (огибающей) ветвью с просветом. Чуть позже появляется передняя межжелудочковая ветвь. Первая венечная артерия приобретает просвет позже. Согласно S. Viragh и С. Challice (1981), сначала образу ется сеть в дорсальной межжелудочковой борозде и атриовентрикулярных бороз дах. Далее эти капилляры из атриовентрикулярной борозды переходят на вент ральную поверхность, достигая луковицы сердца. По мнению R. Hirakow (1983), самые ранние сосуды (первоначально в виде скоплений эритробластов, схожих с кровяными островками) обнаруживаются субэпикардиально в области вырезки верхушки сердца, а затем и в межжелудочковых бороздах. По G. Hutchins и соавт. (1988), первичные скопления кровяных островков наблюдаются в межжелудочко-
2В литературе описаны случаи отхождения передней межжелудочковой ветви от внутренней грудной ар терии в сердцах с единственной венечной артерией при тетраде Фалло (W. Evans, 1933; F. Robicsek, 1967), происхождение которых, согласно P. Angelini (1989), по-видимому, обусловлено сохранением связи, су ществующей между сосудами средостения и венечными артериями на ранних стадиях формирования ар териальной системы.
3Известно, что при атрезии легочной артерии с интактной межжелудочковой перегородкой (АЛА с ИМЖП)
впостнатальной жизни сохраняется губчатая структура миокарда с синусоидным типом кровоснабжения
(Blackman et al., 1981; O'Cornor et al., 1982; A. Gittenberger-de Groot et al., 1987). Опорожнение правого же лудочка (ПЖ) при АЛА с ИМЖП происходит через путь с наименьшим сопротивлением. Поскольку трех створчатый клапан при данном пороке компетентный, то единственно возможный путь опорожнения ПЖ - через синусоиды в сохранившемся недифференцированном миокарде (R. Freedom et al., 1983). Другой при
мер сохранения эмбрионального кровообращения - фистулы венечных артерий. По данным A. Gittenberger-de Groot et al. (1987) и A. Bogers et al. (1988), венечно-артериальные свищи представляют собой прямое соединение ВА с камерами сердца (без промежуточного капиллярного русла между ними).
21
вых бороздах. Как следует из материалов, приведенных A. Bogers (1989), венечноартериальное сплетение формируется в атриовентрикулярных бороздах и в перитрункальной области. Согласно данным D. Levin и G. Gardiner (1988), венечно-ар- териальная система формируется к 9-й неделе из ангиобластического скелета. Рас пространяясь в эпикарде, она образует основные стволы ВА. Проксимальная часть сплетения формирует кольцо вокруг трункуса.
Таким образом, несмотря на различия в сроках появления отдельных частей, в це лом во взглядах на образование субэпикардиального сплетения, формирующегося в бороздах сердца, авторы единодушны. Однозначно и суждение авторов об этой систе ме как о предвестнике будущих основных стволов венечных артерий. Неизвестно только, каково строение этой системы, почему она закладывается в определенных мес тах и чем определено постоянство строения венечно-артериальной системы. Между тем именно знание того, как закладывается первичное венечно-артериальное сплете ние и какова его структура, могли бы позволить ответить на вопрос: почему не страда ет кровоснабжение миокарда ни при одном врожденном пороке сердца?
Согласно нашим данным, первичная закладка венечно-артериальной системы (речь идет о ее втором компоненте - субэпикардиальном сплетении) идентична во всех сердцах (рис. 1). Она закладывается в виде постоянной сети колец (сначала в виде скопления ангиобластов) по окружностям соединения сегментов сердца4 . Схематичес ки система этих колец, образующих венечно-артериальное сплетение, однотипна и включает в себя вокругартериальные, вокругбульбарные (перитрункальные), вокругжелудочковые (расположенные в межжелудочковых бороздах) и вокругпредсердные (расположенные в атриовентрикулярных бороздах) кольца (см. рис. 1). Первоначаль но плоскости расположения этих колец находятся в одной плоскости с межсегментар ными соединениями. По мере завершения процесса петлеобразования сердца стороны окружностей коронарогенных зон5 сближаются по внутренней кривизне петли
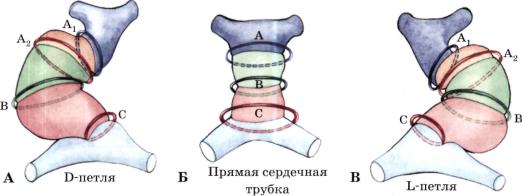
Р и с . 1. Петлеобразование сердца (R. Van Praagh et al., Heart Disease in infants, children and ado lescents. - Baltimore, 1977. - P. 394) и первые этапы ф о р м и р о в а н и я и с б л и ж е н и я венечно-арте- р и а л ь н ы х колец (коронарогенных зон).
