

Строение и химический состав бактериальной клетки (и. В. Асеева)
Общая схема строения бактериальной клетки показана на рисунке 2. Внутренняя организация бактериальной клетки сложна. Каждая систематическая группа микроорганизмов имеет свои специфические особенности строения.
Клеточная стенка. Клетка бактерий одета плотной оболочкой. Этот поверхностный слой, расположенный снаружи от цитоплазматической мембраны, называют клеточной стенкой (рис. 2, 14). Стенка выполняет защитную и опорную функции, а также придает клетке постоянную, характерную для нее форму (например, форму палочки или кокка) и представляет собой наружный скелет клетки. Эта плотная оболочка роднит бактерии с растительными клетками, что отличает их от животных клеток, имеющих мягкие оболочки. Внутри бактериальной клетки осмотическое давление в несколько раз, а иногда и в десятки раз выше, чем во внешней среде. Поэтому клетка быстро разорвалась бы, если бы она не была защищена такой плотной, жесткой структурой, как клеточная стенка.
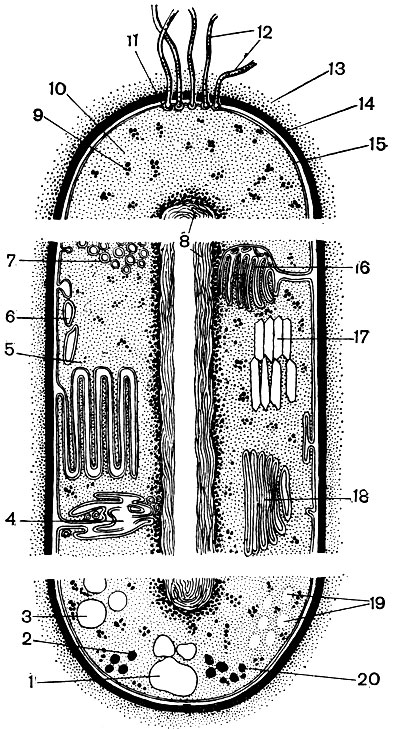 Рис.
2. Схематическое изображение строения
бактериальной клетки (по Г. Шлегелю): 1
- гранулы полироксимасляной кислоты; 2
- жировые капельки; 3 - включения серы; 4
- трубчатые тилакоиды; 5 - пластинчатые
тилакоиды; 6 - пузырьки; 7 - хроматофоры;
8 - ядро (нуклеоид); 9 - рибосомы; 10 -
цитоплазма; 11 - базальное тельце; 12 -
жгутики; 13 - капсула; 14 - клеточная стенка;
15 - цитоплазматическая мембрана; 16 -
мезосома; 17 - газовые вакуоли; 18 -
ламеллярные структуры; 19 - гранулы
полисахарида; 20 - гранулы полифосфата.
Основные структуры бактериальной клетки
представлены в верхней части рисунка,
дополнительные, мембранные структуры,
имеющиеся у фототрофных и нефототрофных
бактерий, - в средней части, а включения
запасных веществ - в нижней части
Рис.
2. Схематическое изображение строения
бактериальной клетки (по Г. Шлегелю): 1
- гранулы полироксимасляной кислоты; 2
- жировые капельки; 3 - включения серы; 4
- трубчатые тилакоиды; 5 - пластинчатые
тилакоиды; 6 - пузырьки; 7 - хроматофоры;
8 - ядро (нуклеоид); 9 - рибосомы; 10 -
цитоплазма; 11 - базальное тельце; 12 -
жгутики; 13 - капсула; 14 - клеточная стенка;
15 - цитоплазматическая мембрана; 16 -
мезосома; 17 - газовые вакуоли; 18 -
ламеллярные структуры; 19 - гранулы
полисахарида; 20 - гранулы полифосфата.
Основные структуры бактериальной клетки
представлены в верхней части рисунка,
дополнительные, мембранные структуры,
имеющиеся у фототрофных и нефототрофных
бактерий, - в средней части, а включения
запасных веществ - в нижней части
Толщина клеточной стенки 0,01 - 0,04 мкм. Она составляет от 10 до 50% сухой массы бактерий. Количество материала, из которого построена клеточная стенка, изменяется в течение роста бактерий и обычно увеличивается с возрастом.
