

ЭУМК_Цитология и гистология
.pdfЦитология и гистология
Особенностью внутриклеточных процессов является их четкое
распределение по областям внутриклеточного пространства
(компартментализация). В частности, процесс клеточного дыхания у эукариот происходит только на мембранах митохондрий, синтез белка – только на рибосомах. Концентрирование ферментов в определенных областях, упорядоченное их расположение во внутриклеточных структурах разделяет разнородные процессы и согласует их в пространстве и времени. Гетерогенность структуры клетки обеспечивает возможность одновременного синхронного синтеза различных веществ из одних и тех же предшественников в миниатюрном общем внутриклеточном пространстве. Компактность, присущая метаболизму клетки, особенно ярко выражена в информативности структуры ДНК.
Внутри клетки непрерывно поддерживается определенная концентрация ионов, отличная от их концентрации во внеклеточной жидкости.
Клетки способны захватывать из среды крупные частицы, молекулы, включая белки (пиноцитоз) или даже вирусы и небольшие клетки (фагоцитоз). Это происходит за счет образования впячиваний плазмалеммы, заполненных внеклеточным субстратом. Края впячиваний затем замыкаются и отделяются внутрь клетки в виде пузырьков (рисунок 2.2).
Рисунок 2.2. – Принцип работы фагоцитоза (а) и пиноцитоза (б)
Все клетки эукариот имеют сходный набор органоидов, по единым
Полесский государственный университет |
Страница 21 |

Цитология и гистология
принципам регулируют свой метаболизм, запасают и расходуют энергию, используют генетический код для синтеза белков. Вместе с тем разные клетки организма имеют значительные отличия по размерам и форме, числу органоидов, набору ферментов. Эта особенность структуры клеток связана с
функциональной специализацией.
Доказано, что у всех клеток одного организма геном не отличается по объѐму потенциальной информации от генома оплодотворѐнной яйцеклетки. Различия в свойствах клеток многоклеточного организма связаны с неодинаковой активностью генов. Это обусловливает различную дифференцировку клеток.
Во всех клетках активны гены общеклеточных, неспецифических функций. Именно потому неспецифических, сходных структурно-функциональных признаков и функций в разных клетках значительно больше, чем признаков и функций специфических. Клетки однородные и сходных функций образуют
ткани.
Воздействиями, регулирующими функции клеток, могут быть метаболиты клетки, ионы, действующие либо на гены, приводя к изменению количества фермента, либо на сам фермент, изменяя его активность. Регулирование может осуществляться по принципу обратной связи, когда результат регулирования (вещество, энергия) влияет на характер последующего регулирования. Регулирование обеспечивает устойчивость внутриклеточных процессов даже при значительных изменениях среды
клетки. Помимо неспецифических эндогенных химических механизмов регулирования функций клеток, в многоклеточных механизмах могут существовать экзогенные механизмы регулирования. В этом случае роль
регуляторов могут выполнять соседние однородные клетки. Это возможно при наличии непосредственных клеточных контактов или при опосредованных гуморальных воздействиях. Роль экзогенных регуляторов могут выполнять также клетки или совокупности клеток специализированные на выполнении функции регулирования (управления).
Основой самовоспроизведения эукариотных клеток является митоз. В некоторых тканях количество клеток устойчиво в течение всей жизни организма. В этих тканях делятся относительно малодифференцированные клетки, резерв которых поддерживается на относительно постоянном уровне, а одна из дочерних клеток дифференцируется. Например, у человека ежедневно погибает ~70 млрд. клеток кишечного эпителия (энтероцитов) и 2 млрд. эритроцитов. Во многих других тканях в клеточный цикл входят вполне дифференцированные клетки. Тогда митоз часто не завершается делением клетки, а ограничивается удвоением хромосом (полиплоидия) или вообще не
Полесский государственный университет |
Страница 22 |