При правом типе петлеобразования сердца (А - D-петля) формируется сердце с нормальным пространственным взаи моотношением желудочков, при левом типе (В - L-петля) - желудочки инвертированы. В зависимости от стороны пет леобразования сердца складываются нормальные и инвертированные взаимоотношения между первичными венечноартериальными кольцами. Б - сердце на стадии прямой трубки. А (Ах, А2) - вокругбульбарные венечно-артериальные кольца, В - бульбовентрикулярное кольцо, С - вокругпредсердное кольцо.
4Мы уже отмечали, что, по данным литературы, венечно-артериальное сплетение закладывается в бороз дах сердца. При интерпретации этих данных в свете посегментного строения сердца становится ясно, что борозды в дефинитивном сердце представляют собой зону межсегментного соединения. Понятно, что от дельные кольца будущего венечно-артериального сплетения первоначально возникают по окружностям соединения сегментов формирующегося сердца.
5После появления просвета в тяжах, образованных ангиобластами, коронарогенные зоны превращаются в сосудистые образования, формирующие сплетение.
22
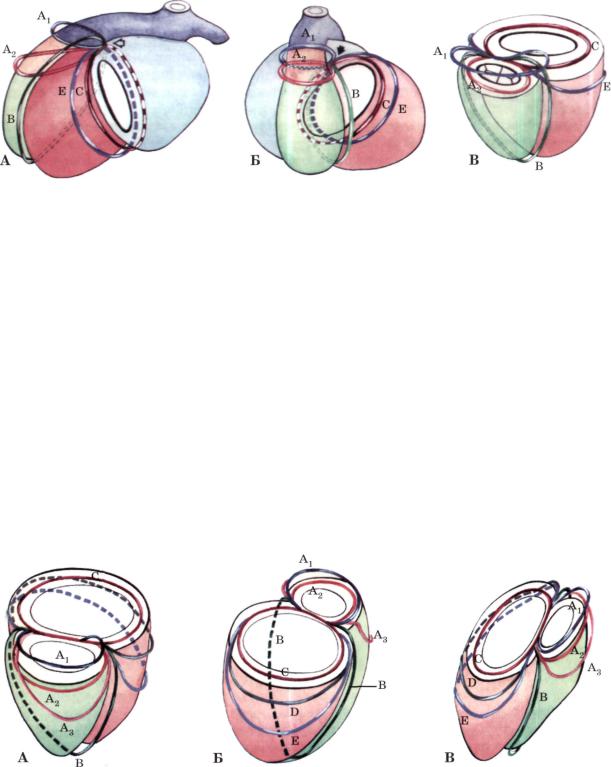
Рис. 2. Соединение венечно - артериальных колец (показано стрелкой) на последующих этапах формирования желудочкового к о м п л е к с а с образованием единого венечно-артериального спле тения.
А - вид на сердце сбоку, Б - вид на сердце спереди, В - вид на сердце спереди после удаления предсердий и трункуса.
Си Е - вокругжелудочковые венечно-артериальные кольца. Обозначения остальных колец те же, что и на рис. 1.
ив конечном счете сливаются (рис. 1,2). После завершения процесса петлеобразова ния сердца, как следует из рис. 3, образуется единая первичная система субэпикардиальной сети ВА, представляющая собой сначала единую, обособленную систему ВА (второй компонент). При этом система субэпикардиальных сосудов - это комплекс из расположенных в виде веера колец, сходящихся в ретробульбарной области6 . В ре зультате такой пространственной переориентации и перестановки колец вокругбульбарное (перитрункальное) кольцо приобретает специфическое положение и особое зна чение: если после этого остальные кольца начинают играть роль коммуникаций меж ду перитрункальным кольцом и миокардом, то от перитрункального кольца начинает зависеть нормальность (и ее тип) или аномальность интеграции в артериальную сис тему. Сохранность центральных концов коммуникационных колец обеспечивает нормальное распределение и следование ВА во всех сердцах (в нормальных и порочно сформированных). Редуцирование же их дистальных фрагментов, связанных с раз личными факторами (процессом роста камер сердца, достаточности центрального
Рис. 3. В з а и м о о т н о ш е н и я венечно - артериальных колец и желудочкового к о м п л е к с а сердца (после удаления предсердий и трункуса - см. рис . 2) на этапе ф о р м и р о в а н и я единого венечноартериального сплетения .
