

смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность. Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи,
характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 14.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами — млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
14.2. ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата, претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей.
Рассмотрим в отдельности эволюцию скелета и мышечной системы.
14.2.1.Скелет
Ухордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы.
14.2.1.1.Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую
97
функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей — хордом, возникающих на его основе.
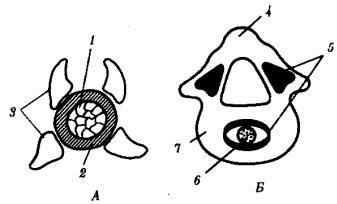
У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов (см. § 13.4). Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка (рис. 14.6).
Рис. 14.6. Развитие позвонка. А—ранний этап; Б—последующая стадия:
1—хорда, 2—оболочка хорды, 3—верхние и нижние позвонковые дуги, 4—остистый отросток, 5—зоны окостенения, 6—рудимент хорды, 7—хрящевое тело позвонка
Замещение хорды позвоночным столбом — более мощным органом опоры, имеющим сегментарное строение,— позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией — заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы.
Урыб только два отдела позвоночника: туловищный и хвостовой. Это связано
сперемещением их в воде за счет изгибов тела.
Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую
98
подвижность головы, а второй — опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5—10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
Урыб все туловищные позвонки несут ребра, не срастающиеся друг с другом
ис грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,— у хвостатых земноводных и пресмыкающихся , поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
Упресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12— 13 пар ребер.
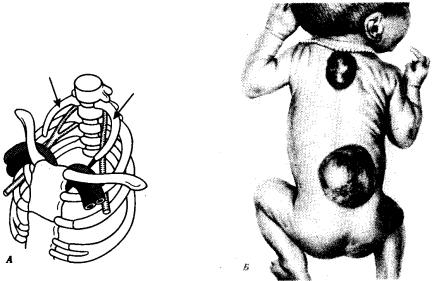
Рис. 14.7. Аномалии развития осевого скелета. А — рудиментарные шейные ребра (показаны стрелками); Б — несращение остистых отростков позвонков в грудной и поясничной областях. Спинномозговые грыжи
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
99
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida — дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа (рис. 14.7).
В возрасте 1,5—3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8—11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
14.2.1.2. Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.
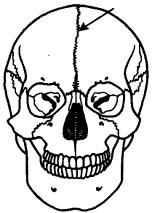
Рис. 14.8. Череп человека с методическим швом (указан стрелкой)
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития:
перепончатую, хрящевую и костную.
100
Укруглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.
Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю.
Укостных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей
сметопическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4—5 пар выполняют опорную функцию для жабр и называются жаберными.
Ухрящевых рыб впереди челюстной дуги располагаются обычно еще 1—2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации.
Челюстная дуга состоит из двух хрящей. Верхний называют нёбноквадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ — первичная нижняя челюсть. На вентральной стороне глотки
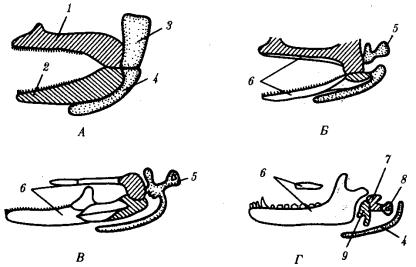
меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным (рис. 14.9).
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей — челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично
101
редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки — столбика,— передающей звуковые колебания от наружного к внутреннему уху.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения.
Одна слуховая косточка — столбик,— характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9).
Рис. 14.9. Эволюция двух первых висцеральных жаберных дуг позвоночных. А—хрящевая рыба; Б—земноводное; В—пресмыкающееся; Г— млекопитающее: 1—нёбно-квадратный хрящ, 2—меккелев хрящ, 3—гиомандибулярный хрящ, 4— гиоид, 5—столбик, 6—накладные кости вторичных челюстей, 7—наковаленка, 8— стремечко, 9—молоточек; гомологичные образования обозначены соответствующей штриховкой
Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной
102