

43. Декарбоксилирование аминокислот (глу, три) в тканях. Обезвреживание биогенных аминов в печени с участием ферментов мао и дао.
Отщепление α-карбоксильной группы аминокислот в виде СО2 катализиpуется декаpбоксилазами, кофактоpом котоpых является пиpидоксальфосфат. В результате декарбоксилирования образуются биогенные амины:
Серотонин Серотонин активно синтезируется в тучных клетках кожи, легких, печени, в селезенке, ЦНС:
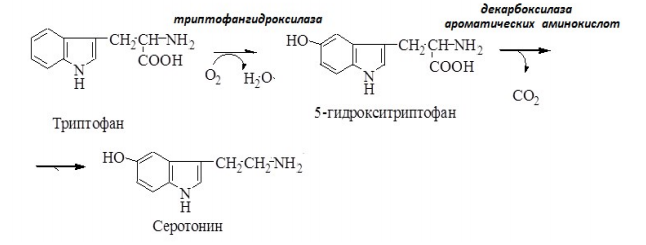
Физиологические эффекты 1. Стимулирует сокращение гладких мышц желудочно-кишечного тракта и, как следствие, повышение перистальтики ЖКТ; 2. Выражено стимулирует сокращение гладких мышц сосудов, кроме сосудов миокарда и скелетных мышц и, как следствие, повышение артериального давления; 3. Слабо увеличивает тонус гладких мышц бронхов; 4. В центральной нервной системе является тормозным медиатором; 5. В периферических нервных окончаниях обуславливает возникновение боли и зуда (например, при укусе насекомых).
γ-аминомасляная кислота (ГАМК) Синтез γ-аминомасляной кислоты происходит исключительно в центральной нервной системе – в подкорковых образованиях головного мозга:

Физиологические эффекты ГАМК. В центральной нервной системе ГАМК (наряду с глутаминовой кислотой) является тормозным медиатором.
Обезвреживание биогенных аминов. Существуют два типа реакций инактивации биогенных аминов – дезаминирование и метилирование. Дезаминирование протекает под действием ферментов моно- и диаминооксидаз (МАО и ДАО) с образованием свободного аммиака и альдегидов, которые окисляются до соответствующих карбоновых кислот и выводятся из организма. МАО и ДАО обнаружены во многих тканях, но наиболее активны в печени, желудке, почках, кишечнике, нервной ткани.

Метилирование биогенного амина происходит при наличии у него гидроксильной группы (дофамин, серотонин). В реакции принимает участие активная форма метионина - S-аденозилметионин (SAM), который превращается в S-аденозилгомоцистеин (SАГ):

44. Пути образования и обезвреживания аммиака в тканях. Токсичность аммиака. Гипераммониемии: причины и следствия.
Пути образования аммиака Аммиак непрерывно образуется во всех органах и тканях организма. Наиболее активными его продуцентами в кровь являются органы с высоким обменом аминокислот и биогенных аминов – нервная ткань, печень, кишечник, мышцы. жизнедеятельность бактерий толстого кишечника, Аммиак является одним из конечных продуктов азотистого обмена. В организме человека аммиак образуется в ходе следующих процессов:
дезаминирование аминокислот в тканях - основной путь;
окислительное дезаминирование биогенных аминов;
распад пуриновых и пиримидиновых оснований;
гниение белков в толстом кишечнике в результате жизнедеятельность бактерий
Токсичность аммиака. Из толстой кишки аммиак поступает в кровь воротной венозной системы. В нормальных условиях печень быстро извлекает аммиак из крови воротной вены, так что кровь, выходящая из печени, практически не содержит аммиака. Но проблема состоит в том, что аммиак – это крайне токсичное вещество, особо опасное для клеток центральной нервной системы. В норме содержание свободного аммиака в крови представлено лишь следовыми количествами (0,4-0,7 мг/л или 25-40 мкмоль/л). Токсическое действие аммиака обусловлено тем, что:
Аммиак диффундирует через мембрану и в клетке гидратируется с образованием гидроксида аммония (NH3 + H2O = NH4OH), что вызывает внутриклеточный метаболический алкалоз. pH вне- и внутриклеточной среды повышается. Это приводит к изменению степени ионизации ионогенных групп и, как следствие, к разрушению ионных связей, стабилизирующих третичную структуру белков. Последнее вызывает изменение нативной конформации белковых молекул и нарушение их функций, в частности, нарушение активности ферментов, структуры мембранных рецепторов.
При повышении внутриклеточной концентрации аммиака активируются процессы восстановительного амминирования α-кетокислот, в частности – α-кетокислот цикла трикарбоновых кислот – αкетоглутарата и оксалоацетата.