Основным структурным компонентом стенок, основой их жесткой структуры почти у всех исследованных до настоящего времени бактерий является муреин (гликопептид, мукопептид). Это органическое соединение сложного строения, в состав которого входят сахара, несущие азот, - аминосахара и 4 - 5 аминокислот. Причем аминокислоты клеточных стенок имеют необычную форму (D-стереоизомеры), которая в природе редко встречается.
Составные части клеточной стенки, ее компоненты, образуют сложную прочную структуру (рис. 3, 4 и 5).
С помощью способа окраски, впервые предложенного в 1884 г. Кристианом Грамом, бактерии могут быть разделены на две группы: грамположительные и грамотрицательные. Грамположительные организмы способны связывать некоторые анилиновые красители, такие, как кристаллический фиолетовый, и после обработки иодом, а затем спиртом (или ацетоном) сохранять комплекс иод-краситель. Те же бактерии, у которых под влиянием этилового спирта этот комплекс разрушается (клетки обесцвечиваются), относятся к грамотрицательным.
Химический состав клеточных стенок грам - положительных и грамотрицательных бактерий различен.
У грамположительных бактерий в состав клеточных стенок входят, кроме мукопептидов, полисахариды (сложные, высокомолекулярные сахара), тейхоевые кислоты (сложные по составу и структуре соединения, состоящие из Сахаров, спиртов, аминокислот и фосфорной кислоты). Полисахариды и тейхоевые кислоты связаны с каркасом стенок - муреином. Какую структуру образуют эти составные части клеточной стенки грамположительных бактерий, мы пока еще не знаем. С помощью электронных фотографий тонких срезов (слоистости) в стенках грамположительных бактерий не обнаружено. Вероятно, все эти вещества очень плотно связаны между собой.
 Рис.
3. Схематическое изображение структуры
глико-пептида клеточной стенки. К
полисахарщщым цепочкам, состоящим из
N-ацетилглюкозамина и N-ацетилмурамовой
кислоты, присоединены цепочки из
аминокислотных единиц - пептиды. Пептиды,
связанные друг с другом пентаглициновыми
мостиками, образуют поперечные связи
полисахаридных цепочек
Рис.
3. Схематическое изображение структуры
глико-пептида клеточной стенки. К
полисахарщщым цепочкам, состоящим из
N-ацетилглюкозамина и N-ацетилмурамовой
кислоты, присоединены цепочки из
аминокислотных единиц - пептиды. Пептиды,
связанные друг с другом пентаглициновыми
мостиками, образуют поперечные связи
полисахаридных цепочек
Стенки грамотрицательных бактерий более сложные по химическому составу, в них содержится значительное количество липидов (жиров), связанных с белками и сахарами в сложные комплексы - липопротеиды и липополисахариды. Муреина в клеточных стенках грамотрицательных бактерий в целом меньше, чем у грамположительных бактерий. Структура стенки грамотрицательных бактерий также более сложная. С помощью электронного микроскопа было установлено, что стенки этих бактерий многослойные (рис. 6).
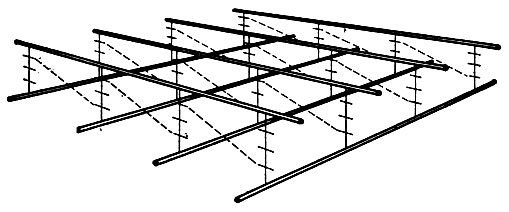 Рис.
4. Расположение макромолекул гликопептида
клеточной стенки. Жирными линиями
показаны скелеты полисахаридных цепей,
расположенные ближе к поверхности;
более тонкими - подстилающие их
полисахаридные цепи. Линии с поперечными
штрихами изображают пептидные цепочки,
связывающие эти полисахаридные цепи.
Межпептидные мостики, состоящие из пяти
остатков глицина, обозначены пунктирными
линиями
Рис.
