

Полезные материалы за все 6 курсов / Учебники, методички, pdf / Биохимия в рисунках
.pdfII. Механизмы взаимодействия гормонов с клеткой |
|
Все гормоны делятся на две большие группы в зависимости от |
клеточной |
локализации рецептора (на клеточной мембране или внутри клетки), с которым взаимодействует гормон. Это в свою очередь зависит от структуры и свойств самого гормона:
гидрофильные гормоны – белково-пептидной природы и производные аминокислот
– не способны проникать внутрь клетки-мишени через липофильную клеточную мембрану, действуют на клетку за счет связывания с мембранным рецептором;
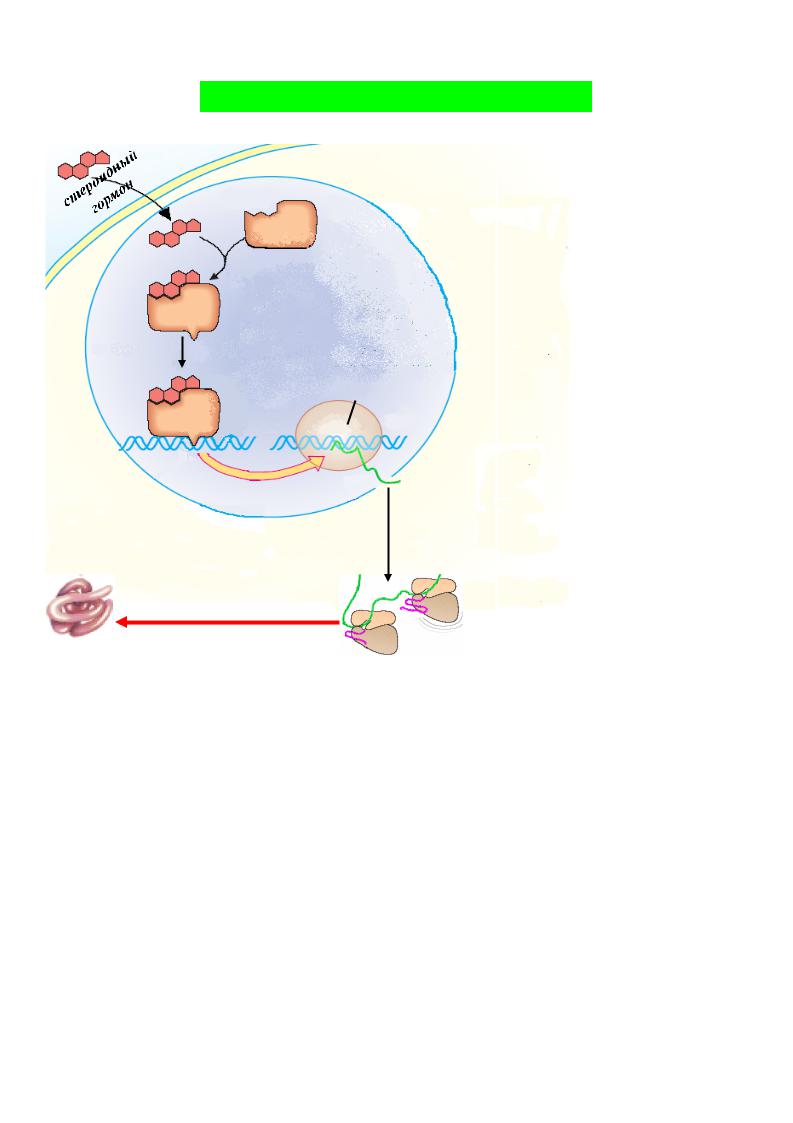
липофильные гормоны – стероидные гормоны и иодтиронины – проникают в клетку-мишень и взаимодействуют со специфическими рецепторами в цитоплазме или в клеточном ядре.