Это приводит к истощению пула α-кетоглутарата и оксалоацетата, что сопровождается снижением обменной мощности цикла Кребса, а, следовательно, и нарушением образования АТФ. В результате ингибируются энергозависимые процессы, прежде всего активный мембранный транспорт, что приводит к нарушению процессов реполяризации и поддержания потенциала покоя, а, следовательно, и нарушению таких свойств нервной ткани как возбудимость и проводимость.
При повышении концентрации аммиака, происходит образование глутамина:

В нейронах снижается количество глутамата, и следовательно, тормозится процесс декарбоксилирования глутамата, что приводит к снижению уровня ГАМК (γ-аминомаслянная кислота – главный тормозный медиатор головного мозга), образующегося при декарбоксилировании глутаминовой кислоты. Поэтому нарушаются процессы торможения в ЦНС. Вследствие токсичности аммиака его концентрация в крови должна быть очень низкой – менее 65 мкмоль/л. Но при малейшем повышении его концентрации возникают симптомы аммиачного отравления: тремор, нечленораздельная речь, затуманивание зрения, возбуждение, судороги, галлюцинации. В тяжелых случаях – коматозное состояние и смерть.
В оpганизме есть системы обезвpеживания аммиака, в pезультате котоpых в кpови поддеpживается низкая концентpация аммиака. Условно выделяют местные (вpеменное связывание) и общие (конечные) механизмы обезвpеживания аммиака.
Местное обезвpеживание аммиака происходит за счет:
образование амидов аминокислот;
восстановительного аминирования α-кетоглутарата;
глюкозо-аланинового цикла. Общее (конечное) обезвреживание аммиака – за счет:
синтеза мочевины в печени;
образования солей аммония в почках.
Местное обезвpеживание аммиака:
Образование амидов аминокислот - связывание аммиака с глутаминовой и pеже аспаpагиновой кислотами с обpазованием соответствующих амидов - глутамина и аспаpагина (фермент глутаминсинтетаза):

Глутамин является нетоксической транспортной формой аммиака и его концентрация в крови значительно выше концентраций других аминокислот. Далее безвредный глутамин, легко прникающий через все биологические мембраны, секретируется в кровь и транспортируется в печень и почки, где под действием фермента – глутаминазы из глутамина высвобождается аммиак. Синтез аспарагина является менее энергетически выгоден, так как при этом затрачивается 2 макроэргические связи.
Восстановительное аминирование α-кетоглутарата – реакция, по сути, обратная реакции окислительного дезаминирования, однако, в качестве кофермента используется НАДФН:

Реакции протекают практически во всех тканях, кроме мышечной, но имеет небольшое значение, т.к. для глутаматдегидрогеназы предпочтительным субстратом является глутаминовая кислота и равновесие реакции сдвинуто в сторону α-кетоглутарата,
Цикл аланин-глюкоза. В мышцах основным акцептором лишнего аминного азота является пируват. При катаболизме белков в мышцах происходят реакции трансаминирования аминокислот, образуется глутамат, который далее передает аминоазот на пируват и образуется аланин. Из мышц с кровью аланин переносится в печень, где в обратной реакции передает свою аминогруппу на глутамат. Образующийся пируват используется как субстрат в реакциях синтеза глюкозы, а глутаминовая кислота дезаминируется и аммиак используется в синтезе мочевины.
Цикл аланин-глюкоза:

Глюкозо-аланиновый цикл выполняет следующие функции:
обезвреживает аммиак в мышцах;
транспортирует аммиак в форме аланина из мышц;
обеспечивает глюкозой мышцы.