А - вид спереди (впереди бульбарное сплетение — круги Вьессена), Б — вид сзади (впереди желудочковое сплетение), В - вид сбоку, справа. Ар А2 и А3 — вокругбульбарные кольца; С, D и Е - вокругжелудочковые кольца; В — бульбовентрикулярное кольцо.
6 Неполное слияние этих колец в ретробульбарной области может быть причиной образования редких ва риантов отхождения ВА: самостоятельное отхождение ОВ и ПМЖВ от 2-го лицевого синуса аорты или отхождение ОВ от 1-го лицевого синуса аорты или правой ВА, например.
23
кровотока и т. д.), обусловливает их отсутствие в дефинитивном сердце. Идентич ность же первичной закладки редуцированных в норме фрагментов определяет иден тичность их локализации: а) в случае появления необходимости их функционирова ния (коллатерали) или б) сохранения (в редких вариантах нормы).
Центральное же, перитрункальное, кольцо после присоединения к аорте (или ле гочному стволу) претерпевает специфические изменения, приводящие к раздельному, двухкоронарному (чаще всего) кровообращению в сердце. Такие изменения связаны с рядом моментов, на которых мы остановимся ниже, после описания процесса его при соединения к аорте, в соответствии с хронологией осуществления этих процессов.
Далее в литературе обсуждается вопрос о присоединении венечно-артериального сплетения к артериальному полю. В основном дискутируются сроки присоединения: до или после появления обособленных артериальных отверстий, до или после септации конотрункуса, до или после формирования заслонок артериальных клапанов. Ответ на эти вопросы позволяет понять последовательность процессов септации и сепарации от дельных сегментов сердца и интеграции венечно-артериальной сети в систему артери ального кровообращения. Понимание последовательности этих процессов и механизма интеграции сплетения ВА важно и для ответа на такие вопросы, как: 1) почему в дефи нитивном сердце, как правило, имеются две ВА? 2) почему ВА в норме отходят от аор ты, а не от легочного ствола? 3) в каких случаях они отходят от легочного ствола? Тре буют ответа и многие другие вопросы, и мы вернемся к ним позже.
Между тем существует и другая группа вопросов, не менее важных. Какова роль венечно-артериального сплетения (целиком как системы или ее частей) в формирова нии вариантов нормы или аномалий ВА? Существуют ли какие-либо закономерности в распределении ВА? Если они есть, то чем обусловлены? В чем заключаются основ ные различия в строении ВА в нормальных сердцах и в сердцах с транспозицией аор ты и легочной артерии (ТАЛА) или отхождением аорты и легочной артерии от ПЖ (ОАЛА ПЖ), например? Почему при тетраде Фалло часто встречается отхождение пе редней межжелудочковой ветви от правой ВА?
Однозначного ответа на эти вопросы в литературе нет. К сожалению, большей час тью отсутствуют и сами вопросы. Причина этого, на наш взгляд, кроется в том, что эпикардиальному сплетению как системе (сначала эмбриональной, а в дефинитивном серд це зрелой) не уделяется должного внимания. До последнего времени данная система представляла интерес в основном в плане изучения ее присоединения к аорте (и была интересна, как правило, только для эмбриологов). При таком подходе понятно отсутст вие критериев нормальности ВА. Между тем при условии, что критерии нормальности анатомических образований должны основываться на представлении о нормальном эм бриогенезе и морфологических факторах - критериях (И. И. Беришвили и др., 1991; P. Angelini, 1989), основу теоретических обобщений, как подсказывает здравый смысл, следовало бы искать именно в формировании и особенностях этой системы.
