

Активирует цАМФ-фосфодиэстеразу, снижая тем самым ее активирующее
влияние на протеинкиназу А и реакции катаболизма.
Медленные эффекты (минуты-часы)
Активация синтеза глюкокиназы, АТФ-цитрат-лиазы, ацетил-S-КоА-карбо-
ксилазы, синтазы жирных кислот, пируваткиназы, люкозо-6-фосфатдегидрогеназы, цитолозольной малатдегидрогеназы.;
Увеличение синтеза тРНК для увеличения скорости транскрипции. Однако на-работку мРНК антагонистических ферментов снижает (например, для ФЕП-
карбоксикиназы);
Повышает фосфорилирование по серину рибосомального белка S6, что сти-мулирует синтез белка.
Очень медленные эффекты (часы-сутки)
Повышает синтез соматомедина, зависимый от гормона роста;
Увеличивает рост и пролиферацию клеток, действуя при этом синергично с
соматомедином;
Стимулирует тирозиновые киназы., вызывает переход клетки из G1 в S-фазу клеточного цикла.
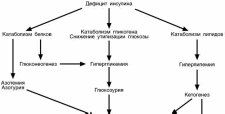
ОСЛЕДСТВИЯ ДЕФИЦИТА ИНСУЛИНА
Быстрые последствия
Гипергликемия – так как отсутствует влияние инсулина и превалирует влия-
ние глюкагона, адреналина, кортизола, гормона роста.
Глюкозурия – почечный порог для глюкозы, т.е. концентрация глюкозы в кро-
ви при которой она появляется в моче, примерно равен 10,0 ммоль/л. В норме в мо-
че уровень глюкозы 0,8 ммоль/л и до 2,78 ммоль/сут, в других единицах около 0,5
г/сут, при СД количество теряемой глюкозы составляет до 100 г/сут и более.
Преобладание катаболизма белков над анаболизмом ведет к накоплению
продуктов азотистого об-
мена, в первую очередь мочевины и ее повышен-
ному выведению. Избыток
аминокислот уходит в глю-
конеогенез.
Глюкоза и мочевина
осмотически удерживают воду в просвете поччного
канальца и возникает по-
лиурия. Объем мочи воз-растает в 2-3 раза.
Повышенный распад
ТАГ в жировой ткани и пе-чени обуславливает ано-
мально высокое окисление
жирных кислот и накопле-ние их недоокисленных
продуктов – кетоновых тел.
Это приводит к кетонемии,
кетонурии и кетоацидозу.
При диабете концентрация кетоновых тел возрастает
100-200 раз и достигает
350 мг% (норма 2 мг% или
0,1-0,6 ммоль/л).
При полиурии с мо-
чой, кроме воды, теряются соли, в частности карбо-
наты, имеющие щелочной характер. Это усугубляет ацидоз.
В результате
.п.п.4,5,6 возникает дегид-
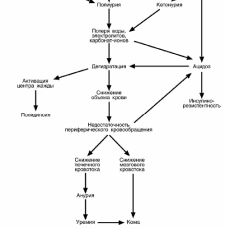
ратация (в тяжелых случаях до 5 л) организма, которая заключается в падении объ-
ема крови, обезвоживанию клеток и их сморщиванию (дряблая кожа, запавшие гла-
за, мягкие глазные яблоки, сухость слизистых), уменьшению артериального давле-ния. Ацидоз вызывает одышку (дыхание Kussmaul, быстрое и глубокое) и дополни-
тельную дегидратацию.
Активируется центр жажды и начинается полидипсия.
Дегидратация неминуемо приводит к недостаточности кровообращения в тка-
нях – активируется анаэробный гликолиз, накапливается лактат и в дополнение к ке-
тоацидозу возникает лактацидоз.
Закисление среды вызывает изменение взаимодействия инсулина с рецепто-
рами, клетки становятся нечувствительными к инсулину – инсулинорезистент-
ность.
Ацидоз крови уменьшает концентрацию 2,3-дифосфоглицерата в эритроци-
тах. Это, повышая сродство гемоглобина к кислороду, создает тканевую гипоксию и
усугубляет лактацидоз
Отдаленные последствия
Гипергликемия резко повышает потребление глюкозы инсулиннезависимыми тканями (клетки артериальных стенок, эндотелия, клетки Шванна, эритроциты, хру-
сталик и сетчатка глаза, семенники и гломерулярные клетки почек), в них вынужден-но активируется особые пути метаболизма глюкозы. Их интенсивность определяется только доступностью глюкозы:
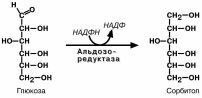
Превращение глюкозы в
сорбитол.
