112.Классификация липидов. Роль в жизнедеят-ти клетки. Метаболизм липопротеинов, транспорт липидов между органами и тканями. Нарушение обмена липидов при сердечно-сосудистых.
Л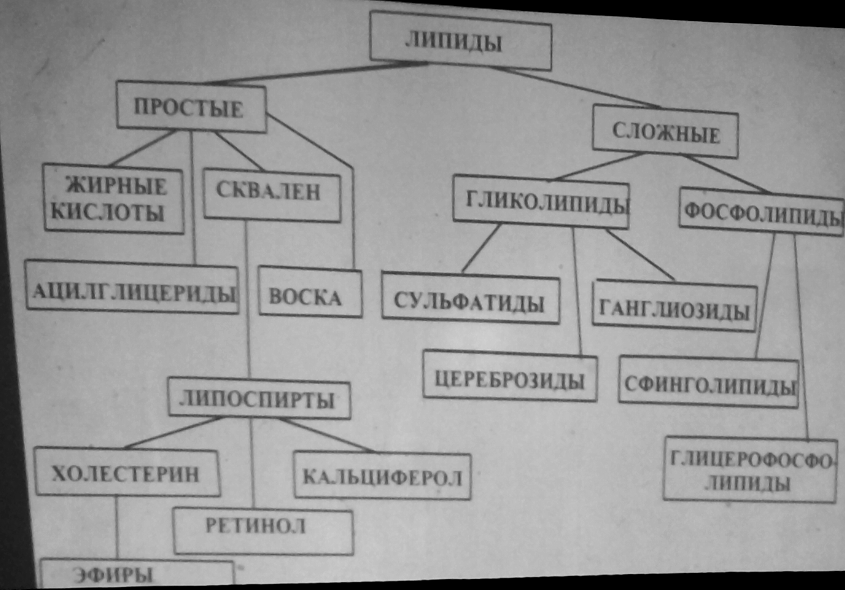 ипиды
- в-ва, обл. общ. физич. св-вом - гидрофоб.
Основ.ф-ция- энергетическая, они дают
в2раза больше энергии чем углеводы;
запасающая ф-ция, ф-ции теплоизоляционной
и мех.защиты, стоительная ф-ция за счет
фосфолипидов, холестерина, участие в
обменных процессах за счет жирораст.
витаминиов, стероидных гормонов.
ипиды
- в-ва, обл. общ. физич. св-вом - гидрофоб.
Основ.ф-ция- энергетическая, они дают
в2раза больше энергии чем углеводы;
запасающая ф-ция, ф-ции теплоизоляционной
и мех.защиты, стоительная ф-ция за счет
фосфолипидов, холестерина, участие в
обменных процессах за счет жирораст.
витаминиов, стероидных гормонов.
ЛП-комплекс белков и липидами, внутри ядро, содерж.неполярные липиды(ТАГ), оболочку из фосфолипидов, белка и своб.холестерина. Ф-ция ЛП-транспорт липидов. ХМ- ЛП,синтезируемые клетками кишечника. В результате действия ЛП-липазы на жиры ХМов, образуются жирные кислоты и глицерол.
Гиперхолестеринемия создает повышенную опасность заболевания атеросклерозом. Вероятность заболевания тем выше, чем больше отношение концентрации ЛНП к концентрации ЛВП в крови; их называют соответственно атерогенными и антиатерогенными липопротеинами. Гл. бх прояв. атероскл. — отложения холест. в стенках артерий.