|
|
Механизмдействиягидрофильныхгормонов |
|
|
|
Трансформация гормонального сигнала в специфический ответ клетки-мишени |
|||
происходит в результате следующих молекулярных событий: |
рецепторы, |
|||
|
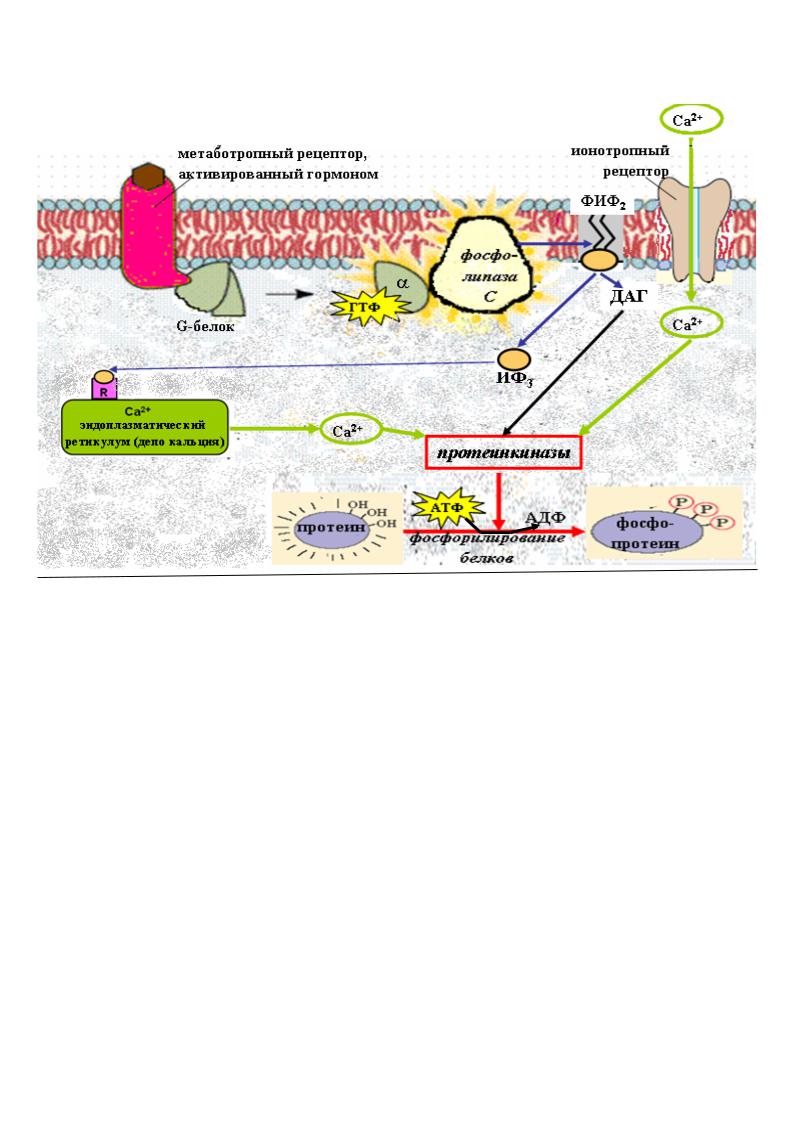
Передача гормонального сигнала осуществляется через белковые |
|||
расположенные на поверхности плазматической мембраны. Различают два основных типа мембранных рецепторов:
ионотропные рецепторы – после взаимодействия с гормоном они способствуют открытию ионных каналов на мембране и развитию быстрых ионных потоков (кальция, натрия, калия, хлора);
метаботропные рецепторы активируют системы клетки через |
белки- |
|
трансдукторы (G-белки). |
|
|
гормон |
эффекторные |
|
метаботропный |
ферменты: |
|
аденилатциклаза, |
||
рецептор |
||
гуанилатциклаза, |
||
|
||
|
фосфолипаза С, |
|
|
фосфолипаза А2. |
G-белки - это внутримембранные белки, которые принимают химический сигнал от рецептора, модифицированного гормоном, и передают этот сигнал на эффекторные системы клетки. Существует несколько типов G-белков, основные из них:
Gs - стимулирующие аденилатциклазу; |
|
Gi - ингибирующие аденилатциклазу; |
|
Gq – активирующие фосфолипазу C. |
150 |

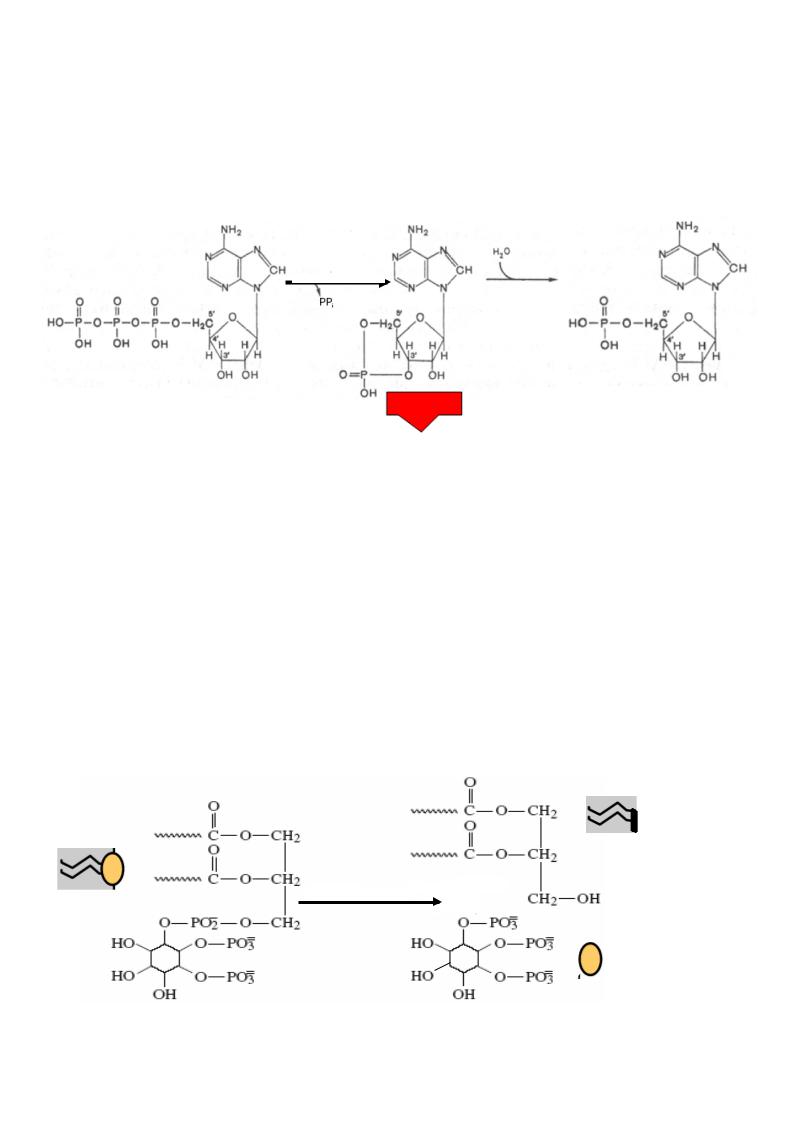
Эффекторные ферменты – аденилатциклаза, гуанилатциклаза, фосфолипаза С и
др. расположены на внутренней поверхности клеточной мембраны. Они стимулируют образование в цитоплазме клетки вторичных посредников – мессенджеров.
Наиболее важными вторичными мессенджерами |
являются: циклический АМФ |
(цАМФ), циклический ГМФ (цГМФ), ионы Са 2+ |
, инозитозтрифосфат ИФ 3,, |
диацилглицерид ДАГ и монооксид азота NO. |
|
Дальнейшая передача гормонального сигнала от вторичных мессенджеров осуществляется с помощью протеинкиназ – ферментов, которые фосфорилируют белки (более 100 различных белков, в том числе ряд ферментов и факторов транскрипции). В результате фосфорилирования изменяется функциональная активность этих белков.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гидрофильные гормоны |
Липофильные гормоны |
|||||||||||
|
|
|
|
|
действуют через |
действуют через |
||||||||||
|
|
|
|
|