4. Расположение макромолекул гликопептида
клеточной стенки. Жирными линиями
показаны скелеты полисахаридных цепей,
расположенные ближе к поверхности;
более тонкими - подстилающие их
полисахаридные цепи. Линии с поперечными
штрихами изображают пептидные цепочки,
связывающие эти полисахаридные цепи.
Межпептидные мостики, состоящие из пяти
остатков глицина, обозначены пунктирными
линиями
Внутренний слой состоит из муреина. Над ним находится более широкий слой из неплотно упакованных молекул белка. Этот слой в свою очередь покрыт слоем липополисахарида. Самый верхний слой состоит из липопротеидов.
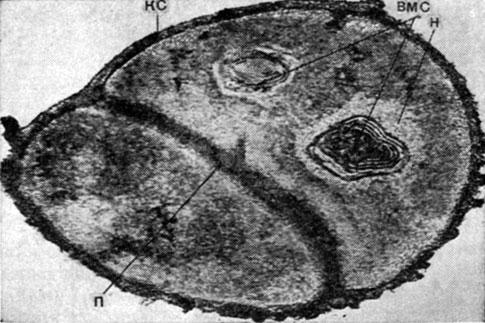 Рис.
5. Тонкое строение клеток грамположительных
кокков - Micrococcus aurantiacus. кс - клеточная
стенка; п - перегородка; вмс -
внутри-цитоплазматические мембранные
структуры; н - нуклеоид. Увел. х 65 000
Рис.
5. Тонкое строение клеток грамположительных
кокков - Micrococcus aurantiacus. кс - клеточная
стенка; п - перегородка; вмс -
внутри-цитоплазматические мембранные
структуры; н - нуклеоид. Увел. х 65 000
Клеточная стенка проницаема: через нее питательные вещества свободно проходят в клетку, а продукты обмена выходят в окружающую среду. Крупные молекулы с большим молекулярным весом не проходят через оболочку.
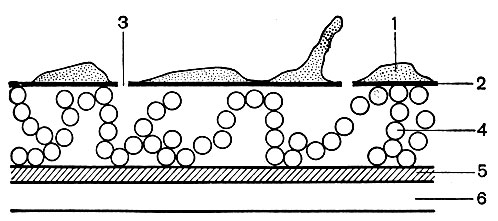 Рис.
6. Схематическое изображение клеточной
стенки грамотрицательных бактерий
- Bacterium coli (по Роузу): 1 - липопротеидный
слой с выступами и бугорками; 2 -
липо-полисахаридный слой; 3 - каналы; 4 -
рыхлоупакованные молекулы белка; 5 -
гликопептидный слой; 6 - цитоплазматическая
мембрана
Рис.
6. Схематическое изображение клеточной
стенки грамотрицательных бактерий
- Bacterium coli (по Роузу): 1 - липопротеидный
слой с выступами и бугорками; 2 -
липо-полисахаридный слой; 3 - каналы; 4 -
рыхлоупакованные молекулы белка; 5 -
гликопептидный слой; 6 - цитоплазматическая
мембрана
Капсула. Клеточная стенка многих бактерий сверху окружена слоем слизистого материала - капсулой (рис. 7). Толщина капсулы может во много раз превосходить диаметр самой клетки, а иногда она настолько тонкая, что ее можно увидеть лишь через электронный микроскоп, - микрокапсула.
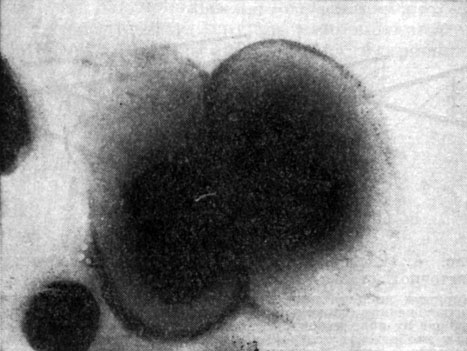 Рис.
7. Капсула клеток Azotobacter chroococcum. Увел. X
25 000
Рис.