Здесь, естественно, есть свои сложности, связанные с набором необходимого числа препаратов для поэтапного изучения важнейших периодов развития сердца и ВА. Но это сложности общего порядка, связанные с технической невозможностью изуче ния в динамике процессов, происходящих в сердце (T.Pexieder et al., 1989). Однако здесь есть и свои преимущества. Существование большой гаммы пороков сердца предо ставляет исследователю уникальную возможность участия в своего рода естественном эксперименте по изучению большого спектра различных вариантов строения ВА (P. Angelini, 1989). Мы пошли по этому пути, изучив разнородный материал: сердца с широким диапазоном отклонений, изменений и нарушений на уровне всех сегментов. При этом общий анализ данных осуществляли в сопоставлении с материалами изуче ния ВА в норме. На основании оценки этих данных мы пришли к выводу об уникаль ности венечно-артериального сплетения: система построена таким образом, что крово снабжение каждого сегмента сердца продублировано два и даже три раза, обеспечивая надлежащее кровоснабжение всех участков сердца независимо от его строения (нор мального или аномального) и образующегося в нем типа соединения ВА.
24
Проанализировав различные варианты отхождения, ветвления и следования ВА в бо лее чем 500 сердцах, мы пришли к выводу, что в принципе все существующие варианты соединения ВА можно разделить на несколько групп. При этом в одну группу могут по пасть и нормальные сердца, и сердца с врожденными пороками, причем разными.
Сопоставив указанные варианты и обнаружив, что группы различаются, в прин ципе, только по набору сегментов, участвующих в образовании венечно-артериальной системы сосудов, мы пришли к выводу о наличии стандартной исходной организации венечно-артериального сплетения, в которой в процессе окончательного формирова ния сердца по ряду причин (детальнее остановимся на них ниже) задействованы те или иные сегменты (тот или иной набор сегментов) общей исходной системы, причем в од ной группе обязательно одни и те же.
Таким образом, закладка системы ВА эмбриологически константна. Констант ность зон закладки «первичных» колец В А и идентичность строения сформировавше гося первичного венечно-артериального сплетения в человеческом сердце обусловле ны постоянством (числа и локализации) сегментов (и, соответственно, - «коронарогенных зон») в первичной сердечной трубке.
В дальнейшем в процессе формирования окончательной (зрелой) системы ВА часть сегментов первичного венечно-артериального сплетения сохраняется, а часть - редуци руется. С небольшими изменениями большая сохранившаяся часть сплетения представ ляет собой остов существующей в постнатальной жизни системы коронарных артерий - эпикардиальных стволов двух систем ВА: правой и левой; меньшая часть исчезает, со ставляя потенциальную основу коллатерального кровообращения и редко встречающих ся вариантов нормы. При различных врожденных пороках сердца (ВПС) инволюция этой меньшей части сплетения неоднозначна. Сохранение тех или иных сегментов - основа су ществующей при различных пороках сердца системы соединения ВА. При приобретен ных патологических состояниях, развивающихся в исходно нормальном сердце, основа компенсации коронарного кровообращения та же и от случая к случаю может отличать ся только по набору участвующих в кровообращении «запасных» сегментов.
Ретроспективный анализ данных литературы о формировании проводящих путей в сердце (R.Anderson et al., 1977; R.Anderson and A. Becker, 1980) показал сходство на шей гипотезы о морфогенезе венечно-артериальной системы с современными пред ставлениями о морфогенезе проводящей системы сердца. В частности, исходя из того, что у низших животных кольца специализированной ткани (проводящей системы) представляют собой соединительную ткань между сегментами формирующегося серд ца и, сопоставив эти данные с некоторыми находками в сердце, было предположено (A. Wenink, 1976), что проводящая система человека тоже образуется из соединитель нотканных колец специализированной ткани. В дальнейшем изучение механизмов развития врожденной атриовентрикулярной блокады подтвердило правомочность предположения о развитии проводящей системы сердца человека из четырех соедини тельнотканных колец, располагающихся между пятью основными сегментами пер вичной сердечной трубки. В процессе формирования сердца эти «первичные» кольца переставляются в иную позицию, причем три из них, соединившись в области схожде ния предсердий, бульбуса и желудочкового компонента, образуют остов дефинитив ной проводящей системы сердца (R. Anderson et al., 1976,1977,1980; A. Wenink, 1976).
