

Регуляция
Карнитинацилтрансфераза I - регуляторный фермент β-окисления: активаторы
- АДФ, АМФ, ацил-КоА
ингибиторы - АТФ, малонил-КоА (печень).
Регуляция окисления ВЖК устроена так, что процесс окисления происходит только при недостатке энергии. Скорость окисления ВЖК зависит от скорости их транспорта из цитозоля в митохондрии, которой контролируется регуляторным ферментом – карнитинацилтрансферазой I и зависит от соотношения количества малонил-КоА/ацил-КоА. Чем выше в клетке концентрация малонил-КоА, тем ниже скорость переноса жирных кислот в матрикс митохондрий, так как малонил-КоА — аллостерический ингибитор карнитинацилтрансферазы I, а ацил-КоА —его активатор. Ингибированием карнитинацилтрансферазы I малонил-СоА обеспечивает подавление окисления жирных кислот каждый раз, когда в печень поступает слишком много глюкозы, из избытка которой активно синтезируются ТАГ.
Два фермента β-окисления: β-оксиацил-СоА дегидрогеназа и тиолаза ингибируются по типу отрицательной обратной связи: избыток НАДН -ингибитор βоксиацил-СоА дегидрогеназы, а высокие концентрации ацетил-СоА ингибируют тиолазу.
Катаболизм жирных кислот в мышцах регулируется соотношением АТФ/АДФ, т.е. при повышении АТФ в клетке активность карнитинацилтрансферазы I снижается.
Жирные кислоты не поступают в митохондрии и β-окисления не происходит.
Энергетический эффект окисления насыщенной жирной кислоты.
Связь с ЦТК и ЦПЭ
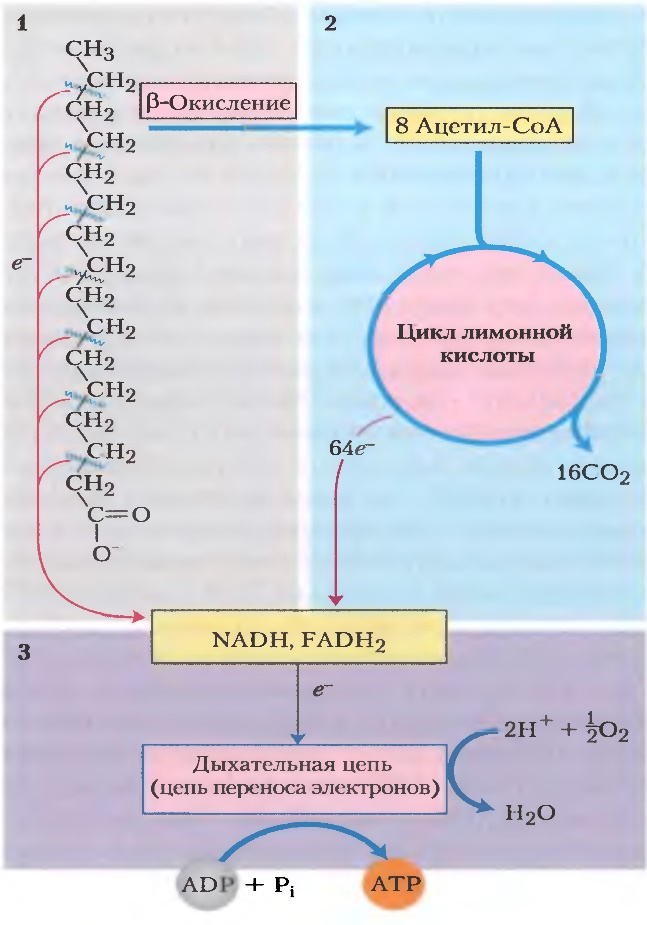
Митохондиральное окисление жирных кислот протекает в три стадии:
н
 а
первой стадии происходит – окисление,
где жирные кислоты подвергаются
последовательному окислительному
отщеплению двухуглеродных фрагментов
в виде АсСоА.
а
первой стадии происходит – окисление,
где жирные кислоты подвергаются
последовательному окислительному
отщеплению двухуглеродных фрагментов
в виде АсСоА.
на второй стадии окисления АсСоА окисляются до СО2 в ЦТК, который также протекает в матриксе.
Восстановленные эквиваленты (НАДН и ФАД.Н2), полученные на первой стадии, далее окисляются в цепи переноса электронов.
За один цикл β-окисления образуются:
1 моль ацетил-КоА, окисление которого в цитратном цикле обеспечивает синтез 12 моль АТФ.
1 моль FADH2 и 1 моль NADH, окисление которых в дыхательной цепи дает 2 и 3 моль АТФ соответственно.
При окислении пальмитиновой кислоты проходит 7 циклов β-окисления и образуется 8 моль ацетил-КоА, 7 моль FADH2 и 7 моль NADH+Н+. Следовательно, выход АТФ составляет 35 АТФ при β-окислении и 96 АТФ в результате цитратного цикла, что в сумме дает 131 моль АТФ. Поскольку активация ЖК требует затраты 1 АТФ, то из 131 следует вычесть 1 моль АТФ.
Общая формула для подсчета выхода АТФ при полном окислении насыщенных ВЖК записывается следующим образом:

где n — число С атомов в молекуле ВЖК;
n/2 — 1 — число циклов β-окисления;
5 — выход АТФ в одном цикле β-окисления;
n/2 — число ацетильных остатков;
10 — выход АТФ при полном окислении ацетил-КоА в цитратном цикле до СО2 и Н2О;
-1АТФ – затрачена на активацию ацил-КоА.
28. Биосинтез жирных кислот: последовательность реакций, регуляция, зависимость от ритма питания, биологическая роль.
Образование субстратов для биосинтеза высших жирных кислот. Субстратами для синтеза ЖК являются продукты катаболизма глюкозы, поэтому синтез ЖК происходит при высокой концентрации глюкозы в крови в абсорбтивный период, в основном, в печени, реже в жировой ткани. В клетках, где происходит синтез ЖК, активируются гликолиз и пентозофосфатный путь катаболизма глюкозы, в результаты образуются субстраты для синтеза ЖК: ацетил-КоА, NADPH, АТФ. Синтезированные ЖК включаются в состав жиров и фосфолипидов. Синтез ЖК и жиров называется липогенезом.
Превращение ацетил-КоА в малонил-КоА:

Последующие реакции катализируются ферментным комплексом – синтазой жирных кислот или пальмитатсинтазой. Конечным продуктом является пальмитиновая кислота. Синтез ЖК начинается с переноса ацетильного,а затем малонильного остатков с помощью ферментов ацетилтрансацилазы и малонилтрансацилазы на активные центры синтазы ЖК (рис. 8.8 реакции 1,2). Далее карбоксильная группа малонила выделяется в виде CO2 и по освободившейся валентности присоединяется ацетил с образованием ацетоацетил-Е. Последующие реакции восстановления,дегидратации,восстановления (реакции 4-6) приводят к образованию радикала бутирила,связанного с ферментом. Затем повторяется такой же цикл реакций и образуется радикал ЖК с 6 углеродными атомами.Цикл повторяется до образования радикала пальмитиновой кислоты, который ферментом тиоэстеразой отщепляется от ферментного комплекса. Процесс синтеза ЖК цикличен,в каждом цикле радикал ЖК увеличивается на два атома углерода,их источник – малонил-КоА. (рис.8.8. 8.9). Реакции восстановления обеспечивают синтез насыщенного алифатического радикала ЖК.