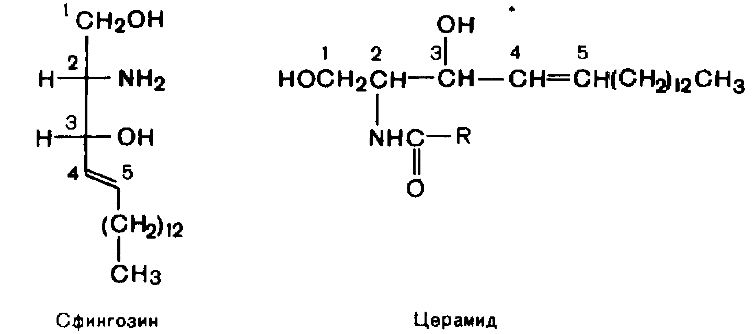
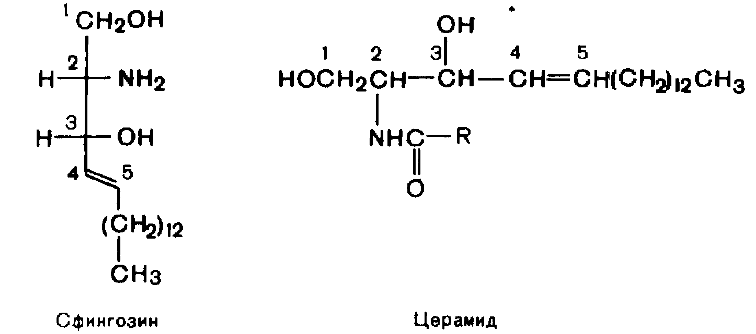
№113 Cфинголипиды. Строение. Роль. Сфинголипиды - производные церамида, образующегося в результате соединения аминоспирта сфингозина и жирной кислоты.. Наиболле распр. сф-ды - сфингомиелины- основные компоненты миелина и мембран клеток мозга и нервной ткани.
Сфинголипидозы
— наслед. болезни обм. в-в, хар-ся пат.
накопл. опр. класса мембранных липидов,
в сост. к-х входит сфингозин. Генет.
дефект церамидазы приводит к развитию
б-ни Фарбера, набл. гепато- и спленомегалия,
а также пораж. суставов. Сфинголипидозы
обычно приводят к смерти в раннем
возрасте, т. к. происх. пораж. кл. нерв.
тк., где сконцентрированы гликосфинголипиды.
№114 Пищевые жиры, их переваривание. Всасывание. Нарушения переваривания и всас. Биосинтез триглицеридов. Переваривание экзо.жира обязательно требует предварительного эмульгирования. Некоторые пищевые жиры поступают в организм уже в эмульгированной форме, например молочный жир. Эмульгаторы – вещества амфифильной природы, сниж. поверхн. натяж. и стабилиз. эмульсию. Создается большая площадь контакта жира с водной фазой, в которой находится фермент. В орг. человека эмульгаторами являются желч. к-ты. В составе желчи желчные кислоты поступают в 12-перстную кишку и аллостерически активируют панкреатическую липазу. Собственно переваривание жиров – это гидролиз сложноэфирных связей. Под действием эстеразы липидов часть моноацилглицеринов может подвергаться гидролизу с образованием глицерина и жирных кислот. Т. о., продуктами перевар. жира являются глицерин, жир. к-ты и моноацилглицерины. Всас. продукты перева. путем предварит. обр. смешанных мицелл с желч. к-тами.
Биосинтез триглицеридов. происх. в энероцитах.. в гладкой ЭПС активир. жир. к-ты, происходит ацилирование моноглицеридов с обр-м сначала диглицеридов. а затем триглицеридов. Катализ триглицеридсинтазой.. Ресинтезированные в клетках кишечной стенки триглицериды соед. внутри цистерн гладкого эндоплазматического ретикулума с небольшим количеством белка и образуют хиломикроны.
Нарушения перевар. и всас. жиров.связ. с недост. поступ. панкреат. липазы в киш., или наруш. поступ. в кишеч. желчи. Наруш. мб. связ. с заб. пищ. тракта (при энтеритах, гиповитаминозах и др забол.). Обр-ся в полости киш. моноглицериды и жир. к-ты не могут нормально всас. вследствие повр. эпит. покрова киш. Кал сод. много нерасщеп. жира или невсосавшихся высших жир. к-т и имеет серовато-белый цвет.
