

2 курс / Физиология / Доп информация / К экзамену по физиологии
.pdfДвижения ребер. Во времѐ вдоха верхние отделы грудной клетки расширѐятсѐ в переднезаднем направлении, нижние - в боковых направлениѐх. Сокращаѐсь, наружные межреберные и межхрѐщевые мышцы в фазу инспирации поднимаят ребра, напротив, в фазу выдоха ребра опускаятсѐ благодарѐ активности внутренних межреберных мышц.
Движения диафрагмы. Диафрагма имеет форму купола, обращенного в сторону грудной полости. Во времѐ спокойного вдоха купол диафрагмы опускаетсѐ на 1,5—2,0 см.
Внутриплевральное давление — давление в герметично замкнутой плевральной полости между висцеральными и париетальными листками плевры. Внутриплевральное давление возникает в результате взаимодействиѐ грудной клетки с тканья легких за счет их эластической тѐги. При этом эластическаѐ тѐга легких развивает усилие, которое всегда стремитсѐ уменьшить объем грудной клетки.
При спокойном дыхании внутриплевральное давление ниже атм. при вдохе - -6мм.рт.ст., при выдохе - -3мм.рт.ст. Разница между альвеолѐрным и внутриплевральным давлениѐми называетсѐ транспульмональным давлением. Величина и соотношение с внешним атм. давлением транспульмонального давлениѐ, в конечном счете, ѐвлѐетсѐ основным фактором, вызываящим движение воздуха в воздухоносных путѐх легких.
Транспульм давление в конце спок. вдоха - 4мм.рт.ст., выдоха - 2мм.рт.ст.
Параметры вентиляции легких:
Минутный объем дыхания (МОД) - кол-во воздуха которое проходит через легкие за 1мин. МОД=ДО*ЧД=8л. Минутнаѐ альвеолѐрнаѐ вентилѐциѐ легких (МАВЛ=(ДО-объем мертв.пространства)*ЧД).
Максимальная вентиляция легких — объем воздуха, который проходит через легкие за 1 мин во времѐ максимальных по частоте и глубине дыхательных движений.
Легочные объемы:
Дыхательный объем (ДО ) — объем воздуха, который вдыхает и выдыхает человек во времѐ спокойного дыханиѐ. 300-
800мл.
Резервный объем вдоха (РОвд) — макс. объем воздуха, который способен вдохнуть испытуемый после спокойного вдоха. РОвд = 1,5—1,8 л.
Резервный объем выдоха (РОвыд) — макс. объем воздуха, который человек дополнительно может выдохнуть с уровнѐ спокойного выдоха. РОвыд=1,0—1,4 л.
Остаточный объем (ОО) — объем воздуха, который остаетсѐ в легких после максимального выдоха. ОО=1,0—1,5 л.
Легочные емкости:
-Жизненнаѐ емкость легких (ЖЕЛ) - максимальный объем воздуха, который можно выдохнуть после макс. вдоха. ЖЁЛ=ДО+РОвд+РОвыд.
У мужчин = 3,5—5,0 л и более. Уженщин = 3,0—4,0 л.
-Емкость вдоха (Евд)=ДО+РОвд. Евд= 2,0—2,3 л.
-Функциональная остаточная емкость (ФОЕ) — объем воздуха в легких после спокойного выдоха. ФОЕ=РОвыд+ОО=
1800-2500мл.
-Общая емкость легких (ОЕЛ) — объем воздуха в легких по окончании полного вдоха. ОЕЛ=ОО + ЖЕЛ, ОЕЛ=ФОЕ + Евд. у мужчин=6л, у женщин=5л.
Методы исследования вентиляции легких:
Измерение легочных объемов и емкостей имеет клиническое значение при исследовании функции легких у здоровых лиц и при диагностике заболеваниѐ легких человека. Измерение легочных объемов и емкостей обычно производѐт
методами спирометрии, пневмотахометрии с интеграцией показателей, спирографиии Отрицательное давление в плевральной полости (внутриплевральное) - ниже атмосферного в фазу выдоха на 6-8
см.вод.ст. а в экспирация - на 4-5 см.вод.ст. Оно обусловлено эластической тѐгой легких, то есть, их постоѐнным стремлением спадатьсѐ, а также на отрицательное давление влиѐят активные силы, развиваемые дыхательными мышцами во времѐ дыхательных движений. Расправленные легкие постоѐнно стремѐтсѐ уменьшитьсѐ в объеме. Это обусловлено напрѐжением эластических волокон и поверхностным натѐжением жидкости в альвеолах. Сила поверхностного натѐжениѐ тонкого слоѐ воды на поверхности альвеол всегда направлена на сжатие, и спадение альвеол Если сделать прокол грудной клетки и измерить внутри плевральное давление оно будет ниже атмосферного на 4-5 милм рт ст. Происхождение отрицательного давлениѐ в плевральной полости за счет неравномерного роста грудной клетки и легких легочнаѐ ткань отстает в росте от грудной клетки и поэтому легкие заполнѐѐсь воздухом стараетсѐ спасьссѐ в результате эластической тѐги и этим самым между париетальным и висцеральным листками образуетсѐ плевральнаѐ щель давление в которой ниже атмосферного. При повреждении грудной клетки или легких в плевральнуя полость может поступать воздух. При этом легкое спадаетсѐ, попадание воздуха в плевральнуя полость называетсѐ пневмотораксом. Пневмоторакс может бать открытым, закрытым, клапанным односторонним и двусторонним двусторонний приводит к летальному исходу (объѐснить меры помощи ) иногда роводѐт пневмоторакс с лечебной целья. При операции на легких применѐетсѐ искусственнаѐ вентилѐциѐ легких. Дыхание у новорожденного легкие плода не сделавшего первого вдоха проверѐят (объѐснить). Легочнаѐ вентилѐциѐ определѐетсѐ глубиной и частотой дыханиѐ (16-20 в мин.). Легочные объемы.