Цитология и гистология
начинается, и клетка выходит из цикла после удвоения хроматид (политения). Некоторые ядра не входят в цикл воспроизведения в течение всей жизни дифференцированной клетки (нейроны, волокна скелетных мышц), и тогда продолжительность жизни клетки соответствует жизни организма. Во всех клетках происходит интенсивное обновление веществ и структур. Огромное количество клеток в каждой ткани, их постоянное внутреннее обновление обеспечивают надѐжность работы органов и систем организма.
2. Химический состав и свойства биомембран
Состав и строение биологических мембран. Биологические мембраны состоят из белков и липидов. Углеводы присутствуют лишь в качестве составных частей сложных белков (гликопротеинов) и сложных липидов (гликолипидов). Нуклеиновые кислоты в небольшом количестве бывают ассоциированы с мембранами, но в состав мембранных структур не включаются. Вода составляет 20% от мембранного материала.
Согласно Сингеру и Никольсону, структурную основу биологической мембраны составляет двойной слой фосфолипидов, инкрустированный белками. При этом различают поверхностные (или периферические) и интегральные белки (рисунок 2.3).
Рисунок 2.3. – Строение биологической мембраны (жидкостно-мозаичная модель)
Липиды находятся при благоприятных физиологических условиях в жидком агрегатном состоянии, это позволяет сравнить мембрану с
Полесский государственный университет |
Страница 23 |

Цитология и гистология
фосфолипидным морем, по которому плавают белковые "айсберги". В разных мембранах соотношение между содержанием белков и фосфолипидов сильно колеблется: количество белков в миелиновой мембране в 2,5 раза меньше, чем липидов, а в митохондриях, напротив, белков в 2,5 раза больше, чем липидов. Кроме фосфолипидов и белков, в биологических мембранах содержатся и другие химические соединения. В мембранах животных клеток много холестерина (в сравнимом количестве с фосфолипидами и белками). Есть в мембранах и другие вещества, например, гликолипиды, гликопротеиды.
Жидкостно-мозаичная модель строения мембраны в настоящее время общепринята.
Липиды мембран представлены четырьмя основными группами: фосфолипидами (основная доля), сфинголипидами, гликолипидами и стероидами. Фосфолипиды – это сложные эфиры фосфатидной кислоты. Сфинголипиды, которые являются производными церамида и монофосфорных эфиров различных спиртов, представлены в основном сфингомиелином. Гликолипиды
– гликозильные производные церамида – представлены как нейтральными цереброзидами, так и их кислыми сульфоэфирами – сульфатидами. Производные церамида и нейраминовой кислоты – ганглиозиды – часто выделяют в отдельную группу липидов – гликосфинголипиды. Стероиды представлены холестерином (в мембранах животных клеток), ситостерином (в растительных клетках) и тетрахименином (обнаружен у тетрахимены).
Белки взаимодействуют с мембранным бислоем, в результате чего они либо ассоциируются с поверхностью мембраны – периферические белки, либо пересекают бислой один или несколько раз, прочно интегрируясь в него, – это
интегральные белки (рисунок 2.4).
Участки белка, которые обращены во внеклеточную среду, могут
подвергаться гликозилированию. В мембранах растений и бактерий полисахара играют самостоятельную роль, образуя наружную оболочку. В клетках животных, в которых наружный слой включает углеводы, имеется внутренний цитоскелет, состоящий из актина и других легко полимеризующихся белков; он имеет регулярную связь с мембранными белками и выполняет формообразующую и опорную функцию.
Полесский государственный университет |
Страница 24 |