№115, 116Ненасыщенные жир. к-ты. Физ-хим св-ва. Биологическая роль. Примерно 3/4 всех жирных кислот являются непредельными (ненасыщенными), т.е. сод. двойные связи, имеют цис-конфигурацию,которая. делает ненасыщенную кислоту менее стабильной и более подверженной катаболизму. Ненасыщ. жир. к-ты: пальмитиновая (C15H29COOH, 1 двойная связь), олеиновая (C17H33COOH, 1 двойная связь)Полиненасыщ. жир. к-ты: линолевая (C17H31COOH, 2 двойные связи), линоленовая (C17H29COOH, 3 двойные связи), арахидоновая (C19H31COOH, 4 двойные связи).Ткани жив. обладают весьма огранич. спос-тью превращать насыщенные жирные кислоты в ненасыщенные. Поэтому в пище животных должны обязательно присутствовать некоторые полиненасыщенные жирные кислоты, содержащиеся в продуктах растительного происхождения. Фосфолипиды клет. мембр. содержат ненасыщ. к-ты. Высокое содержание полиненасыщ. жир. к-т в пищ. рационе является основным фактором, обесп. сниж. холестерола в плазме крови. Простагландины и тромбоксаны являются гормонами местного действия; при необх. они быстро синт. и действуют в непосредственной близости от места их синтеза. Основная физиол. ф-ция простагландинов состоит в модулировании акт-ти аденилатциклазы и выраж., например, в регуляции агрегации тромбоцитов или ингибировании действия АДГ в почках.
117. Липидный состав мембран. В мембранах имеются фосфолипиды 2 типов — глицерофосфолипиды и сфингофосфолипиды. 1ые- явл. производными фосфатидной к-ты (диацилглицеринфосфата).2ые сод. аминоспирт сфингозин и явл. производными церамидов.
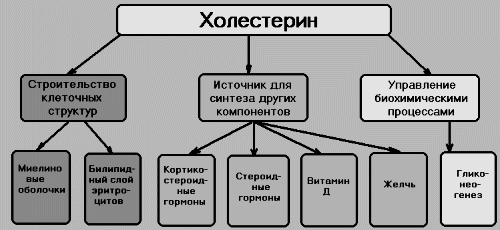
Холестерин
— относится к стероидами.
 Физические свойства мембран зависят
от расположения белков в липидном слое.
Липиды и белки мембран способны к
диффузии в пределах слоя параллельно
поверхности мембраны (латеральная
диффузия). Фосфолипазы-
ферменты класса гидролаз, катал. гидролиз
фосфоглицеридов. В зав-ти от положения
гидролизуемой связи в фосфолипиде
различают 4 осн. класса : A, B, C и D.Фосфолипиды
в кишеч. подвергаются дейст. фосфолипазыА2,
катализир. гидролиз. сложноэфирной
связи в -положении. Глицрофосфолипиды
расщеп. с обр-м лизофосфолипида и жир.
к-ты. Лизофосфо-липид подв.
дейст.лизофосфолипазы панкр. сока,
обр-ся глицерофосфохолин. который всас.
в кровь
Физические свойства мембран зависят
от расположения белков в липидном слое.
Липиды и белки мембран способны к
диффузии в пределах слоя параллельно
поверхности мембраны (латеральная
диффузия). Фосфолипазы-
ферменты класса гидролаз, катал. гидролиз
фосфоглицеридов. В зав-ти от положения
гидролизуемой связи в фосфолипиде
различают 4 осн. класса : A, B, C и D.Фосфолипиды
в кишеч. подвергаются дейст. фосфолипазыА2,
катализир. гидролиз. сложноэфирной
связи в -положении. Глицрофосфолипиды
расщеп. с обр-м лизофосфолипида и жир.
к-ты. Лизофосфо-липид подв.
дейст.лизофосфолипазы панкр. сока,
обр-ся глицерофосфохолин. который всас.
в кровь
118.Распад
и синтез триацилглицеринов.
Синтез жиров происходит в абсорбтивный
период в печени и жировой ткани.