7. Капсула клеток Azotobacter chroococcum. Увел. X
25 000
Капсула не является обязательной частью клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она служит защитным покровом клетки и участвует в водном обмене, предохраняя клетку от высыхания.
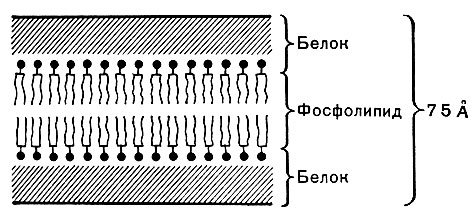 Рис.
8. Схема строения цитоплазматической
мембраны
Рис.
8. Схема строения цитоплазматической
мембраны
По химическому составу капсулы чаще всего представляют собой полисахариды. Иногда они состоят из гликопротеидов (сложные комплексы Сахаров и белков) и полипептидов (род Bacillus), в редких случаях - из клетчатки (род Acetobacter).
Слизистые вещества, выделяемые в субстрат некоторыми бактериями, обусловливают, например, слизисто-тягучую консистенцию испорченного молока и пива.
Цитоплазма. Все содержимое клетки, за исключением ядра и клеточной стенки, называется цитоплазмой. В жидкой, бесструктурной фазе цитоплазмы (матриксе) находятся рибосомы, мембранные системы, митохондрии, пластиды и другие структуры, а также запасные питательные вещества. Цитоплазма обладает чрезвычайно сложной, тонкой структурой (слоистая, гранулярная), С помощью электронного микроскопа раскрыты многие интересные детали строения клетки.
Внешний липопротеидный слой протопласта бактерий, обладающий особыми физическими и химическими свойствами, называется цнтоплазматической мембраной (рис. 2, 15).
Внутри цитоплазмы находятся все жизненно важные структуры и органеллы.
Цитоплазматическая мембрана выполняет очень важную роль - регулирует поступление веществ в клетку и выделение наружу продуктов обмена.
Через мембрану питательные вещества могут поступать в клетку в результате активного биохимического процесса с участием ферментов. Кроме того, в мембране происходит синтез некоторых составных частей клетки, в основном компонентов клеточной стенки и капсулы. Наконец, в цнтоплазматической мембране находятся важнейшие ферменты (биологические катализаторы). Упорядоченное расположение ферментов на мембранах позволяет регулировать их активность и предотвращать разрушение одних ферментов другими. С мембраной связаны рибосомы - структурные частицы, на которых синтезируется белок. Мембрана состоит из липопротеидов. Она достаточно прочна и может обеспечить временное существование клетки без оболочки. Цитоплазматическая мембрана составляет до 20% сухой массы клетки.
На электронных фотографиях тонких срезов бактерий цитоплазматическая мембрана представляется в виде непрерывного тяжа толщиной около 75А, состоящего из светлого слоя (липиды), заключенного между двумя более темными (белки). Каждый слой имеет ширину 20 - 30А. Такая мембрана называется элементарной (табл. 30, рис. 8).
Между плазматической мембраной и клеточной стенкой имеется связь в виде десмозов - мостиков. Цитоплазматическая мембрана часто дает инвагинации - впячивания внутрь клетки. Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. Некоторые виды мезосом представляют собой тельца, отделенные от цитоплазмы собственной мембраной. Внутри таких мембранных мешочков упакованы многочисленные пузырьки и канальцы (рис. 2). Эти структуры выполняют у бактерий самые различные функции. Одни из этих структур - аналоги митохондрий. Другие выполняют функции эндо-плазматической сети или аппарата Гольджи. Путем инвагинации цитоплазматической мембраны образуется также фотосинтезирующий аппарат бактерий. После впячивания цитоплазмы мембрана продолжает расти и образует стопки (табл. 30), которые по аналогии с гранулами хлоропластов растений называют стопками тилакоидов. В этих мембранах, часто заполняющих собой большую часть цитоплазмы бактериальной клетки, локализуются пигменты (бактериохлорофилл, каротиноиды) и ферменты (цитохромы), осуществляющие процесс фотосинтеза.