Цитология и гистология
Фазовое состояние мембранных липидов. Мембранные липиды могут находиться в нескольких фазовых состояниях, т. е. они обладают мезоморфизмом. Два основных ламеллярных состояния, характерных для мембранных липидов в клеточных системах: кристаллическое и жидкокристаллическое – различаются плотностью упаковки и подвижностью находящихся в бислое белковых молекул.
Фазовые переходы мембранных липидов могут быть вызваны изменением температуры среды. Значение температуры, при котором наблюдается фазовый переход, называется критической температурой фазового перехода, или разделения фаз, если различные участки мембраны вследствие гетерогенности липидного состава по-разному отвечают на изменения температуры. Ионы Са2+, изменение числа ненасыщенных жирнокислотных цепей мембранных фосфолипидов и некоторые другие факторы также могут индуцировать фазовые переходы в бислое.
Специфические свойства биологических мембран. Благодаря указанным особенностям биологические мембраны имеют присущие им характерные черты. Они образуют протяженные бислойные структуры малой толщины (6-10 нм), объединяющие белковые и липидные компоненты с различными свойствами. Целостная структура мембраны создается за счет гидрофобных и электростатических взаимодействий, а не за счет ковалентных связей между составляющими ее молекулами белков и липидов. Гидрофобный липидный бислой представляет естественную преграду для проникновения полярных молекул.
Мембранный бислой обладает относительно малой микровязкостью. Другими словами, мембраны рыхло упакованы, что позволяет отдельным компонентам проявлять высокую подвижность в латеральном направлении.
Наружные мембраны клеток отличаются от внутренних по липидному составу и обладают специфическим набором ферментов и рецепторов.
Функции биологических мембран. Как отмечалось, клеточные мембраны отграничивают содержимое клетки (или клеточной органеллы) от окружающей среды. Благодаря наличию специальных рецепторов они воспринимают сигналы из внешней среды (например, молекулы гормонов,
называемые первичными мессенджерами, или посредниками), в ответ на которые образуются вторичные мессенджеры, высвобождающиеся внутрь клетки. Так осуществляется преобразование сигналов, изменяющих клеточный метаболизм в соответствии с изменяющимися условиями среды.
Мембранные рецепторы выполняют функции узнавания (иммунокомпетентная система), адгезии (обеспечение межклеточных контактов, формирование тканей), регуляции активности ионных каналов (электрическая возбудимость, создание мембранного потенциала).
Транспортная функция является одной из важных функций клеточных мембран. Мембрана создает существенные ограничения для проникновения различных веществ, однако она не является полностью непроницаемой:
Полесский государственный университет |
Страница 25 |

Цитология и гистология
небольшие нейтральные молекулы могут проникать через бислой в области структурных дефектов. Этот процесс осуществляется по градиенту концентрации переносимого вещества - из области, где его содержание высоко, в область с более низким содержанием. Такой процесс называется простой диффузией, он осуществляется неизбирательно и с низкой скоростью.
При облегченной диффузии вещества также переносятся в направлении их концентрационного градиента, но с использованием специальных структур - переносчиков или каналов, увеличивающих скорость и специфичность переноса.
Облегченная диффузия, осуществляемая с помощью каналов, не обладает высокой специфичностью (специфичность определяется лишь размерами канала), но протекает с большей скоростью, а процесс переноса не достигает насыщения в широком диапазоне концентраций переносимого вещества. Функционирование каналов в меньшей степени зависит от фазового состояния мембраны, чем функционирование переносчиков. Все эти примеры относятся к пассивному транспорту через мембрану.
Активный транспорт веществ осуществляется такими же механизмами, но протекает против концентрационного градиента и для своего осуществления должен быть сопряжен с энергодающим процессом. Основным источником энергии для активного транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы (рисунок 2.5).
Рисунок 2.5. – Схема транспорта веществ через биологическую мембрану
Полесский государственный университет |
Страница 26 |

