

Янко Слава (Библиотека Fort/Da) || slavaaa@yandex.ru || http://yanko.lib.ru || Icq# 75088656
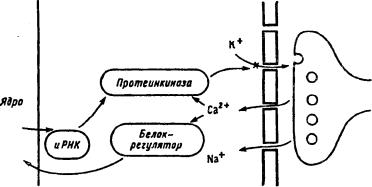
постсинаптической мембраны за счет входящего кальциевого тока включает разные реакции. Эффекты ионов Са2+ реализуются либо путем их непосредственного связывания с белкамимишенями, либо через посредство фосфорилирования мембранных белков специфическими Са2+- завискмыми протеинкиназами. Предполагается, что кальций активирует специфическую протеинкиназу, которая активирует рецепторный белок, т. е. неактивные до этого медиаторные рецепторы постсинаптической мембраны активируются и эффективность синаптической мембраны возрастает. Здесь можно отметить, что Са2+-зависимые протеинкиназы фосфорилируют ряд ферментов и клеточных белков (мембраны и цитоплазмы), которые влияют на состояние проводимости мембраны нейрона (процесс развертывается в пределах 10-20 м после импульсации) (рис. 25).
Существует предположение, что процесс электрогенеза, т. е. генерация импульса нейроном тоже запускает цикл биохимических процессов, опосредованных вторичными посредниками. Сюда включены индукция синтеза циклических нуклеотидов (цАМФ) и возрастание концентрации ионов кальция, активация ими ферментов — протеинкиназ, которые и осуществляют фосфорилирование белков.
Однако, как отмечает И.П. Ашмарин [3], в действии активированных протеинкиназ важно различать два эффекта. С одной стороны, протекает фосфорилирование белков и липидов мембраны клетки. С другой
228
Рис. 25. Схема развития посттетанической потенциации (по Т.Н. Греченко, E.H. Соколову, 1987).
Два контура управления: краткосрочный (медиатор—>Са2+ — протеинки-наза—>инактивация калиевого канала) и долгосрочный (медиатор—>Са2+—>белок-регулятор—>ядро—>иРНК— >синтез протеинкиназы—>усиление блокады калиевых каналов)
стороны, следует выделить процесс фосфорилирования гистонов и других регуляторных белков хроматина, т. е. белков — регуляторов активности генома нейрона. Этот процесс завершается синтезом ряда нейроспецифических белков.
Процессы, обусловливающие изменение состояния мембраны нейрона, развертываются в пределах 10-20 м после импульсации. В этом эффекте участвуют описанные процессы, вызванные действием обычного нейромедиатора, а также возможно участие и нейропептидов-спутников, воздействие которых состоит в относительно кратковременном (опосредованном «вторичными посредниками», а возможно, и прямом) действии на сродство рецепторов постсинаптической мембраны. Число активных рецепторов возрастает и возникает состояние повышенной проводимости синапса, удерживающееся 3-6 суток. Эффективность синапса может быть связана с длительным изменением
229
мембранного потенциала постсинаптического нейрона. Например, нейрогормон АГТГ приводит к фосфорилированию ряда мембранных белков (особенно В-50) и обусловливает длительное уменьшение отрицательного заряда нейрона, тем самым определяется состояние его повышенной возбудимости.
Феномен синтеза нейроспецифических белков индуцируется процессом фосфорилирования регуляторных ядерных белков (белков хроматина). Установлено, что фосфорилирование гистонов, представленных в хроматине вместе с ДНК, осуществляется при участии опять-таки цАМФзависимых протеинкиназ. Это означает, что участие генетического аппарата в регуляции формирования следа памяти обеспечивается механизмами, в основе которых лежит принцип
Данилова H.H. = Физиология высшей нервной деятельности - Ростов н/Д: «Феникс», 2005. — 478 с.
Янко Слава (Библиотека Fort/Da) || slavaaa@yandex.ru || http://yanko.lib.ru || Icq# 75088656
обратной связи: нейромедиатор (через вторичные посредники) принимает участие в активации генетического аппарата, а синтез нейроспецифических белков, часть которых участвует в ферментативной регуляции медиаторных рецепторов, индуцируется генетическим аппаратом. Иными словами, образование и устранение нейромедиатора регулируются ферментами, а синтез и регуляция ферментов осуществляются активацией генов, т.е. транссинаптической индукцией энзимов (ферментов), связанной с избирательной экспрессией генов.