В цитоплазме бактерий содержатся рибосомы- белок-синтезирующие частицы диаметром 200А. В клетке их насчитывается больше тысячи. Состоят рибосомы из РНК и белка. У бактерий многие рибосомы расположены в цитоплазме свободно, некоторые из них могут быть связаны с мембранами.
Рибосомы являются центрами синтеза белка в клетке. При этом они часто соединяются между собой, образуя агрегаты, называемые полирибосомами или полисомами.
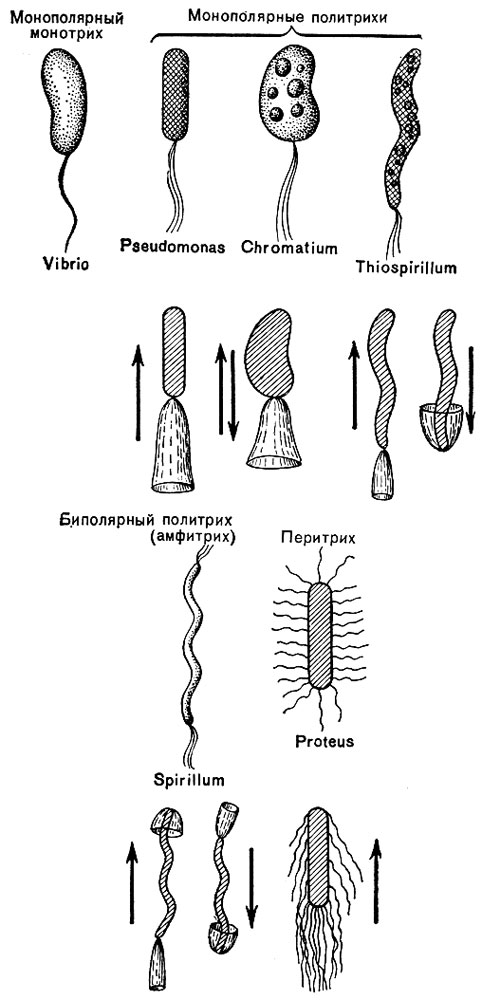 Рис.
9. Основные типы расположения жгутиков
и направления движений бактерий
Рис.
9. Основные типы расположения жгутиков
и направления движений бактерий
В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Однако их присутствие нельзя рассматривать как какой-то постоянный признак микроорганизма, обычно оно в значительной степени связано с физическими и химическими условиями среды. Многие цитоплазматические включения состоят из соединений, которые служат источником энергии и углерода. Эти запасные вещества образуются, когда организм снабжается достаточным количеством питательных веществ, и, наоборот, используются, когда организм попадает в условия, менее благоприятные в отношении питания.
У многих бактерий гранулы состоят из крахмала или других полисахаридов - гликогена и гранулезы. У некоторых бактерий при выращивании на богатой сахарами среде внутри клетки встречаются капельки жира. Другим широко распространенным типом гранулярных включений является волютин (метахроматиновые гранулы). Эти гранулы состоят из полимета-фосфата (запасное вещество, включающее остатки фосфорной кислоты). Полиметафосфат служит источником фосфатных групп и энергии для организма. Бактерии чаще накапливают волютин в необычных условиях питания, например на среде, не содержащей серы. В цитоплазме некоторых серных бактерий находятся капельки серы.
Помимо различных структурных компонентов, цитоплазма состоит из жидкой части - растворимой фракции. В ней содержатся белки, различные ферменты, т-РНК, некоторые пигменты и низкомолекулярные соединения - сахара, аминокислоты.
В результате наличия в цитоплазме низкомолекулярных соединений возникает разность в осмотическом давлении клеточного содержимого и наружной среды, причем у разных микроорганизмов это давление может быть различным. Наибольшее осмотическое давление отмечено у грамположительных бактерий - 30 атм, у грамотрицательных бактерий оно гораздо ниже - 4 - 8 атм.
Ядерный аппарат. В центральной части клетки локализовано ядерное вещество - дезоксирибонуклеиновая кислота (ДНК).