Сорбитол плохо проникает через клеточные мембраны, его накопление в цитозоле приводит
осмотическому набуханию кле-ток и нарушению их функций. Например, возникновение ката-ракты хрусталика и нейропатий (нарушение осязания) в клетках Шванна
Неферментативное гли-козилирование различных бел-
ков, изменение их свойств и активация их синтеза за счет избытка энергии:
увеличивается синтез гликопротеинов базальной мембраны почечных клу-бочков. Это приводит к окклюзии капилляров и нарушению фильтрации
увеличивается синтез гликопротеинов в сетчатке глаза, что вызывает отек
сетчатки и кровоизлияния
увеличивается синтез гликопротеинов в стекловидном теле
увеличивается синтез тканевых белков за счет доступности глюкозы и энер-
гии
гликозилированные белки хрусталика объединяются в крупные агрегаты,
рассеивающие свет. Это вызывает помутнение хрусталика и катаракту.
гликозилирование гемоглобина в эритроцитах, образование HbA1C
белков свертывающей системы, что увеличивает вязкость крови
белков ЛПНП, что уменьшает их связывание с рецепторами и повышает концентрацию ХС в крови
белков ЛПВП, что усиливает их сродство к рецепторам и быструю элимина-
цию из кровотока
Из-за двух последних нарушений возникают макроангиопатии развивается ате-
росклероз сосудов мозга, сердца, почек, конечностей. Характерно в основном для
ИНЗСД.
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ
При калорийности рациона 2000–3000 ккал суточное потребление углеводов составляет 300-450г. С пищей поступает крахмал, сахароза, лактоза, пищевые волокна (клетчатка и др.). Переваривание углеводов начинается в ротовой полости при участии α-амилазы слюны, которая расщепляет в крахмале α-1,4-гликозидные связи. Полное расщепление крахмала здесь не происходит, так как пребывание пищи во рту кратковременно. Из крахмала в ротовой полости образуются крупные фрагменты – декстрины. Желудочный сок не содержит ферментов, расщепляющих углеводы. Дальнейшее переваривание углеводов происходит в тонком кишечнике. Фермент поджелудочной железы α-амилаза расщепляет α-1,4-гликозидные связи крахмала и декстринов, α-1,6-гликозидные связи расщепляются ферментом кишечного сока – амило-1,6-гликозидазой. При действии двух ферментов образуется дисахарид мальтоза. Амилаза поджелудочной железы не расщепляет β-1.4-гликозидные связи, которыми соединены остатки глюкозы в молекуле целлюлозы. Поэтому пищевые волокна не перевариваются, но они должны присутствовать в рационе, так как улучшают перистальтику, ускоряют чувство насыщения и снижают уровень холестерина в крови, поскольку на них происходит адсорбция желчных кислот и выведение их из организма. Пищевые волокна обязательно должны присутствовать в рационе при ожирении, запорах, атеросклерозе, сахарном диабете. Мальтоза, образовавшаяся из крахмала, а также дисахариды пищи – сахароза и лактоза перевариваются ферментами тонкого кишечника – дисахаридазами. Эти ферменты работают не в просвете кишечника, а на поверхности эпителиальных клеток кишечника. Мальтоза расщепляется мальтазой до 2 молекул глюкозы, лактоза – лактазой до глюкозы и галактозы, сахароза – сахаразой до глюкозы и фруктозы (рис.3). Все моносахариды всасываются, сначала путем облегченной диффузии, а затем активным транспортом в симпорте с ионами Nа+.
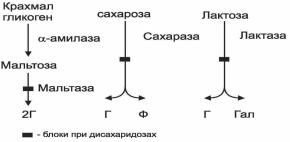
Рис.3. Катаболизм дисахаридов и патогенез дисахаридозов
В крови воротной вены содержатся три моносахарида: глюкоза, фруктоза и галактоза. Все они попадают в печень, где происходит унификация фруктозы и галактозы, т.е. они превращаются в глюкозу – единственный моносахарид, используемый всеми клетками нашего организма.