Участие нейроспецифических белков в формировании следов памяти доказывается усилением их новообразования в процессе обучения и регистрации расстройства памяти при блокаде их синтеза. Усиление синтеза белков доказывается увеличением внедрения в них метки после предварительного введения меченых аминокислот. Феномен синтеза нейроспецифических белков проявляется в пределах часа после обучения и достигает максимума на 3-й и 6-й ч[3].
230
Начало исследования нейроспецифических белков мозга было положено в 1965 г. Б.Муром, который выделил и идентифицировал первый из них — белок S-100. К настоящему времени обнаружено свыше трех десятков мозгоспецифических белков, однако вследствие методических сложностей не все они выделены в очищенном виде. Имеются сведения их структурнотаксономического распределения. Так, белок S-100 содержится в мембранах, цитоплазме и ядрах клеток, белок ДНК-110 связан с рибосомами нейронов, ряд белков определяются в синаптических образованиях, а белки 14-3-2 и GP-350 нейрональны по происхождению и локализации. Кроме того, выделена группа белков исключительно глиальных элементов, в частности GFAP, 10В, NS-1, NS-2.
Общепризнанным является представление об исключительной роли мозгоспецифических белков в пластических модификациях, обеспечивающих фиксацию энграммы. Тем не менее убедительные данные об участии в функциях обучения и памяти получены лишь для двух нейроспецифических белков — S-100 и 14-3-2 (или гликопротеина). Об этом свидетельствуют данные о влиянии белка S-100 на фосфорилирование других белков в ядрах нейронов и об усилении синтеза РНК и белков в глиальных клетках. Было показано, что при переучивании крыс в гиппокампе усиливается новообразование именно этого белка. Отмечается, что нарастание синтеза белка S-100 в гиппокампе сопровождается параллельно увеличением концентрации ионов кальция. Интрацеребральное введение крысам антисыворотки к белку S-100 резко нарушает способность крыс к переучиванию. Позднее было обнаружено, что процесс обучения сопровождается новообразованием еще одного специфического белка — гликопротеина, при этом содержание одной из фракций гликопротеина увеличивалось более чем
231
в 4 раза. У золотых рыбок процесс переучивания плавать по-новому (со специальным плавником) сопровождается усилением синтеза двух нейроспецифических гликопротеинов (эпендимов α и β). Интрацеребральная инъекция антиэпендиминовой иммунной сыворотки блокирует консолидацию памяти в этой задаче вестибуломоторного обучения.
Процесс обучения и сохранение выработанных навыков значительно нарушаются под действием ингибиторов синтеза РНК и белков (актиномицина Д, пуромицина, циклогексимида, анизомицина
идр.). Путем торможения синтеза нейроспецифических белков был обнаружен холинорецепторный белок. Блокада синтеза рецепторного белка приводит к нарушению фиксации энграммы. Ряд литературных данных позволяет предполагать, что роль нейроспецифических белков, в частности белка S-100, в механизмах обучения заключается в обеспечении функциональной специфичности разных синаптических входов нейрона. Например, в экспериментах на простой модели обучения — привыкания к сенсорным раздражениям [20; 35] обнаружено, что антитела к белку S-100 значительно ускоряют снижение выраженных ответов в командных нейронах оборонительного рефлекса моллюска на повторные тактильные раздражения. Используя стимуляцию биологически неравнозначных участков тела моллюска, т.е. задействуя разные синаптические входы командного нейрона, было обнаружено, что белки S-100
игамма-глобулины к ним различно влияют на эффективность синаптической передачи в разных входах нейрона. Позднее, в 1984 г. на полимодальных сенсорных входах к командным нейронам моллюска E.H. Соколовым и А.Г. Тер-Маргарян был получен эффект превращения «привыкающего» синапса в «непривыкающий». Блокада белкового синтеза анизомицином (в течение 8 ч и более)
232
привела к «выключению» механизма синаптической пластичности. Торможение белкового
Данилова H.H. = Физиология высшей нервной деятельности - Ростов н/Д: «Феникс», 2005. — 478 с.