У бактерий нет такого ядра, как у высших организмов (эукариотов), а есть его аналог - "ядерный эквивалент" - нуклеоид (см. рис. 2, 8), который является эволюционно более примитивной формой организации ядерного вещества. Микроорганизмы, не имеющие настоящего ядра, а обладающие его аналогом, относятся к прокариотам. Все бактерии - прокариоты. В клетках большинства бактерий основное количество ДНК сконцентрировано в одном или нескольких местах. В клетках эукариотов ДНК находится в определенной структуре - ядре. Ядро окружено оболочкой - мембраной.
У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками - гистонами - и в нуклеоиде расположена в виде пучка фибрилл.
Жгутики. На поверхности некоторых бактерий имеются придаточные структуры; наиболее широко распространенными из них являются жгутики - органы движения бактерий.
Жгутик закрепляется под цитоплазматиче-ской мембраной с помощью двух пар дисков (см. на стр. 206). У бактерий может быть один, два или много жгутиков. Расположение их различно: на одном конце клетки, на двух, по всей поверхности и т. д. (рис. 9). Жгутики бактерий имеют диаметр 0,01 - 0,03 мкм, длина их может во много раз превосходить длину клетки. Бактериальные жгутики состоят из белка - флагеллина - и представляют собой скрученные винтообразные нити.
На поверхности некоторых бактериальных клеток имеются тонкие ворсинки - фимбрии. Подробнее они описаны на странице 205.
Химический состав бактериальной клетки сходен с химическим составом других живых организмов. Ее компонентами являются вода, минеральные и органические соединения – белки, нуклеиновые кислоты, углеводы, липиды.
Клеточная водасоставляет 75…90% массы вегетативной клетки. Нормальный метаболизм и размножение микроорганизмов возможны только в водной среде. Вода является растворителем органических и минеральных веществ, дисперсионной средой для коллоидов, источником водородных и гидроксильных ионов, а также водорода и кислорода. В клетке вода находится в двух состояниях: свободная вода, которая является растворителем, принимает участие в процессах метаболизма; связанная вода - связана с клеточными колодами, обуславливает изменение значений криоскопической точки цитоплазмы (определяет устойчивость микроорганизмов к низким и высоким температурам).
Минеральный состав. Из множества известных в настоящее время химических элементов в состав живого вещества чаще всего входит только 22, из них лишь 16 элементов встречаются во всех группах организмов. Шесть основных элементов (углерод, кислород, азот, водород, фосфор и сера) составляют 95% сухой биомассы бактериальной клетки. Углерод, азот, водород, кислород легко образуют прочные ковалентные связи посредством спаривания электронов, благодаря чему они способны давать разнообразные химические соединения. Бактерии также содержат калий, натрий, кальций, магний, хлор, железо (в пределах 1… 0,5% сухого вещества). На остальные элементы приходится около 0,3% (таблица.1.1).
Микроорганизмы используют металлы в форме катионов неорганических солей. Минеральные соли составляют от 2 до 30% сухого вещества клетки. Содержание солей варьирует в зависимости от состава питательной среды и возраста культуры. В молодых клетках их в 6…7 раз больше, чем в старых.
Таблица 1. 1 - Приблизительный элементарный состав бактериальных клеток
Элементы |
Количество, % СВ клетки |
Элементы |
Количество, % СВ клетки |
Элементы |
Количество, % СВ клетки |
Углерод |
|
Фосфор |
|
Кальций |
0,5 |
Кислород |
|
Сера |
|
Магний |
0,5 |
Азот |
|
Калий |
|
Хлор |
0,5 |
Водород |
|
Натрий |
|
Железо |
0,2 |
Некоторым бактериям для роста необходимы редкие элементы. По значению элементы можно условно разделить на существенные(отсутствие которых нарушает рост и размножение бактерий) и несущественные, которые могут быть заменены. Имеется группа олигоэлементов (греч. oligos - малый), которые в очень малых количествах стимулируют рост и развитие бактерий. Их общая масса не превышает 0,1% клеточного вещества. К ним относятся кадмий, ванадий, молибден, олово, никель, серебро и др. Эти элементы в питательные среды обычно не добавляют, так как они содержатся в водопроводной воде, пептоне, мясном экстракте и т.д.