субстратами в синтезе жиров являются
ацил-КоА и глицерол-3-фосфат.
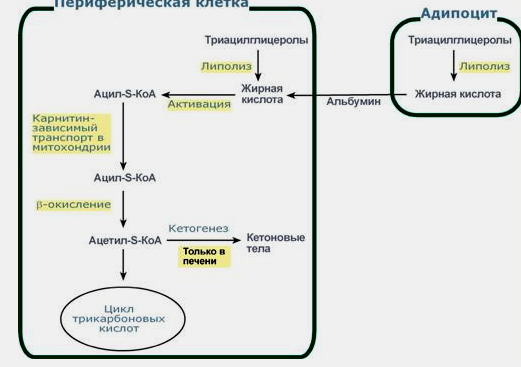 Синтезированные в печени жиры упаковываются
в ЛПОНП и секретируются в кровь.Гидролиз
внутриклет. жира осущ. под действием
гормончувствительного фермента
ТАГ-липазы. Глицерол как водорастворимое
в-во транспортируется кровью в своб.
виде, а жир. к-ты (гидрофобные молекулы)
в комплексе с белком плазмы - альбумином.
Регуляция:
в абсорбтивном состоянии под действием
инсулина происходит липогенез, в
постабсорбтивном состояни-липолиз-расщепление
жиров, активируемый глюкагоном. Адреналин,
секреция которого увеличивается при
физической активности, также стимулирует
липолиз. В абсорбтивный период при
увеличении соотношения инсулин/глюкагон
в печени активируется синтез жиров.
Синтезированные в печени жиры упаковываются
в ЛПОНП и секретируются в кровь.Гидролиз
внутриклет. жира осущ. под действием
гормончувствительного фермента
ТАГ-липазы. Глицерол как водорастворимое
в-во транспортируется кровью в своб.
виде, а жир. к-ты (гидрофобные молекулы)
в комплексе с белком плазмы - альбумином.
Регуляция:
в абсорбтивном состоянии под действием
инсулина происходит липогенез, в
постабсорбтивном состояни-липолиз-расщепление
жиров, активируемый глюкагоном. Адреналин,
секреция которого увеличивается при
физической активности, также стимулирует
липолиз. В абсорбтивный период при
увеличении соотношения инсулин/глюкагон
в печени активируется синтез жиров.
119
Биосинтез и катаболизм фосфолипидов и
гликолипидов. Ф-ции фосфолипидов и
гликолипидов.
 Фосфолипиды играют важную роль в струк.
и ф-ции клеточ. мембр.. актив. мембр. и
лизосомальных ферм., проведении нерв.
имп., сверт. крови, иммунологич. р-циях,
процессах кл. пролиферации и регенерации
тк., переносе эл. в цепи дыхат. ферм.
формир. липопрот. комплексы.
Фосфолипиды играют важную роль в струк.
и ф-ции клеточ. мембр.. актив. мембр. и
лизосомальных ферм., проведении нерв.
имп., сверт. крови, иммунологич. р-циях,
процессах кл. пролиферации и регенерации
тк., переносе эл. в цепи дыхат. ферм.
формир. липопрот. комплексы.
Биосинтез происх. в печени, стенки кишеч., семенниках, яичниах, мол. ж-зе, ЭПС. Центр. роль в биосинтезе играют 1,2-диглицериды и сфингозин. ЦТФ. Фосфатидилэтаноламин-кефалин синт. из этаноламина. и явл. предш. Фосфатидилхолина-лецитин. Сфингомиелин синт-ся в рез-те р-ции церамида с ЦДФ-холином. Различные типы фосфолипаз, лок. в кл. мембранах или в лизосомах, катализируют гидролиз глицерофосфолипидов.