мембранные рецепторы |
внутриклеточные рецепторы |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Гормон |
|
|
Гормон |
Гормон |
||||||||
|
(белково-пептидные гормоны, |
|
(инсулин, СТГ) |
(стероидные гормоны, |
||||||||||||
|
|
|
|
адреналин) |
|
Ауто- |
|
|
|
иодтиронины) |
||||||
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
Рецептор ( |
|
внутри клетки) |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
фосфорили |
|
||||||
|
|
Рецептор (в мембране) |
|
|||||||||||||
|
|
|
рование |
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
рецептора |
Комплекс «гормон – рецептор» |
||||||
|
|
|
|
G – белок |
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
фермент |
|
|
G – белок |
|
Транспорт |
|
комплекса |
|||||
|
|
|
Эффекторный |
|
|
|
||||||||||
|
|
|
(аденилатциклаза, |
|
|
|
|
|
|
«гормон - рецептор» в ядро |
||||||
|
|
|
фосфолипаза С, др.) |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
Каскад |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
Взаимодействие комплекса |
|||||||
|
|
|
|
|
|
|
|
|
фосфорилирования |
«гормон - рецептор» с ДНК |
||||||
|
|
|
Вторичный мессенджер |
|
|
|||||||||||
|
|
|
|
|
|
белков |
|
|
|
|||||||
|
|
(цАМФ, Са 2+, ИФ3, ДАГ) |
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Индукция или репрессия |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
транскрипции |
||
|
|
|
|
|
|
|
|
|
|
Активация |
||||||
|
|
|
Протеинкиназы |
|
|
|||||||||||
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
ферментов и |
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
факторов |
|
|
|
|||
|
|
|
|
|
|
|
|
|
Трансляция - |
|||||||
|
|
Фосфорилирование белков |
|
транскрипции |
синтез белков (ферментов) |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Изменение функциональной активности белков |
Изменение количества |
|||||||||||||
|
151 |
|
|
(ферментов) |
|
|
|
|
|
белков (ферментов) |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

III. Гормоны гипоталамо-гипофизарной системы
ГОРМОНЫ ГИПОТАЛАМУСА
Гипоталамус - это зона головного мозга, которая регулирует активность гипофиза и периферических эндокринных желез путем продукции специфических рилизинг-
