

79. Щитовидная, околощитовидная, вилочковые железы. Строение. Функции.
ЩИТОВИДНАЯ ЖЕЛЕЗА
Щитовидная железа (glandula thyroidea) расположена на шее впереди гортани. В ней различают две доли и перешеек, который лежит на уровне дуги перстневидного хряща, а иногда - 1-3-го хряща трахеи (рис. 120). Щитовидная железа как бы охватывает гортань спереди и с боков. Задне-боковая поверхность каждой доли щитовидной железы прилежит к гортанной части пищевода и передней полуокружности общей сонной артерии. Масса железы у взрослого человека составляет 20-30 г.
Железа покрыта снаружи капсулой, образованной соединительной тканью, с пучками коллагеновых и эластических волокон. От капсулы внутрь железы отходят слабо выраженные перегородки - трабекулы, в которых проходят сосуды и нервы. Трабекулы делят железу на неполные дольки. Паренхима железы состоит из пузырьков-фолликулов, являющихся основными структурными и функциональными единицами (рис. 121).
В щитовидной железе человека имеется около 30 млн фолликулов. Стенки фолликулов образованы одним слоем кубических клеток - тироцитов, лежащих на базальной мембране, в которой имеются отверстия, где клетки соседних фолликулов
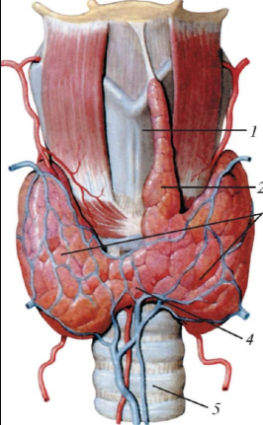
Рис. 120. Щитовидная железа, вид спереди: 1 - щитовидный хрящ; 2 - пирамидальная доля; 3 - правая и левая доли; 4 - перешеек щитовидной железы; 5 – трахея
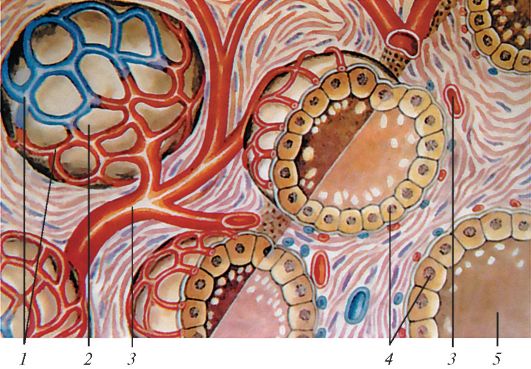
Рис. 121. Схема строения фолликулов щитовидной железы: 1 - кровеносные капилляры; 2 - фолликул; 3 - артерия; 4 - железистые клетки; 5 - коллоид, содержащий гормоны щитовидной железы (по Ю.И. Афанасьеву
и Е.Ф. Котовскому)
контактируют между собой. Встречаются разветвленные фолликулы. Размеры фолликулов колеблются от 0,05 до 0,5 мм. Форма тироцита за- висит от его функционального состояния. Чем клетка крупнее (выше), тем активнее в ней происходят синтетические процессы. В фолликулах имеется рыхлая соединительная ткань. Фолликулы оплетены густой сетью кровеносных капилляров.
В полости фолликула имеется густой вязкий коллоид щитовидной железы, содержащий тиреоглобулин, который синтезируется тироцитами. Крупное ядро расположено в центре тироцита. Цитоплазма тироцитов богата свободными рибосомами, разветвленными широкими цистернами зернистой эндоплазматической сети, развитый комплекс Гольджи расположен над ядром или вокруг ядра. Количество митохондрий невелико, лизосом - множество. Межклеточные щели расширены, кроме участков межклеточных контактов вблизи клеточной поверхности. Поверхность клеток несет на себе множество коротких тонких микроворсинок, погруженных в коллоид. Апикальная часть клеток заполнена многочисленными мелкими гранулами умеренной электронной плотности (апикальные везикулы), которые содержат вещество, похожее по ультраструктуре на коллоид.
Синтез и освобождение гормонов представляют собой сложный процесс. На рибосомах зернистой эндоплазматической сети синтезируется белок, который поступает в цистерны, где к нему присоединяются боковые углеводные цепи. Белок переносится транспортными пузырьками в комплекс Гольджи, где происходит его дальнейшее гликозилирование - образование тиреоглобулина. От комплекса Гольджи отпочковываются просекреторные гранулы, которые, созревая, превращаются в секреторные и затем в апикальные пузырьки, которые выходят в полость фолликула.