Гликосфинголипиды - гликолипиды, в состав к-х входят церамид и один или несколько остатков углеводов, и к-та. Локализованы в мембр.клеток так, что углеводная часть молек. расп. на пов-ти кл. и часто обладает АГ св-вами. Синтез нач. с обр. церамида. Серин конденсируется с пальмитоил-КоА. Продукт их взаимодействия сначала восст.НАДФН, затем к аминогруппе дигидросфингозина амидной связью присоед. жир. к-та. После окисления ФАД-зависимой дегидрогеназой обр. церамид, к-й служит предш. в синтезе гликосфинголипидов. Послед. р-ции синтеза катализ. специф. трансферазами. Соед. фосфорилхолина с церамидом сфингомиелинсинтазой приводит к образованию сфингомиелина. Присоединение углеводных компонентов катализируется специф. гликозилтрансферазами. Донорами углеводных компонентов служат активированные сахара: УДФ-галактоза и УДФ-глюкоза. Галактоцереброзид - главный липид миелиновых об-к; глюкоцереброзид входит в состав мембран многих клеток и служит предш. в синтезе более сложных гликолипидов.
Катаб. сф-нов и гликолипидов происх. в лизосомах. В распаде сфингомиелинов уч. сфингомиелиназа, отщепляющая фосфорилхолин, и церамидаза, продуктами действия которой являются сфингозин и жир. к-та. В рез-те послед. р-ций гидролиза α- и β-галактозидазой, β-глюкозидазой, нейраминидазой и церамидазой, гликосфинголипиды расп. до моном.: гл-зы, галактозы, жир. к-ты, сфингозина и др. метаб.
120 Обр-е желчных к-ты и их роль в переваривании жиров. Коньюгирование желч. к-т..(см.130)
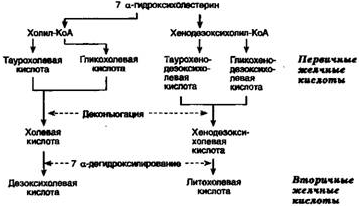 Жёлч. к-ты-эмульгаторы.
В составе желчи желчные кислоты поступают
в 12-перстную кишку и аллостерически
активируют панкреатическую липазу.
Жёлч. к-ты-эмульгаторы.
В составе желчи желчные кислоты поступают
в 12-перстную кишку и аллостерически
активируют панкреатическую липазу.
Жёлч. к-ты - произв.е холестерола. В организме синтез. холевая и хенодезокеихолевая к-ты. Не являются эффективными эмульгаторами. В печени эмульгирующие св-ва жёлч. к-т увеличиваются за счёт р-ции конъюг., в к-й к карбоксильной гр. жёлч. к-т присоед. таурин или глицин. Эти производные - конъюгированные жёлчные кислоты назыв.солями жёлчных кислот. Именно они служат главными эмульгаторами жиров в кишечнике. Вторичные желчные кислоты , включая дезоксихолевую кислоту и литохолевую кислоту, образуются из первичных желчных кислот в толстой кишке под действием бактерий. Литохолевая кислота всасывается значительно хуже, чем дезоксихолевая.
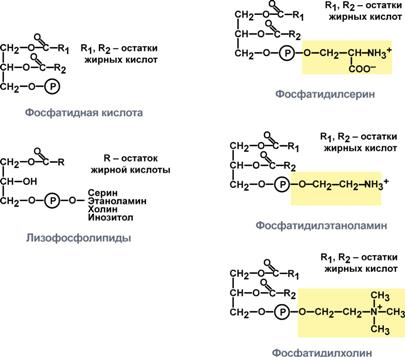
121. Классификация фосфолипидов и пути их биосинтеза. Значение фосфолипидов в жизнедеятельности клетки, участие их в передаче гормонального сигнала. (см.119в)
Фосфолипиды играют важную роль в струк. и ф-ции клеточ.мембр, актив. мембр. и лизосомальных ферм., проведении нерв. имп., сверт. крови, иммунологич. р-циях, процессах кл. пролиферации и регенерации тк., переносе эл. в цепи дыхат. ферм. формир. липопрот. комплексы. Они есть во всех живых клетках.