Разрабатывая свою концепцию формирования ВА, мы не располагали сведениями о теории морфогенеза проводящей системы сердца человека, предложенной группой исследователей из Нидерландов и Англии (A.Wenink, 1976; R.Anderson et al., 1977).
И хотя наши предположения основаны только на сопоставлении вариантов строения ВА, существующих в сердцах с различным разворотом конотрункуса, сопоставление наших соображений с фрагментами эмбриологических исследований по зарождению ВА и теорией эмбриогенеза проводящей системы сердца, делает, на наш взгляд, пра вомочность существования предлагаемой гипотезы вполне реальной.
Наша концепция формирования венечно-артериальной системы также опирается на предположение, что первоначально венечно-артериальная система зарождается
25
в виде разобщенных колец коронарогенных зон, закладывающихся вокруг межсег ментных втяжений первичной сердечной трубки. Далее вследствие петлеобразования сердца и целого ряда преобразований в процессе его окончательного формирования эти кольца сопоставляются в области наружной кривизны бульбовентрикулярной складки. На этом этапе завершается формирование единого субэпикардиального ве нечного сплетения. Создание же двухкоронарной системы кровообращения в сердце - результат дальнейших изменений, и этот вопрос детально обсуждается ниже.
Приведенные в данном разделе материалы имеют важное практическое значение, поскольку определяют наши представления о строении ВА при вариантах нормы (ко торые из-за их редкости в литературе обычно зачисляли в разряд аномалий), при по роках сердца и приобретенных нарушениях в сердце и ВА. В доступной литературе мы не обнаружили этих данных.
4.Соединение субэпикардиального сплетения
сартериальными стволами
|
До недавнего времени, характеризуя развитие ВА, большинство исследователей |
||||||||||||||
(А. |
Т. |
Юрченко, |
1968; Р. |
В. |
Дубинина, 1972; И. |
И. |
Новиков, 1975; А. |
К. |
Габченко, |
||||||
1978; |
Л. |
Г. |
Шелия, 1985; |
G. |
Martin, 1894; |
S. Levis, |
1904; |
Н. |
Bennett, |
1936; S. |
Rao, |
||||
1958; |
J. |
Wilson, |
1961; S. |
Viragh and С |
Challice, |
1981; |
G. |
Hutchins |
et |
al., |
1988; |
||||
P. |
Angelini, |
1989) |
исходили из того, что ВА появляются сначала в виде эндотелиаль- |
||||||||||||
ных «почек» или колбовидных «бутонов», вырастающих из просвета аорты. Ряд авто ров (И. И. Новиков, 1975; A. Hirakow, 1983) считают, что ВА возникают в виде ворон кообразной эндотелиальной инвагинации начального сегмента аорты или будущего синуса аорты. По данным R. Licata (1955, 1956), у эмбриона человека венечные арте рии появляются на 6-7-й неделе утробной жизни. По мнению автора, их зарождение, по-видимому, следует констатировать с момента прободения перитрункальным коль цом лицевых синусов аорты. По различным данным, ВА у эмбриона человека появля ются на 42-45-й день утробной жизни (J. Wilson, 1961), на 37-й день (XVIII горизон та по Streeter; N. Sissman, 1970), на 38-42-й день (Р. В.Дубинина, 1972), на 39-й день (D. Goor и W. Lillehei, 1975). Z. Rychter и В. Ostadal (1971), изучив механизм развития ВА у куриных эмбрионов, показали, что слепой отросток стенки аорты, являющийся зачатком ВА, присоединяется к ретробульбарному венозному сплетению, проникаю щему в межжелудочковую перегородку (МЖП). Только после присоединения к веноз ному сплетению формируются примитивный ствол венечной артерии и ее главные вет ви. Из этого авторы делают вывод, что ствол и главные ветви ВА формируются путем трансформации венозного сплетения сердца. Согласно исследованиям G. Conte и A. Pellegrini (1984), существующая сосудистая сеть при сближении с артериальным стволом (трункусом) индуцирует образование венечных ростков. В течение XIX гори зонты (после завершения септации трункуса на аорту и легочную артерию и образова ния полулунных клапанов) компоненты венечно-артериального сплетения соединя ются, приводя к образованию нормальной венечной циркуляции крови от аорты к миокардиальному капиллярному сплетению (и далее к венечным венам).