Янко Слава (Библиотека Fort/Da) || slavaaa@yandex.ru || http://yanko.lib.ru || Icq# 75088656
синтеза привело к нарушению ферментативных реакций, способствующих пластическим перестройкам в синапсах. Таким образом, существуют основания считать, что продолжительное удержание следов памяти может обусловливаться синтезом нейроспецифических белков, участвующих в транссинаптической индукции ферментов, связанных с активацией (экспрессией) генов нейрона.
К отличительным особенностям генома нервных клеток относится их исключительно высокий по сравнению с другими тканями уровень функционально активных (транскрибируемых) уникальных последовательностей ДНК. Характерным для нервных клеток является прогрессирующее увеличение в них (в течение индивидуальной жизни) числа открытых для синтеза уникальных кодонов ДНК, чего не происходит в тканях других органов. В частности, у эмбриона человека в возрасте 22 недель число гибридных молекул ДНК/РНК, т. е. общее число генов, активных в нервной клетке, составляет 8,2%, а у взрослого человека эта величина достигает 24,6%, а в некоторых зонах мозга — 38%, тогда как в мышцах с возрастом она не меняется.
В онто- и филогенезе прогрессивно увеличивается в нервной ткани число уникальных кодонов ДНК, открытых для синтеза РНК и белков. Существуют доказательства, что транскрибируемость ДНК (а также ее синтез) в нейронах увеличивается как при обучении животных, так и при содержании их в условиях информационно обогащенной среды.
Согласно данным последних лет, объем гибридизации ДНК/РНК в таких отделах мозга, как кора, ретикулярная формация, продолговатый и средний мозг, четверохолмие, значительно различается, коррелируя
233
с мощностью интегративных функций, приписываемых каждой из них. Характерно, что РНК, выделенная из лобной области коры, резко отличается по способности к гибридизации, значительно превосходя по этому показателю все вышеуказанные структуры.
Изучение метилирования ДНК различных нейронов, коррелирующего с их способностью к транскрипции (считыванию), выявило значительно более высокий уровень модификации в неокортексе. Примечательно, что ДНК неокортекса, особенно его гностических зон, у олигофренов существенно менее метилирована по сравнению с ДНК новорожденного ребенка, имеющего нормальную нервную систему.
В настоящее время накоплено немало фактов по интенсификации фракций РНК, часть которых отличается по составу от средних характеристик РНК контрольных нейронов. Однако сейчас принято считать, что «новосинтезируемая» РНК специфична для обучения вообще, а не для каждого вида стимуляции. Это можно продемонстрировать довольно известными экспериментами Э. Кэндела с соавторами по гибридизации РНК/ ДНК. Гибридизация РНК/ДНК проводилась путем использования немеченых препаратов ДНК и РНК из очищенных гонад моллюска и меченого препарата РНК из идентифицированного нейрона, который активировали синаптически. При сравнений РНК нейронов и гонад, т. е. функционально и структурно разных образований одного и того же животного, общей оказывается 92% РНК. Это означает, что только 8% РНК является специфичной для нервной клетки и только 4% ее вовлекается в процесс активации нейрона. При активации нейрона, когда усиливается процесс транскрипции (считывания), образуется новая РНК, но одновременно может быть использована и уже имеющаяся в клетке «нейронспецифическая» РНК.
234
Эти данные сейчас объясняют не синтезом РНК, в которой последовательность нуклеотидов не предшествовала ранее в клетке (например, в ДНК), а тем, что синтез определенной РНК (мРНК) отражает включение определенной комбинации экспрессированных участков генома. Иными словами, обучение объясняется не синтезом РНК, а активацией участков генома, ответственных за метаболизм нуклеиновых кислот и синтез «белков обучения».
Каким образом осуществляется избирательная активация определенной комбинации генов, отражающей ту или иную информацию, остается пока неясным. Существуют предположения, что нейромедиаторы, а также отдельные нейропептиды и нейрогормоны (типа АГТГ) могут проникать непосредственно внутрь ядра нейрона и включать модификацию белков хроматина и РНК, энзиматическое метилирование и избирательный синтез ДНК.