глицерофосфолипиды— содержат остаток глицерина
фосфатидилхолин (лецитин)
фосфатидилэтаноламин (кефалин)
фосфатидилсерин
фосфосфинголипиды — содержат остаток сфингозина
сфингомиелины
фосфоинозитиды — содержат остаток инозитола
Центральную роль в биосинтезе фосфолипидов играют 1,2-диглицериды (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфин-гомиелинов). Цитидинтрифосфат (ЦТФ) участвует в синтезе практически всех фосфолипидов. В качестве примера рассмотрим синтез отдельных представителей фосфолипидов.
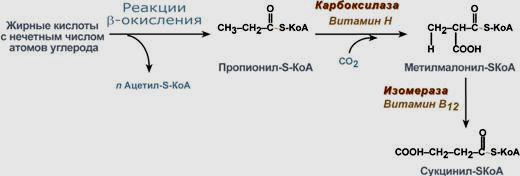
126. Особенности окисления жирных кислот с нечетным числом углеродных атомов. Жирные к-ты с нечетным числом углеводов поступают в организм с раст.пищей и морепродуктами. Их окисление происх.по обычному пути до посл.реакции, в котор.образ.пропионал-SCoA. Суть превращения пропионила сводится к его карбоксилированию, изомеризации и образов. сукцинил-SKoA, в р-циях участвуют биотин и витам.В12.
122.
Химическое строение гликолипидов и их
биологическая роль.
Гликолипиды широко представлены в
нервной ткани и мозге. Размещаются они
на наружной поверхности плазматических
мембран, при этом олигосахаридные цепи
направлены наружу. Наиболее вероятная
их функция – рецепторная.
Общей частью всех гликолипидов является
церамид –
соединение аминоспирта сфингозина с
длинноцепочечной жирной кислотой:

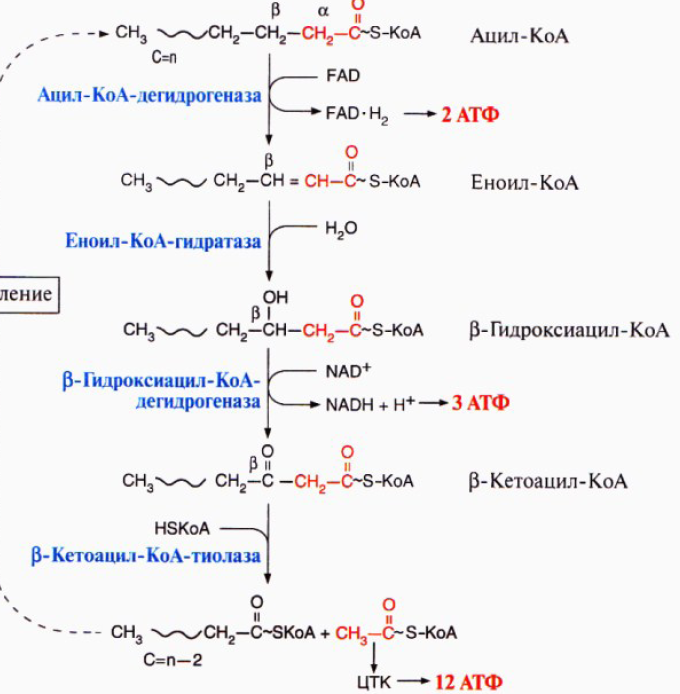
123. β-окисление ненасыщенных жирных кислот – спец.путь катаболизма жир.к-т, продуктом явл.ацетил-КоА. Окисление идет по b-углеродному атому. Служит источ.энергии для синтеза 12АТФ. Процесс идет в матриксе митохондрий в аэробных усл. Для переноса ацил-КоА ч/з мембрану митох.сущ.система – в комплексе с молекулой карнитина, ф-т карнитинацилтрансфераза I(ингибитором явл.малонил-КоА).
4реакции
b-окисления
наз.циклом т.к. эти 4 реакции повтор.до
тех пор пока к-та не расщипится на
ацетильные остатки. Регуляция: при
голодании-глюкагон, в постабсорбтивном
периоде-инсулин.