Таким образом, согласно мнению большинства исследователей, венечно-артери- альная система развивается из трех различных компонентов: синусоид (первый ком понент), формирующихся как емкостные образования и осуществляющих обмен меж ду миокардом и полостями сердца; эндотелиальных сосудов, развивающихся в субэпикардиальном слое (второй компонент) самостоятельно, и венечных зачатков (про ксимальных ВА), образующихся из стенок артериальных стволов.
Несмотря на некоторые различия в толковании механизма соединения, по данным отдельных авторов, процесс формирования венечно-артериальной системы предпола-
26
гает обязательное соединение этих раздельно образующихся компонентов. По мнению большинства авторов, сначала субэпикардиальная сеть проксимально соединяется с одним или обоими артериальными стволами. Позже она дистально соединяется с интрамуральной венечно-артериальной системой.
Характеризуя приведенные данные, необходимо отметить, что наибольшие разно гласия касаются именно механизма присоединения второго компонента к третьему. В первую очередь это обусловлено тем, что вопрос генеза системы кровоснабжения сердца человека в литературе ограничен единичными исследованиями (А. К. Габчен-
ко, 1973,1978,1980; Н. А. Джавахишвили и |
М. Э. Комахидзе, 1982; Л. Г. Шелия, |
1985; |
|
J. Wilson, |
1961; R. Hirakow, 1983; G. Conte |
and A. Pellegrini, 1983; A. Bogers et al., |
1988; |
G. Hutchins |
et al., 1988). Экспериментальная же эмбриология до настоящего времени |
||
не смогла разработать приемлемую модель животного, позволяющую изучить процесс соединения второго и третьего компонентов ввиду «скоротечности появления зачат ков ВА» (F.Orts-Llorca et al., 1982; A. Bogers et al., 1988; G. Hutchins et al., 1988; P.Angelini, 1989) и невозможности оценить этот процесс, что связано с недостаточной разрешающей способностью используемой техники (A. Bogers, 1988; P.Angelini, 1989). Лишь недавно с помощью специального метода протравливания материала (A. Bogers, 1989) этот процесс удалось изучить на эмбрионах перепелов.
Подробнее остановимся на этих вопросах ниже. Скажем лишь, что согласно этим дан ным, венечно-артериальное сплетение сначала дистально соединяется с интрамуральной системой, а позже, после разделения артериального отверстия, - и проксимально.
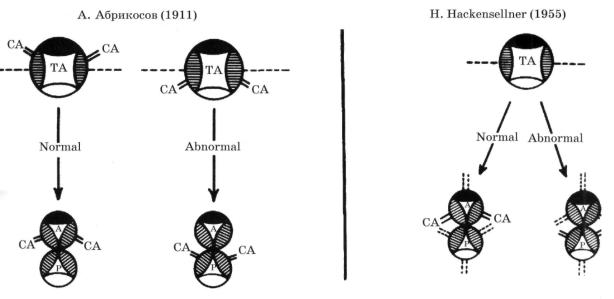
Из существующих гипотез, объясняющих механизм отхождения ВА от артериаль ных клапанов, до последнего времени наибольшее распространение получили теории А. Абрикосова (1911) и Н. Hackensellner (1955) (рис. 4).
Рис. 4. Схема формирования венечных артерий по А. Абрикосову (1911) и Н. Hackensellner (1955).
СА - коронарная артерия; ТА - общий артериальный ствол; А - аорта; Р - легочная артерия. Объяснение в тексте.