Йодирование тиреоглобулина происходит в полости фолликула, в коллоиде. Из крови капилляров в тиреоциты транспортируются ионы йода, откуда он поступает в коллоид. Капельки коллоида, отделенные псевдоподиями, имеющимися на апикальной поверхности тироцита, фагоцитируются. В цитоплазме эти капельки соединяются с лизосомами, ферменты которых расщепляют коллоид, в результате чего освобождаются гормоны трийодтиронин и тетрайодтиронин. Эти гормоны выделяются через базальную поверхность тироцита в перикапиллярное пространство, а из него в кровь. Основная функция этих гормонов - стимуляция окислительных процессов в клетке. Гормоны влияют на водный, белковый, углеводный, жировой, минеральный обмен, рост, развитие и дифференцировку тканей.
Секреция тиреоидных гормонов регулируется ТТГ гипофиза, который, в свою очередь, зависит от ТРГ-рилизинг-гормона. ТТГ действует на тироциты путем активации аденилатциклазы.
В стенках фолликулов между тироцитами и базальной мембраной имеются более крупные, светлые пара(около)фолликулярные клетки (их верхушка не достигает просвета фолликула). Парафолликулярные эн- докриноциты (кальцитониноциты) лежат на базальной мембране. Крупное округлое ядро расположено в центре клетки. В цитоплазме находятся множество пучков микрофиламентов, митохондрий, выраженный комплекс Гольджи. Зернистая эндоплазматическая сеть развита хорошо. В клетках имеется большое количество мембранных аргирофильных секреторных гранул высокой электронной плотности диаметром около 0,15 мкм, содержащих гормон тиреокальцитонин, который выделяется путем экзоцитоза в перикапиллярное пространство, откуда поступа- ет в кровеносные капилляры.
Тиреокальцитонин, участвующий в регуляции обмена кальция и фосфора, является антагонистом гормона паращитовидных желез. Он тормозит реабсорбцию кальция из костей и уменьшает содержание кальция в крови.
Кровоснабжение щитовидной железы. К верхним полюсам обоих долей подходят правая и левая верхние щитовидные артерии (ветви наружных сонных артерий), к нижним полюсам правой и левой долей - нижние щитовидные артерии (из щитошейных стволов подключичных артерий). Венозная кровь от щитовидной железы оттекает по верхней и средней щитовидным венам во внутреннюю яремную вену, по нижней щитовидной вене - в плечеголовную вену (или нижний отдел внутренней яремной вены).
Лимфатические сосуды щитовидной железы впадают в щитовидные, предгортанные, пред- и паратрахеальные лимфатические узлы.
Нервы щитовидной железы происходят из шейных узлов симпатического ствола и от блуждающего нерва.
Развитие и возрастные особенности щитовидной железы. Щитовидная железа начинает развиваться из эпителия передней кишки на 4-й неделе эмбрионального периода. Ее вырост - щитоязычный проток в дистальном отделе делится на два отростка - будущие правую и левую доли. У новорожденного масса железы равна 5-6 г, к 1 году она уменьшается до 2-2,5 г, затем постепенно возрастает, достигая к 12-14 годам 10-14 г, а к 25 годам - 18-24 г. После 60-65 лет масса железы уменьшается в связи с возрастной атрофией.
ПАРАЩИТОВИДНЫЕ ЖЕЛЕЗЫ
Две пары мелких паращитовидных (околощитовидньгх) желез (верхние и нижние) (glandulae parathyroideae)располагаются на задней поверхности долей щитовидной железы (рис. 122), но их число варьирует от 2 до 8. Размеры каждой железы 4-8x3 - 4x2-3 мм. Обычно одна железа расположена вверху, другая внизу, позади каждой доли щитовидной же- лезы. Общая масса всех паращитовидных желез у взрослого человека равна 0,2-0,35 г.