Дисахаридозы – нарушение переваривания дисахаридов, связанные с недостаточной активностью дисахаридаз. Недостаточная активность ферментов может быть врожденной и приобретенной. Симптомы врожденных форм проявляются достаточно рано, например, после первого кормления грудным молоком (при дефиците лактазы) или при добавлении в рацион сахара или крахмала. Приобретенные формы могут наблюдаться при заболеваниях кишечника. Нерасщепленные дисахариды вызывают осмотическую диарею, сбраживаются микрофлорой кишечника с образованием углекислого газа, что приводит к метеоризму, коликам.
ОБМЕН ГЛИКОГЕНА
Многие ткани в качестве резервной формы глюкозы синтезируют гликоген. Синтез и распад гликогена обеспечивают постоянство концентрации глюкозы в крови. Синтезгликогена происходит в покое и сытости, как любой анаболический процесс требует энергии. Депонируется гликоген главным образом в печени и мышцах. Глюкоза, поступившая в клетку, фосфорилируется при участии гексокиназы за счет АТФ, при этом образуется глюкозо-6-фосфат, который в ходе обратимой реакции под действием фосфоглюкомутазы превращается в глюкозо-1-фосфат. Затем при участии УТФ глюкозо-1-фосфат превращается в УДФ-глюкозу. Эта молекула используется как донор остатков глюкозы при синтезе гликогена.
Так как гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путем удлинения уже имеющейся молекулы полисахарида, называемой «затравка». К «затравке» последовательно присоединяются остатки глюкозы из УДФ-глюкозы α- 1,4-гликозидной связью при участии фермента гликогенсинтазы. Разветвленная структура гликогена образуется при участии «фермента ветвления» (рис.4). Регуляторными ферментами в синтезе гликогена являются гликогенсинтаза и гексокиназа. Синтез гликогена увеличивается под влиянием инсулина, а тормозится глюкагоном, катехоламинами, глюкокортикостероидами.
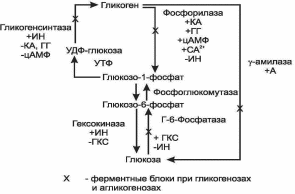
Рис.4. Обмен гликогена печени
Распад гликогена происходит путем последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата. Гликозидная связь расщепляется с присоединением неорганического фосфата, поэтому процесс называется фосфоролизом, а фермент – фосфорилазой. Образовавшийся глюкозо-1-фосфат затем изомеризуется фосфоглюкомутазой до глюкозо-6-фосфата. В печени (но не в мышцах) глюкозо-6-фосфат может гидролизоваться с образованием глюкозы, которая выделяется в кровь. Эту реакцию катализирует глюкозо-6-фосфатаза. Мышечный гликоген не используется для поддержания уровня глюкозы в крови, так как в мышцах нет фермента глюкозо-6-фосфатазы и образование свободной глюкозы там невозможно, а глюкозо-6-фосфат не может проникать через мембрану клеток. Таким образом, печень запасает глюкозу в виде гликогена не столько для собственных нужд, сколько для поддержания постоянной концентрации глюкозы в крови. Функция мышечного гликогена заключается в освобождении глюкозо-6-фосфата, потребляемого в самой мышце для окисления и использования энергии.
Регуляторными ферментами распада гликогена являются фосфорилаза и глюкозо-6-фосфатаза. Процесс распада усиливают катехоламины, глюкагон, глюкокортикостероиды; тормозит инсулин.
Распад глюкозы в аэробных и анаэробных условиях
Катаболизм глюкозы – основной поставщик энергии для большинства клеток организма.
Аэробный распад протекает в условиях достаточного снабжения тканей кислородом. Он включает несколько стадий:
1. Аэробный распад глюкозы до двух молекул пирувата в гиалоплазме клеток,который можно разделить на 2 этапа:
а)подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз.
б) основной этап, сопряженный с синтезом АТФ. В результате серии реакций фосфотриозы превращаются в пируват.
Превращение пирувата в ацетил-КоА и окисление последнего в цикле Кребса
Дыхательная цепь
Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с учас-тием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Это обратимая реакция изомеризации протекает при участии глюкозофосфатизомеразы. Затем следует еще одна реакция фосфорилирования за счет АТФ, катализирует ее фосфофруктокиназа. В ходе этой реакции фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат. Этот метаболит далее расщепляется на 2 триозофосфата: фосфоглицериноый альдегид (ФГА) и диоксиацетонфосфат (ДОАФ), реакцию катализирует альдолаза. В последующих реакциях гликолиза используется только ФГА, поэтому ДОАФ превращается с участием фермента триозофосфатизомеразы в ФГА. На этом подготовительный этап заканчивается.