факторов:
либеринов -способствующих выработке соответствующих тропных гормонов; статинов - ингибирующих секрецию тропных гормонов гипофиза.
Хорошо изучены следующие рилизинг-факторы:
соматолиберин; |
соматистатин; |
тиреолиберин; |
пролактостатин; |
гонадолиберин; |
|
кортиколиберин. |
|
Рилизинг-факторы по химической структуре представляют собой полипептиды, которые секретируются нейронами гипоталамуса и через систему портального кровообращения поступают в аденогипофиз, где стимулируют выработку тропных гормонов.
ГОРМОНЫ ГИПОФИЗА
В передней и средней долях - а деногипофизе вырабатываются тропные гормоны - “гормоны гормонов”. Тропные гормоны реализуют свое действие путем влияния на гормонообразование в перифирических эндокринных железах или непосредственно влияя на обмен веществ в клетках-мишенях. Ряд этих гормонов по структуре являются
гликопротеинами, состоящими из двух белковых субъединиц и углеводного |
|
|||||
компонента. α-Субъединица у всех этих гормонов одинакова, а |
|
β-субъединица |
– |
|||
различна, и именно она определяет биологическую специфичность. |
|
|
||||
Тиреотропный гормон (ТТГ) |
- гликопротеид, состоит из двух |
|
|
|||
субъединиц, влияет на выработку |
щитовидной железой гормонов |
|
||||
иодтронинов (см. с. 49): |
|
|
|
|
|
|
стимулирует рост фолликулярных клеток щитовидной железы; |
|
|
||||
активирует метаболизм иода в щитовидной железе; |
|
|
|
|
||
усиливает включение иода в состав тироксина; |
|
|
ТТГ |
|
||
активирует протеолитические ферменты, освобождающие тироксин |
|
|||||
из тиреоиодглобулина. |
|
|
|
|
|
|
Гонадотропные гормоны |
- сложные белки |
– |
|
|
|
|
гликопротеиды, влияют на гонады (см. с. 59). |
|
|
|
|
||
Фолликулостимулирующиц гормон |
(ФСГ) |
- |
|
|
|
|
стимулирует выработку женских половых гормонов |
|
ФСГ |
ЛГ |
|
||
эстрогенов, а также созревание фолликулов яичников у |
|
|||||
|
|
|
||||
женщин и сперматогенез у мужчин. |
|
|
|
|
|
|
Лютеинизирующий гормон ( |
ЛГ) |
- стимулирует |
|
|
|
|
выработку прогестерона у женщин и тестостерона у |
|
|
|
|
||
мужчин. |
|
|
|
|
|
|
Хорионический гонадотропин (ХГ) – гликопротеин. |
|
ХГ |
156 |
|||

