

9. Физиология мозжечка
Мозжечок располагается у человека позади больших полушарий над продолговатым мозгом и мостом мозга. В эволюционном плане мозжечок представляет собой очень древнюю структуру. Впервые мозжечок появляется у круглоротых, что связано преимущественно с усложнением моторной координации.
9.1. Морфофункциональная организация мозжечка
Анатомически в мозжечке можно выделить среднюю часть – червь, полушария мозжечка и боковые флокулонодулярные доли. Наиболее филогенетически молодой частью мозжечка является передняя часть задних долей полушарий мозжечка (неоцеребеллум). В полушариях мозжечка выделяют верхнюю поверхность – кору мозжечка и скопления нервных клеток – ядра мозжечка (рис. 2.5).
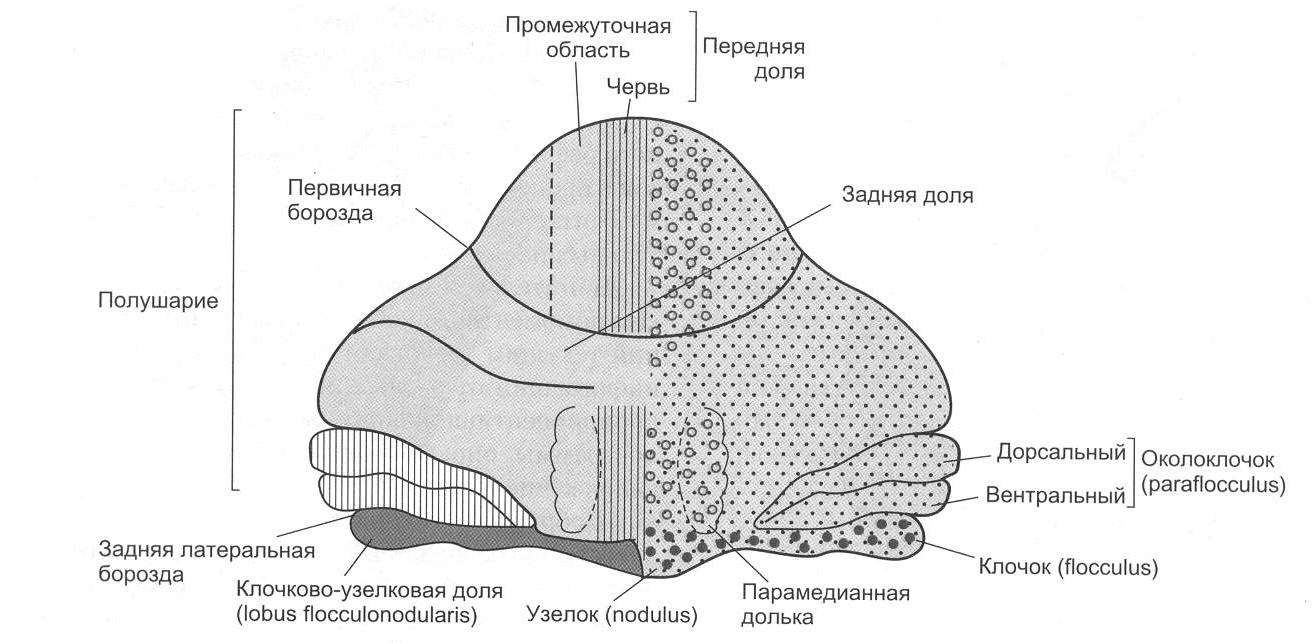
Рис. 2.5. Подразделение мозжечка (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004)
Нейронная организация коры мозжечка отличается исключительной упорядоченностью. В ней выделяют три слоя:
I – поверхностный, или молекулярный слой. В этом слое располагаются дендритные разветвления грушевидных нейронов (клеток Пуркинье), параллельные волокна, которые являются аксонами вставочных нейронов. В нижнем слое молекулярного слоя находятся тела корзинчатых клеток, аксоны которых образуют синапсы с телами грушевидных нейронов, и некоторое число звездчатых клеток.
II – ганглиозный слой. Здесь располагаются тела грушевидных нейронов или клеток Пуркинье. Клетки Пуркинье являются одними из наиболее сложно устроенных нейронов центральной нервной системы. Для этих нейронов примечательным является чрезвычайное ветвление дендритов. Дендритные отростки (дистальные части в особенности) обильно покрыты шипиками, что вместе с разветвлениями дендритов увеличивает площадь поверхностной мембраны, а следовательно, создает условия для размещения огромного числа синапсов. Один грушевидный нейрон может иметь до 200 000 синапсов.
III – гранулярный слой. В этом слое находятся тела вставочных нейронов (клеток-зерен, или гранулярных клеток). Аксоны клеток-зерен поднимаются в молекулярный слой, где они Т-образно разветвляются. Здесь же находятся клетки Гольджи, аксоны которых также направляются в молекулярный слой (рис. 2.6).
Важным для понимания функций мозжечка является изучение синаптических переключений в коре мозжечка и роли ее структур в деятельности ядер мозжечка. Изучением межклеточных связей в коре мозжечка занимался Экклс (рис. 2.7).