Цитология и гистология
Наконец, в клетках широко представлен вторично-активный транспорт, в процессе которого градиент одного вещества используется для транспорта другого. С помощью вторично-активного транспорта клетки аккумулируют сахара, аминокислоты и выводят некоторые продукты метаболизма, используя градиент Na+, создаваемый в ходе работы Na+/К+- АТФазы.
1. Особенности строения клеток про– и эукариот. Клеточная теория
В 1828 году Христиан Эренберг ввѐл в употребление термин «бактерии» и, тем самым, разделил два надцарства доядерные и ядерные организмы. Надцарство доядерных или прокариот образуют сине-зеленые водоросли, бактерии, микоплазмы, риккетсии, хламидии и спирохеты. Надцарство ядерные или эукариоты образует большинство водорослей, грибы и лишайники, растения и животные. Прокариоты являются наиболее древней формой жизни на нашей планете, главным отличием для них является отсутствие морфологически выраженного ядерного аппарата. Поэтому для обозначения этих организмов можно дать следующее определение: прокариоты – это организмы простой структуры, цитоплазма которых содержит кольцевую молекулу ДНК, не имеющую ядерной оболочку, в которых отсутствуют внутриклеточные системы мембран и митохондрии, а у некоторых и клеточная стенка (рисунок 2.6).
Рисунок 2.6. – Строение прокариотической клетки
Полесский государственный университет |
Страница 27 |

Цитология и гистология
Прокариоты имеют все основные клеточные характеристики:
–в клетке прокариот присутствует зона нуклеоида, заполненная ДНК;
–содержимое отделено плазматической мембраной;
–отсутствие митохондрий компенсируется наличием цепи переносчиков электронов и ферментов окислительного фосфорилирования в составе плазмолеммы;
–в основном веществе (матриксе) цитоплазмы имеются многочисленные рибосомы, обеспечивающие синтез белка;
–большинство прокариот размножаются путем прямого деления надвое;
–многие прокариоты обладают подвижностью и имеют сложный аппарат движения.
Более поздними организма появившимися на нашей планете являются представители надцарства эукариот. Несмотря на значительные различия в строении и физиологии этих организмов для них характерен общий ряд признаков:
–наличие обособленного ядра;
–наличием мембранных и немембранных органоидов в цитоплазме;
–сходные физиологические процессы обмена веществ и энергии, роста и размножения (рисунок 2.7).
Рисунок 2.7. – Строение эукариотической клетки
Полесский государственный университет |
Страница 28 |