Органические соединения. Большинство элементов находится в бактериальной клетке в составе биомолекул, среди которых аминокислоты, азотистые основания, сахара, жирные кислоты, спирты и т.д. являются основными компонентами органических веществ.
Белки составляют 40…80% клетки и представлены простыми и сложными белками. К сложным белкам относятся нуклеопротеиды, липопротеиды, гликопротеиды, фосфопротеиды, металлопротеиды (содержащие железо, цинк, медь), флавопротеиды. Содержание белков в бактериальной клетке варьирует в зависимости от вида бактерий, возраста культуры, состава питательной среды и т.д.
Бактериальные белки состоят из тех же 20 важнейших аминокислот, что и белки растений и животных, которые образуют полипептидные цепи (-СО-NН-).. Аминокислотный состав белков различных видов бактерий качественно и количественно различен. Только у бактерий встречаются такие аминокислоты, как диаминопимелиновая (ДАП), аминомасляная, гомосерин, D-аланин, D-глутаминовая, отсутствующие у высших организмов. Большинство бактерий сами синтезируют все необходимые им аминокислоты, но некоторые нуждаются в готовых аминокислотах, которые вносятся в питательные среды.
У бактерий преобладают глобулярныебелки, полипептидные цепи которых плотно свернуты в компактные сферические и глобулярные структуры. Большинство из них растворимы в водных растворах, легко диффундируют, для них характерны биологические функции (это ферменты, токсины и др.). Белки бактерий часто выполняют одновременно структурную и функциональную роль (например, белки цитоплазматической мембраны).
Нуклеиновые кислоты – сложные полимеры, состоящие из большого количества (от 1,5 тыс до 5 млн) мононуклеотидов. Мононуклеотиды построены из азотистого основания (пуринового – аденин, гуанин, или пиримидинового – цитозин, тимин или урацил), сахара (рибозы или дезоксирибозы) и остатка фосфорной кислоты. Мононуклеотиды ковалентно связываются между собой фосфодиэфирными связями, образуя полинуклеотиды – нуклеиновые кислоты .
Рибонуклеиновая кислота (РНК)составляет 10% сухого вещества, она обычно одноцепочная. В клетках существует три типа РНК: информационная, или матричная (используется в качестве матрицы, определяющей последовательность аминокислот в растущей полипептидной цепи); транспортная (переносит на рибосому определенные аминокислоты); рибосомальная (находится в рибосомах).
Дезоксирибонуклеиновая кислота (ДНК) составляет 3…4% сухого вещества, она состоит из двух полинуклеотидных цепей, образующих одну двойную спираль. Нуклеоид бактерий представлен гигантской молекулой ДНК, в которой закодирована вся генетическая информация клетки. Кроме того, у некоторых бактерий в цитоплазме находятся молекулы внехромосомной ДНК – плазмиды.
Углеводы в бактериальной клетке составляют 12…30% сухого вещества. Они представлены моно- и полисахаридами. Полисахаридыбактерий чрезвычайно разнообразная группа биополимеров. Среди них есть соединения, характерные, как для прокариот, так и эукариот (целлюлоза, гликоген). У бактерий обнаружен ряд полисахаридов, не встречающиеся у других организмов (тейхоевые кислоты, пептидогликаны, декстран и др.). Высокая активность полисахаридов обусловлена их способностью легко вступать в реакции с другими макромолекулами путем гидрофобных взаимодействий, а также соединяясь ионными и водородными связями. Большинство полисахаридов включает самые распространенные сахара – глюкозу, галактозу, рамнозу.
Тейхоевые кислоты – это полимеры глицерина (трехатомного спирта) или рибита (пятиатомного спирта) – глицеротейхоевая и рибитотейхоевая кислоты. В этих кислотах от 8 до 50 остатков спирта связано фосфодиэфирными связями и могут содержать моносахарид в качестве заместителя. Они содержатся в клеточной стенке бактерий (греч teichos – стенка).