В соответствии с гипотезой Абрикосова, основывающейся на работах G. Martin (1894), ВА в человеческом сердце образуются из двух эндотелиальных зачатков артери ального ствола (трункуса), формирующихся еще до его разделения на аорту и легочную артерию. В норме зачатки ВА расположены в аортальной части делящихся эндокардиальных «подушечек», дающих начало лицевым заслонкам аорты и легочной артерии, вследствие чего после разделения трункуса устья венечных артерий располагаются в аортальных синусах, смотрящих на легочный ствол (ЛС). При неправильной заклад-
27
ке зачатков ВА (в легочной части трункуса) или при неправильном разделении общего артериального клапана ВА отходят от заслонок клапана легочного ствола. Теория несо стоятельна по нескольким причинам. Она не объясняет, почему первичные зачатки ВА
впреобладающем большинстве случаев закладываются в аортальной части трункуса и
вкаких случаях (и почему) - в легочной. Кроме того, как показывают наши данные, а также материалы A. Bogers и соавт. (1988), G. Hutchins и соавт. (1988), в преоблада ющем числе наблюдений имеется равноделенность трункуса, в том числе и в сердцах с аномальным отхождением левой ВА от ЛС. Более того, как показали исследования A. Gittenberger-de Groot и соавт. (1983), деление артериального отверстия и образова ние артериальных заслонок - процессы самостоятельные, и последний не имеет отно шения к формированию устьев ВА. Несостоятельность теории Абрикосова окончатель но доказывается фактом образования ВА уже после (а не до!) разделения артериально го ствола на восходящую аорту и легочный ствол. И хотя в литературе на этот счет име ются некоторые расхождения7 , несостоятельность теории Абрикосова очевидна.
Вторую, наиболее расхожую теорию происхождения ВА предложил Н. Hackensellner (1955). Согласно этой теории, аорта и легочная артерия в соответст вии с числом синусов, имеющихся в их клапанах, содержат по шесть первичных за чатков ВА. По мере окончательного формирования ВА персистируют только два ве нечных ствола, отходящих от лицевых синусов аорты, в то время как остальные реду цируются. Эта теория, несомненно, содержит рациональное зерно, заключающееся
вконстатации потенциальной возможности отхождения ВА от любого синуса аорты или легочной артерии. В подтверждение правомочности гипотезы Н. Hackensellner, S. Heifeltz и соавт. (1986) обнаружили первичные зачатки ВА в виде двух фиброзных тяжей, тянущихся от синусов аорты у двух больных с тотальным отхождением В А от ЛС. На наш взгляд, несостоятельность теории заключается в том, что она не может объяснить механизма отхождения двух и более ВА от одного венечного синуса. Более того, при ее правомочности значительно чаще встречалось бы аномальное отхождение ВА от ЛС и значительно большим было бы число случаев отхождения ВА от нелице вых синусов. Кроме того, теория не объясняет, почему пенетрирующими чаще всего оказываются стволы ВА, отходящие от лицевых синусов аорты.
Справедливости ради отметим, что в определенной степени правомочность гипоте зы Н. Hackensellner по этой позиции недавно подтвердили G. Hutchins и соавт. (1988). Опираясь на круговую (по всему периметру артериального отверстия) теорию зарож дения проксимальных ВА, авторы объясняют избирательное соединение перитрункального кольца с лицевыми синусами аорты катеноид ной формой последних. Однако и теория указанных авторов не объясняет причину катеноидной формы одних и некатеноидной формы других синусов. Более того, при малых размерах и нефиксирован ное™ артериальных заслонок вряд ли с достоверностью можно судить о различиях
вих форме, тем более, что тщательные исследования G. Moore и соавт. (1980) не под тверждают каких-либо различий в строении и форме синусов.
Завершая обзор эмбриогенеза ВА, можно заключить, что характер зарождения ве- нечно-артериального сплетения в общих чертах известен. Сегодня уже ясно и то, что не В А растут из аортальных выростов, а уже сформированное сплетение В А пенетрирует стенку аорты (A. Bogers, 1989) (рис. 5). Но сам механизм присоединения сплетения к ар териальным стволам и причины избирательного их присоединения к лицевым синусам
аорты неизвестны |
(В. Maron et al., 1974; М. Meredith et al, 1979; S. Teal et al, 1986; |
M. Lomonico et al, |
1986; A. Bogers, 1989). |
7 Поскольку начало формирования ВА большинство авторов фиксируют с момента появления венечных зачатков аорты, представляет интерес сопоставление этого факта с процессом септации артериального ствола и артериального отверстия. Согласно данным R. Rowe и L. James (1957), к моменту закладки за чатков ВА трункус уже разделен на аорту и ЛС. Как полагают С. Bull и соавт. (1982), это происходит уже после формирования заслонок легочного клапана. Согласно материалам М. Lomonico и соавт. (1986) и G. Hutchins и соавт. (1988), ВА подсоединяются к артериальным клапанам еще позднее - после заверше ния процесса ротации конотрункуса (XVI-XIX горизонты no Streeter).
28