Подобно всем эндокринным железам, паращитовидные железы покрыты тонкой соединительнотканной капсулой, от которой вглубь отходят соединительнотканные прослойки, делящие ткань железы на неполные дольки. Соединительная ткань капсулы и перегородок богата нервными волокнами, кровеносными и лимфатическими сосудами. Паренхима железы образована паратироцитами, формирующими переплетающиеся между собой эпителиальные перекладины. У детей иногда видны фолликулоподобные образования, напоминающие фолликулы щитовидной железы. Различают два вида паратироцитов: светлые главные
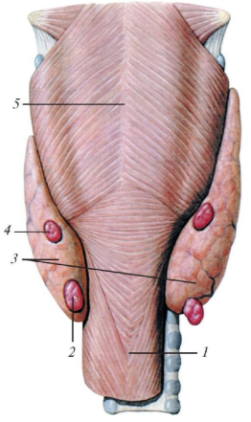
Рис. 122. Паращитовидные железы,
вид сзади: 1 - пищевод; 2 - нижняя левая паращитовидная железа; 3 - щитовидная железа; 4 - верхняя левая паращито- видная железа; 5 - глотка (нижний констриктор)
с базофильной цитоплазмой (у детей в возрасте до 8-10 лет других клеток нет) и ацидофильные (появляются после 8-10 лет). Каких-либо закономерностей расположения клеток обоих типов в паренхиме железы не выявлено. Мелкие полиэдрические главные клетки имеют базофильную цитоплазму, богатую свободными рибосомами, хорошо развитый ком- плекс Гольджи, от которого отшнуровываются окруженные мембранами электронно-плотные гранулы, размерами 15-20 нм, которые часто ассоциируются с липидными каплями различных размеров. Зернистая эндоплазматическая сеть представлена уплощенными цистернами. Среди главных клеток встречаются тем- ные и светлые паратироциты. В цитоплазме последних много частичек гликогена и мало секреторных гранул. По-видимому, обе разновидности главных клеток характеризуют различные фазы секреторной активности: темные активно секретируют, светлые неактивны, либо находятся в фазе подготовки к синтезу. Границы главных паратироцитов четко очерчены, клетки соединены между собой с помощью десмосом и интердигитаций плазмалеммы.
Крупные оксифильные клетки полиэдрической формы также имеют четкие контуры. Их цитоплазма заполнена мелкими митохондриями с ламеллярными кристами и частичками гликогена, комплекс Гольджи выражен слабо. В настоящее время считается, что оба вида клеток - по существу одни и те же клетки на разных этапах развития и функционального состояния. Главные клетки синтетически активны, оксифильные представляют стадии жизненного цикла паратироцитов. Между клетками
обоих типов существуют переходные формы. Возможно, что некоторые оксифильные клетки сохраняют способность синтезировать гормон.
Тяжи и скопления паратироцитов окружены базальной мембраной и сеточкой, образованной нежными ретикулярными фибриллами. Кровеносные капилляры проходят в соединительнотканных прослойках. Секреторные гранулы опорожняются по всей клеточной поверхности, гормон выделяется в перикапиллярное пространство, а затем в просвет капилляров. Главные клетки синтезируют и секретируют белковый паратиреоидный гормон, который регулирует концентрацию кальция и опосредованно фосфора в крови и тем самым влияет на возбудимость нервной и мышечной системы. После удаления паращитовидных желез содержание кальция в крови снижается, а фосфора повышается. Гормон действует на костную ткань, активируя остеокласты, которые обусловливают деминерализацию костной ткани, что приводит к выделению ионов кальция и фосфора в кровь. Избыток фосфора под влиянием гормона удаляется почками. Вместе с тем гормон усиливает реабсорбцию кальция в почке, что способствует уменьшению его выделения с мочой и повышению содержания в крови. Гормон повышает всасывание кальция в кишечнике при условии поступления в организм необходимого количества витамина D. Антагонистом паратгормона является тиреокальцитонин, вырабатываемый описанными выше околофолликулярными клетками щитовидной железы. Он тормозит выделение кальция из костей, ввиду чего снижается его содержание в крови. Клетки паращитовидных желез и парафолликулярные клетки реагируют на изменение содержания кальция в крови изменением синтеза и секреции своих гормонов, что и регулирует содержание кальция в крови.
Кровоснабжение паращитовидных желез. Железы снабжаются кровью из ветвей верхних и нижних щитовидных артерий, а также из пищеводных и трахеальных ветвей. Венозная кровь оттекает по венам, прилежащим к артериям.
Околощитовидные железы иннервируются симпатическими волокнами, отходящими от шейных узлов симпатического ствола, и от ветвей блуждающих нервов.
Развитие и возрастные особенности паращитовидных желез. Па- ращитовидные железы начинают развиваться на 7-й неделе эмбрионального периода из эпителия третьего и четвертого жаберных карманов. Эпителиальные зачатки желез обособляются от стенок жаберных карманов и в процессе роста перемещаются в каудальном направлении. В дальнейшем формирующиеся паращитовидные железы занимают
постоянное положение на задней поверхности правой и левой долей щитовидной железы.
У новорожденных масса паращитовидных желез не превышает 10 мг, к 1 году она достигает 20-30 мг, к 5 годам удваивается, к 10 годам возрастает в 3 раза, а к 20 годам достигает постоянной величины, не изменяясь в течение всей жизни человека.