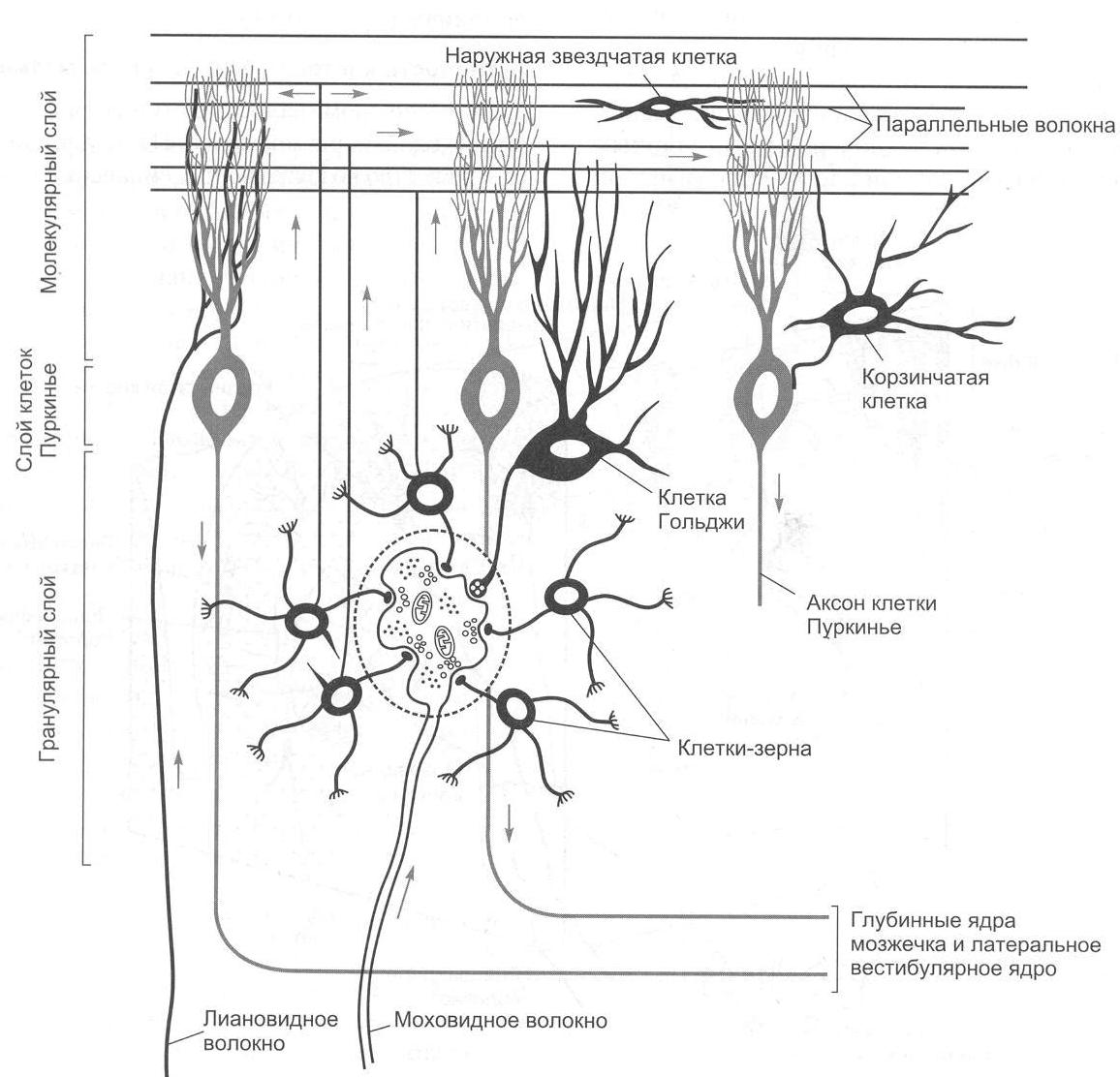
Рис. 2.6. Схема среза коры мозжечка (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004)
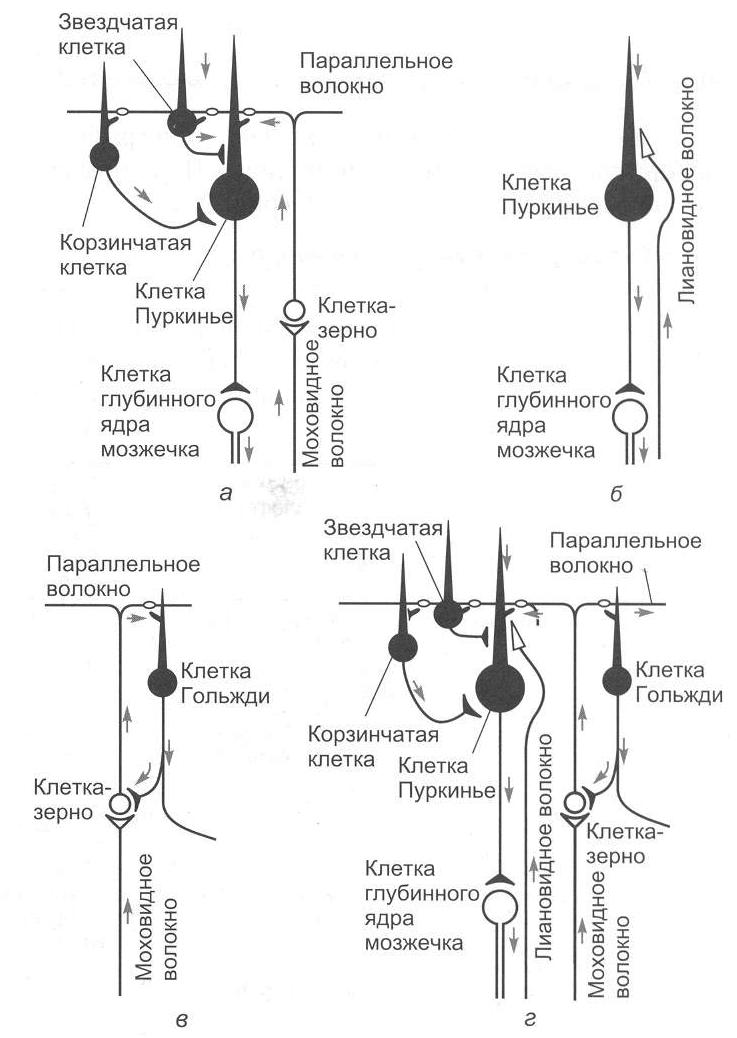
Рис. 2.7. Нейронные цепи коры мозжечка (рисунок взят из книги: Фундаментальная и клиническая физиология / под ред. А. Камкина и А. Каменского. М., 2004)
Вход в мозжечок происходит через кору мозжечка с помощью двух типов волокон – лиановидных (лазающих) и мшистых (моховидных). Лиановидные волокна образуют многочисленные возбуждающие синапсы на дендритах клеток Пуркинье. При этом одного импульса достаточно для того, чтобы клетка Пуркинье ответила целым разрядом.
Мшистые волокна возбуждают клетки-зерна, а те через параллельные волокна оказывают возбуждающее действие на все остальные нейроны, которые, однако, всегда являются тормозными: клетки Гольджи тормозят клетки-зерна по принципу обратной связи, а разряды клеток Пуркинье, воникающие под действием мшистых или лиановидных волокон, приводят к торможению ядер мозжечка. Таким образом, все клетки, нейроны, тела которых лежат в коре мозжечка, кроме клеток-зерен, выполняют тормозные функции. Ни в одном отделе центральной нервной системы нет такого преобладания торможения над возбуждением. Нейроны Пуркинье обладают активностью покоя, который обусловливает тоническое торможение ядер мозжечка. При увеличении активности клеток Пуркинье вследствие возбуждения мшистых или лиановидных волокон торможение ядер мозжечка усиливается; если же происходит торможение клеток Пуркинье (прямое – звездчатыми, непрямое – клетками Гольджи), то ядра мозжечка растормаживаются. Поскольку любое возбуждение, которое поступило в мозжечок, пройдя самое большое, через два синапса, превращается в торможение, уже через 100 мс это возбуждение угасает, и область мозжечка, в которое оно пришло, вновь становится готовой принять новую информацию. Возможно, такое автоматическое стирание информации играет важную роль в связи с участием мозжечка в организации быстрых движений.