Основной этап аэробного распада включает реакции, связанные с синтезом АТФ. Сна-чала происходит окисление ФГАпри участии ФГА-дегидрогеназы. Этот фермент является НАД+-зависимымым. Восстановленный НАД+ затем окисляется в дыхательной цепи (перенос НАДН в митохондрии, где находится дыхательная цепь, происходит при участии челночных механизмов, это связано с тем, что митохондриальная мембрана непроницаема для НАДН). Кроме того, свободная реакция окисления этой реакции концентрируется в макроэргической связи продукта реакции. Продуктом реакции является 1,3-бисфосфоглицерат. Для фосфорилирования используется Н3РО4. В следующей реакции высокоэнергетический фосфат передается на АДФ с образованием АТФ. Катализирует реакцию фосфоглицераткиназа. АТФ в данной реакции образуется путем субстратного фосфорилирования. Продуктом реакции является 3-фосфоглицерат. В следующей реакции происходит перенос фосфата из 3 положения во второе с образованием 2-фосфоглицерата. Из него при отщеплении воды образуется фосфоенолпируват – макроэргическое соединение, фосфатная группа которого в следующей реакции переносится на АДФ с образованием АТФ при участии пируваткиназы (это вторая реакция субстратного фосфорилирования). Продуктом реакции является пируват.
Во второй стадии пируват превращается в ацетил-КоА, который сгорает в цикле Кребса и дыхательной цепи (третья стадия) до СО2 и Н2О.
Всего при аэробном окислении глюкозы выделяется 32 АТФ.
Анаэробный гликолиз представляет собой распад глюкозы в анаэробных условиях (условиях недостаточного снабжения кислородом), поэтому он не зависит от работы митохондриальной дыхательной цепи. АТФ в гликолизе образуется путем субстратногофосфорилирования. Конечным продуктом является лактат (рис. 5).
Все реакции анаэробного гликолиза (11 реакций) протекают в гиалоплазме. Первые 10 реакций до пирувата идентичны аэробному распаду. В последней реакции происходит восстановление пирувата в лактат цитозольным НАДН. Реакцию эту катализирует лактатдегидрогеназа. С помощью этой реакции обеспечивается регенерация НАД+ из НАДН без участия дыхательной цепи в ситуациях, связанных с недостаточным снабжением клеток кислородом. Роль акцептора водорода от НАДН (подобно кислороду в дыхательной цепи) выполняет пируват. Таким образом, значение реакции восстановления пирувата заключается не в образовании лактата, а в том, что данная цитозольная реакция обеспечивает регенерацию НАД+. К тому же лактат не является конечным продуктом метаболизма, удаляемым из организма (лактат – тупиковый метаболит). Он выводится в кровь и утилизируется: либо превращается в глюкозу (75%), либо при доступности кислорода окисляется до СО2 и Н2О (25%).
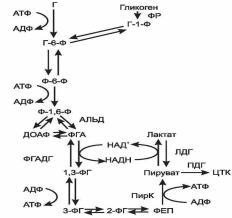
Рис.5. Гликолиз
Энергетический выход анаэробного гликолиза составляет 2 АТФ.
Недостатками анаэробного гликолиза являются: малый энергетический выход по сравнению с аэробным распадом и накопление лактата, что приводит к ацидозу, усталости.
Достоинства гликолиза: он полезен при выполнении кратковременной работы, является единственным источником энергии для эритроцитов (в них отсутствуют митохондрии), а также необходим в разных органах при дефиците кислорода (наложение жгута, падение давления, нарушение кровотока).
Регуляция.Регуляторными ферментами гликолиза являются: гексокиназа, фосфофруктокиназа и пируваткиназа. Аллостерическими ингибиторами являются АТФ и цитрат. Стимулирует гликолиз инсулин, а тормозят глюкагон, глюкокортикостероиды. Катехоламины тормозят гликолиз в печени и активируют в мышцах.