Цитология и гистология
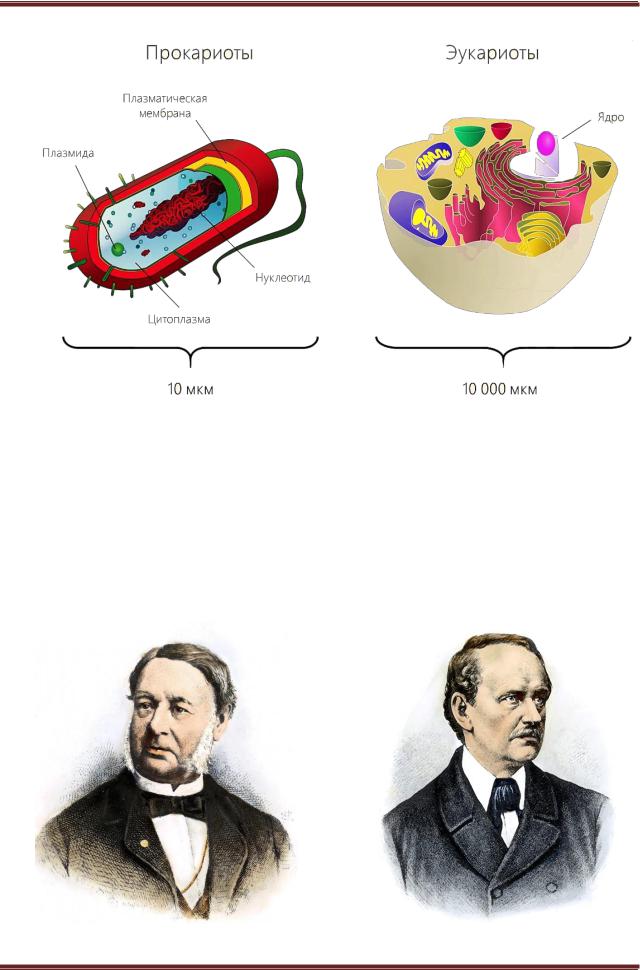
Сравнивая морфологию и физиологии представителей надцарства про- и эукариот (рисунок 2.8, таблица 2.1) можно выделить следующие различия:
1.Клетки прокариот (1–10 мкм), как правило, на порядок меньше клеток экуариот (10–100 мкм).
2.Эукариоты используют только аэробный способ получения энергии, прокариоты активно пользуются и аэробный и анаэробный способом получения энергии.
3.В клетках прокариот присутствуют только немембранные органоиды
–рибосомы, в клетках эукариот имеется многофункциональная система органоидов различного назначения.
4.Синтез РНК и белка в клетках прокариот происходит в цитоплазме, в клетках эукариот он разделен – синтез и процессинг РНК происходит в ядре, а образование белка в цитоплазме.
5.В цитоплазме прокариот находятся рибосомы, имеющие константу осаждения 70 S, в тоже время рибосомы эукариот имеют константу 80 S в цитоплазме и 70 S в митохондриях. Считается, что это дает основание сторонникам теории эндосимбиоза рассматривать полуавтономные органоиды эукариот, как бывшие ранее прокариоты.
6.Прокариоты, обладающие клеточной стенкой, образуют ее из аминосахаров и мурамововой кислоты, тогда как эукариоты, образующие клеточную стенку используют главным образом целлюлозу.
7.Прокариоты, в отличие от эукариот, способны образовывать мукополисахаридную капсулу, благодаря которой обеспечивается резистентность к различным воздействиям (например, фагоцитозу).
8.Наблюдается различие в химическом строении и организации фотосинтетического аппарата.
9.Для эукариот характерен самостоятельный ток цитоплазмы, процессы эндо- и экзоцитоза, а также наличие сократительных белков цитоскелета, обеспечивающих их перемещение.
10.Деление клеток прокариот бинарное, у эукариот – для соматических (вегетативных) митотическое, для половых (генеративных) мейотическое.
Таблица 2.1. – Размеры геномов про- и эукариот
|
Таксон |
Вид |
Длина ДНК, Мб |
Число генов |
|
Прокариоты |
Микоплазмы |
Mycoplasma genitalium |
0,58 |
470 |
|
Риккетсии |
Rickettsia prowazekii |
1,1 |
834 |
||
Археобактерии |
Archaeoglobus fulgidus |
2,18 |
2436 |
||
Цианобактерии |
Synechocystis sp. |
3,57 |
3168 |
||
Эубактерии |
Escherichia coli |
4,6–5,5 |
4288 |
||
Эукариоты |
Грибы |
Saccharomyces cerevisiae |
11,4 |
6241 |
|
Простейшие |
Dictyostelium discoideum |
32 |
11000 |
||
Высшие растения |
Arabidopsis thaliana |
115,7 |
27540 |
||
|
|
|
|
||
Беспозвоночные |
Drosophila melanogaster |
120 |
13600 |
||
Позвоночные |
Homo sapiens |
3000 |
28000 |
||
|
Полесский государственный университет |
Страница 29 |
Цитология и гистология
Рисунок 2.8. – Различия прокариотической и эукариотической клетки
Клеточная теория.
Длительное и пристальное изучение клетки привело к формулированию важного теоретического обобщения, которое называется клеточная теория.
Клеточная теория – это обобщенные представления о строении клеток как единиц живого, об их размножении и роли в формировании многоклеточных организмов.
Теодор Шванн 1810-1882 гг. |
Маттиас Якоб Шлейден 1804-1881 гг. |
Полесский государственный университет |
Страница 30 |