Из литературных источников известно, что существенное усиление процессов метилирования и синтеза ДНК после обучения достигает максимума к 2-3 ч и нормализуется через несколько суток. Исследователи считают, что метилирование ДНК можно рассматривать как один из эпизодов модификации ДНК, причем такая модификация ДНК может быть пожизненной [3].
Данилова H.H. = Физиология высшей нервной деятельности - Ростов н/Д: «Феникс», 2005. — 478 с.
Янко Слава (Библиотека Fort/Da) || slavaaa@yandex.ru || http://yanko.lib.ru || Icq# 75088656
По мнению И.П. Ашмарина [3], даже наиболее устойчивые из известных белков организма имеют период полураспада (сутки, месяцы). Следовательно, в процессе фиксации памяти, включения в мембрану нейрона специфических белков, кодирующих новую информацию, должен одновременно запускаться какой-то механизм их обновления по устойчивому коду. Запись информации выступает как феномен включения (или выключения) участков генома, т.е. как усиление и модификация ДНК при обучении, активации
235
иинактивации генов, ответственных за синтез и обновление белков, устойчиво изменяющих проводимость синапсов. Идея о возможности устойчивого включения — выключения (экспрессиидепрессии) генов, модификации ДНК метилированием-деметилированием была высказана Жакобом и Моно и до сих пор дискутируется.
Относительно механизмов модификации участков генома как феномена, ответственного за синтез
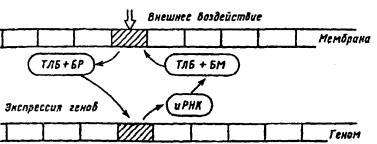
ихранение кода «белков обучения», E.H. Соколов [38; 39] предлагает модель. Согласно его гипотетической модели вся мембрана нейрона функционально проецируется на геном (рис. 26). Каждому локусу мембраны, содержащему специфический рецепторный белок, на геноме соответствует определенный участок экс премированных или депрессированных генов. Воздействие медиатора или его спутника будет избирательно усиливать реактивность соответствующих ему рецепторных белков мембраны. Таким образом, каждый активированный участок мембраны выступает в форме «меченого белка». Белок-«метка» обладает специфической транслокационной группой и
Рис. 26. Гипотетическая схема отражения мембраны на геноме нейрона (по Т.Н.
Греченко, E.H. Соколову, 1987).
ТЛБ — транслокационный белок, БР — белок-регулятор, БМ — белок-модулятор, иРНК — информационная рибонуклеиновая кислота
236
транспортируется к определенным участкам генома, вызывая избирательную экспрессию генов. В результате экспрессии определенных генов и при участии информационных молекул РНК происходит их транскрипция. Синтезируемый энзим (белок-модулятор) транспортируется белком к тому участку мембраны клетки, который был закодирован «маркерным» белком.
Эта схема предполагает, что экспрессия генов, вызванная однократно, затем длительно удерживается, обеспечивая поддержание уровня синтеза белков, регулирующих работу рецепторов, задействованных нейромедиатором активного синапса (см. рис. 26). По этой схеме переход от привыкания к выработке следа памяти происходит по пре- и постсинаптическому механизму (см. рис. 24). В случае выработки нового навыка, условного рефлекса должна начать действовать другая комбинация синапсов и нейромедиаторов. Действует другой белок-модулятор, который переводит соответствующий рецепторный белок в реактивное состояние, и он может длительно присоединять к себе специфический медиатор (см. рис. 25). На пресинаптическом уровне кальций, активируя протеинкиназу, запирает калиевые каналы и тем самым усиливает вход кальция и выброс нейромедиатора в синаптическую щель. На постсинаптическом уровне к активации натриевых каналов присоединяется вход кальция, включение протеинкиназы и выключение калиевых каналов, что повышает возбудимость нейрона. При участии генома нейрона усиливается синтез белков-модуляторов, влияющих на эффективность протеинкиназ, регулирующих закрытие калиевых каналов [10].
Таким образом, в основе кратковременной и долговременной памяти лежат пластические процессы в синаптическом аппарате и в самой соме нейрона.
237
Под пластичностью подразумевается длительное изменение свойств нейрона, влияющих на передачу сигнала. В основе мембранных процессов постсинаптической пластичности лежит
Данилова H.H. = Физиология высшей нервной деятельности - Ростов н/Д: «Феникс», 2005. — 478 с.