Липополисахариды (ЛПС) состоят из липидной части и полисахаридной, которая представлена различными углеводами.
По функциональной активности полисахариды бактерий подразделяются на внутриклеточные (запасные вещества клетки, эндотоксины и пр.) и внеклеточные, которые входят в состав капсулы.
Липиды бактерий представлены нейтральными жирами, фосфолипидами, восками. Общее содержание липидов варьирует от 5% до 40%. Основная масса липидов в бактериальной клетке связана с другими компонентами (белками, полисахаридами). Липиды бактерий значительно разнообразнее липидов высших организмов. Они выполняют разнообразные функции – являются аккумуляторами энергии у некоторых бактерий (поли-β-оксимасляная кислота), служат структурными компонентами клетки (цитоплазматическая мембрана), участвуют в метаболизме углеводов, в энергетическом обмене, входят в состав антигенов, определяют кислотоустойчивость бактерий.
Нейтральные жиры (ацилглицерины, или глицериды) бактерий часто содержат насыщенные жирные кислоты. Ненасыщенные жирные кислоты представлены исключительно кислотами с одной двойной связью. Содержание свободных жирных кислот в клетках варьирует от 5 до 30% в зависимости от видовых особенностей бактерий и условий их выращивания. Качественный и количественный состав жирных кислот изменяется с возрастом культуры, характеризуется высокой чувствительностью к физическим и химическим факторам внешней среды. Так, молодые клетки содержат значительное количество ненасыщенных жирных кислот. Бактерии также способны включать в себя жирные кислоты из окружающей среды. Бактерии не содержат полиненасыщенные жирные кислоты, стерины и стероиды.
Фосфолипиды – основные представители бактериальных липидов. В бактериальной клетке их количество составляет от 0,4% до 6,5%. Основная масса фосфолипидов содержится в цитоплазматической мембране или в клеточных оболочках.
Воски содержат кислотоустойчивые бактерии, например, микобактерии.
Пигменты бактерий. Среди бактерий есть значительное число пигментообразующих видов. Пигменты вырабатываются в зависимости от условий выращивания культуры бактерий – минерального состава и реакции среды, источника углерода, температуры, количества кислорода, наличия света. Важными элементами для образования пигментов являются азот, магний, железо, кальций и др. Так, магний необходим для синтеза бактериохлорофиллов, железо – для продигиозина, пиоцианина. Пигменты могут выполнять в бактериальной клетке различные функции. Установлено их участие в фотосинтезе, в дыхании, в окислительно-восстановительных реакциях, защите от экстремальных факторов внешней среды (УФ-излучение, повышенная концентрация минеральных солей и т.д.).
По растворимости микробные пигменты можно разделить на две группы: нерастворимые (хромофорные) пигменты, связанные с клеточными компонентами и обуславливающие окраску колоний бактерий, но не среды; растворимые (хромоларные) пигменты, которые растворяются в питательной среде и окрашивают ее при росте бактерий. По химическому составу пигменты разнообразны – каротиноиды, меланины, хиноны, бактериохлорофиллы, пирролы.
Бактериохлорофиллами называются хлорофиллы, содержащиеся в клетках фотосинтезирующих бактерий. Их существует несколько типов, и они структурно отличаются как между собой, так и от хлорофилла высших растений. Большинство фотосинтезирующих бактерий содержит бактериохлорофилл а, у пурпурных бактерий обнаружен бактериохлорофилл b, у зеленых серобактерий – c,d,e.
Бактериохлорофиллы поглощают свет в более длинноволновой области (800…1100 нм), чем хлорофиллы высших растений.
Каротиноидные пигменты содержатся у всех фотосинтезирующие микроорганизмы и у ряда нефотосинтезирующих. Известно свыше 300 каротиноидов и они в определенной степени определяют окраску колоний.
Меланиновые пигменты (черного и коричневого цвета) – это тесно связанные с белками клетки биополимеры, которые обуславливают защиту клетки от экстремальных окислительных и восстановительных условий.