ГЛЮКОНЕОГЕНЕЗ
Некоторые ткани нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно, содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счет расщепления гликогена в печени. Однако запасы гликогена в печени невелики, они практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается глюконеогенез (ГНГ) – процесс синтеза глюкозы из веществ неуглеводной природы. Его основной функцией является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. В первую очередь это важно для головного мозга (он не может обеспечивать потребности в энергии за счет окисления жирных кислот) и эритроцитов – в них аэробный путь распада веществ невозможен из-за отсутствия митохондрий. Процесс в основном (на 90%) протекает в печени и менее интенсивно в корковом веществе почек и слизистой тонкого кишечника.
Субстратами ГНГ являются лактат, аминокислоты, глицерин. Аминокислоты включаются в ГНГ при длительном голодании или продолжительной мышечной работе. Глицерин освобождается в процессе липолиза из жировой ткани в период голодания или при
длительной физической нагрузке. Лактат – продукт анаэробного гликолиза. Его источником в покое являются эритроциты, а при работе – мышцы. Таким образом, лактат используется в глюконеогенезе постоянно. Большинство реакций глюконеогенеза протекает за счет обратимых реакций гликолиза и катализируется теми же ферментами. Однако три реакции гликолиза необратимы. На этих стадиях реакции ГНГ протекают другими путями (обходные пути ГНГ).
Первая необратимая реакция – образование из пирувата фосфоенолпирувата . В ГНГ это происходит в ходе двух реакций. Пируват в митохондриях карбоксилируется с образованием оксалоацетата при участии пируваткарбоксилазы – биотинзависимого фермента. Реакция протекает с использованием энергии АТФ. Дальнейшие превращения оксалоацетата протекают в цитозоле. Фосфоенолпируваткарбоксикиназа с затратой энергии ГТФ превращает оксалоацетат в фосфоенолпируват. Дальнейшие реакции ГНГ вплоть до образования фруктозо-1,6-бисфосфата протекают в цитозоле и катализируются гликолитическими ферментами. Затем следует еще одна необратимая реакция ГНГ, катализируемая фруктозо-1,6-бисфосфатазой, в которой происходит отщепление остатка фосфорной кислоты гидролитическим путем. Образующийся при этом фруктозо-6-фосфат гликолитическим ферментом изомеризуется до глюкозо-6-фосфата, от которого отщепляется остаток фосфорной кислоты при участии еще одного необратимо работающего фермента – глюкозо-6-фосфатазы. Образовавшаяся свободная глюкоза из клетки выходит в кровь. В ходе ГНГ расходуется 6 моль АТФ на синтез 1 моль глюкозы из пирувата или лактата.
Ключевыми ферментами ГНГ, катализирующими необратимые обходные реакции, являются: пируваткарбоксилаза, фосфоенолпируваткарбоксикиназа, фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза. Активируют ГНГ аллостерически АТФ и цитрат, обязательным активатором первого фермента является ацетил-КоА. ГНГ усиливают гормоны: глюкагон, глюкокортикостероиды. Тормозит ГНГ гормон покоя и сытости – инсулин.
Лактат, образовавшийся в ходе анаэробного гликолиза, не является конечным продуктом метаболизма. Использование лактата связано с его превращением в печени в пируват и с дальнейшим использованием последнего. Лактат, поступивший из интенсивно работающих мышц, выходит в кровь, а затем поступает в печень. В печени в лактатдегидрогеназной реакции лактат превращается в пируват, который включается в ГНГ, а образовавшаяся глюкоза поступает в кровь и поглощается скелетными мышцами. Эту последовательность событий называют «глюкозо-лактатным циклом», или «циклом Кори». Цикл Кори выполняет две важнейшие функции: обеспечивает утилизацию лактата и предотвращает лактатацидоз. Часть пирувата, образовавшегося из лактата (25%), окисляется печенью до СО2 и Н2О с освобождением энергии.
Пентозофосфатный путь превращения глюкозы
Функционирует в цитозоле клетки с пластической целью, то есть его главные продукты – рибозо-5-фосфат и НАДФН используются для синтезов. Пентозофосфатный путь (ПФП) может протекать в двух вариантах – окислительного и неокислительного.
Окислительный вариант включает 2 реакции дегидрирования, катализируемых НАДФ-зависимыми дегидрогеназами.