В состав мозжечка входят следующие ядра: ядро Шатра, вставочное ядро (у человека это ядро состоит из пробкового и шаровидного ядер) и зубчатое ядро.
Афферентные и эфферентные связи мозжечка. Афферентные и эфферентные связи мозжечка очень многочисленные. Мозжечок получает афферентные связи от вестибулярных ядер, соматосенсорные пути, главным образом от спинного мозга, нисходящие пути – в основном от коры головного мозга.
От спинного мозга к мозжечку подходят два спиномозжечковых пути – вентральный и дорсальный (Говерса и Кларка). В настоящее время выделяют приблизительно еще 10 трактов. Примерно половина из них, включая пути Говерса и Кларка, входят в мозжечок в виде мшистых волокон. Остальные же пути представляют собой спинооливарные тракты, которые, переключаясь на нейроны в оливах, посылают лиановидные волокна к коре мозжечка.
Все области коры больших полушарий, а также другие отделы головного мозга посылают нисходящие пути к мозжечку. Большинство этих путей образуют синапсы в ядрах варолиева моста, волокна от которых идут к нео- или понтоцеребеллуму. Сигналы от двигательных зон коры больших полушарий поступают главным образом в промежуточную часть мозжечка, а импульсы от остальных корковых участков – к его полушариям.
Эфферентные связи. Кора червя мозжечка посылает пути к ядру Шатра, средняя часть коры мозжечка (латеральнее червя) – к вставочному ядру, а кора полушарий мозжечка – к зубчатому ядру. Пути от каждого из этих ядер поступают к различным образованиям ствола мозга и больших полушарий. Таким образом, ядра мозжечка являются выходными структурами мозжечка.