Суммарное уравнение окислительного варианта можно представить в таком виде:
Глюкозо-6-фосфат
+ 2 НАДФ+ Рибозо-5-фосфат + 2 НАДФН+ 2Н+
+СО2
Рибозо-5-фосфат + 2 НАДФН+ 2Н+
+СО2
Неокислительный вариант ПФП включает серию обратимых реакций, катализируемых транскетолазой и трансальдолазой, первая в качестве кофермента использует ТПФ. В качестве промежуточных соединений в этом варианте образуются углеводы, содержащие от 3 до 7 атомов углерода. В этом варианте нет реакций дегидрирования, поэтому он используется только для синтеза пентоз.
Суммарное уравнение реакций неокислительного варианта:
5 Глюкозо-6-фосфат 6 рибозо-5-фосфат
Роль ПФП определяется его конечными продуктами: рибозо-5фосфатом и НАДФН. Рибозо-5-фосфат необходим для синтеза нуклеотидов, а затем и нуклеиновых кислот (ДНК и РНК). Быстрый синтез ДНК происходит в быстро делящихся тканях: красный костный мозг, эмбрион, базальный слой эпителия кожи, слизистая тонкого кишечника и др. Также рибозо-5-фосфат необходим для синтеза РНК и далее биосинтеза белка – в печени, экзокринных железах, синтезирующих белок на экспорт, а также усиленный синтез белка происходит при гипертрофии тканей. При этом у взрослого человека есть ткани, которые практически не делятся, а значит, в них ПФП неактивен. Но и в этих тканях ПФП может активироваться, например, при процессах репарации. Другой продукт ПФП – НАДФН необходим для реакций гидроксилирования и для восстановительных биосинтезов. Он активно используется для биосинтеза жирных кислот в жировой ткани, в печени; для синтеза холестерина (печень), для синтеза стероидных гормонов в эндокринных железах, для реакций гидроксилирования эндогенных веществ и ксенобиотиков. В эритроцитах НАДФН необходим для работы антиоксидативной системы, что обеспечивает их устойчивость к гемолизу.
.Гомеостаз глюкозы крови
Уровень глюкозы крови поддерживается на постоянном уровне за счет того, что пути поступления глюкозы в кровь и пути расходования глюкозы крови между собой сбалансированы. При этом в течение суток уровень глюкозы не является одинаковым, но у здорового человека эти колебания лежат в пределах нормы (3,3 – 5,5 ммоль/л). Источниками глюкозы крови являются: углеводы пищи, гликоген печени и органические кислоты, которые в результате глюконеогенеза превращаются в глюкозу. Расходуется глюкоза крови всеми клетками организма, при этом в одни клетки глюкоза поступает диффузно (например, головной мозг), а в другие – с помощью инсулина. Кроме того, глюкоза из крови в результате фильтрации попадает в первичную мочу, но затем в результате реабсорбции она возвращается в сосудистое русло (рис. 6).
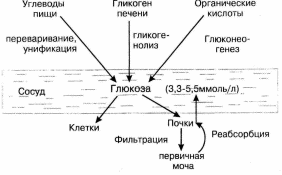
Рис.6. Гомеостаз глюкозы крови
В регуляции уровня глюкозы крови принимают участие гормоны. Гормоны делятся на гипогликемические, т.е. снижающие уровень глюкозы крови, и гипергликемические, т.е. повышающие уровень глюкозы крови. Единственным гипогликемическим гормоном
является инсулин. Гипергликемическими гормонами являются: глюкагон, катехоламины (адреналин и норадреналин), ГКС и соматотропный гормон (СТГ).
При гипергликемии (повышении глюкозы крови, которое может быть вызвано алиментарной причиной, сахарным диабетом, стрессом, некрозом поджелудочной железы, избытком гипергликемических гормонов) усиливается выработка инсулина, который за счет своих эффектов снижает уровень глюкозы крови. Это происходит за счет усиления потребления глюкозы тканями (в связи с увеличением проницаемости мембран для глюкозы в мышечной, жировой и соединительной тканях, усилением синтеза гликогена, гликолиза и пентозофосфатного пути), а также за счет торможения поступления глюкозы из тканей в кровь (в результате торможения распада гликогена и торможения глюконеогенеза).
При гипогликемии (снижении уровня глюкозы крови, которое может быть вызвано алиментарной причиной, патологией печени и ЖКТ, избытком инсулина, при алкоголизме и др.) усиливается выработка гормонов гипергликемических, которые повышают уровень глюкозы крови. Это обусловлено усилением глюконеогенеза и распада гликогена, торможением синтеза гликогена и гликолиза. СТГ при этом действует опосредованно, путем усиления секреции глюкагона.