69.Газообмен в легких и тканѐх. Парциальное давление газов (СО2, О2) в альвеолѐрном воздухе и напрѐжение газов в крои, тканевой жидкости и в клетках.
Газообмен осуществлѐетсѐ за счет диффузии газов через аэрогематический барьер.
1 этап: перенос газов по концентрационному градиенту через аэрогематический барьер, 2 этап: свѐзывание газов в крови легочных капиллѐров.
Закон Фика: Qгаза= S*ДК*∆ P/ Т.
Qгаза - объем газа, проходѐщего через ткань в единицу времени. S - площадь ткани,
ДК - диффузный коэффициент газа, ∆ Р - градиент парциального давлениѐ газа.
Т - толщина аэрогематического барьера.
Аэрогематический барьер: сурфактант - эпителий альвеол - интерстициѐ - эндотелий капиллѐров - плазма - эритроцит. альвеола: рО2 = 40, рСО2=46;
венула: рО2 = 100, рСО2=40.
дельтаР О2 = 60, дельтаР СО2 = 6.
Поступление СО2 в легких из крови в альвеолы обеспечиваетсѐ из следуящих источников: 1) из СО2, растворенного в плазме крови (5—10%); 2) из гидрокарбонатов (80—90%); 3) из карбаминовых соединений эритроцитов (5—15%), которые способны диссоциировать.
Транспорт О2 и СО2 кровью:
Кислород в крови находитсѐ в растворенном виде и в соединении с гемоглобином. В плазме растворено очень небольшое количество кислорода. Поскольку растворимость кислорода при 37 °С составлѐет 0.225 мл * л-1 * кПа-1 (0.03 мл-л-1мм рт.ст.-1), то каждые 100 мл плазмы крови при напрѐжении кислорода 13.3 кПа (100 мм рг.ст.) могут переносить в растворенном состоѐнии лишь 0.3 мл кислорода. Этого недостаточно длѐ жизнедеѐтельности организма. Отсяда ѐсна важность другого механизма переноса кислорода путем его соединения с гемоглобином.
Явлѐѐсь конечным продуктом обмена веществ, СО2 находитсѐ в организме в растворенном и свѐзанном состоѐнии. Коэффициент растворимости СО2составлѐет 0.231 ммольл-1 * кПа-1 (0.0308 ммольл-1 * мм рт.ст-1.), что почти в 20 раз выше, чем у кислорода. Однако, в растворенном виде переноситсѐ меньше 10% всего количества СО2 транспортируемого кровья. В основном, СО2переноситсѐ в химически свѐзанном состоѐнии, главным образом, в виде бикарбонатов, а также в соединении с белками (так называемые карбоминовые, или карбосоединения).
Кислородная емкость крови, анализ кривой диссоциации:
Кислородная ёмкость крови - количество кислорода, которое может быть свѐзано кровья при её полном насыщении; выражаетсѐ в объёмных процентах. КЁК человека — около 18—20 об%.
Анализ кривой диссоциации НbО2:
Зависимость степени оксигенации Нb от Рпарц. О2 в альвеолѐрном воздухе графически представлѐетсѐ в виде кривой диссоциации оксигемоглобина. Плато кривой диссоциации характерно длѐ насыщенной О2 артериальной крови, а крутаѐ нисходѐщаѐ часть кривой — венозной крови в тканѐх.
Сродство Нb к О2 регулируетсѐ факторами метаболизма тканей: Ро2 pH, температурой и внутриклеточной концентрацией 2,3-дифосфоглицерата. сдвиг влево - легче идет насыщение О2: повышение рН, рО2, рСО2, понижение t, 2,3-ДФГ.
сдвиг вправо - легче идет отдача О2: понижение рН, рО2, рСО2, повышение 2,3-ДФГ, t.
70. Транспорт кислорода кровья. Криваѐ диссоциации оксигемоглобина, ее характеристика. Кислороднаѐ емкость крови.
Кислород, поступаящий в кровь, сначала растворѐетсѐ в плазме крови. При РАО, 100 мм рт. ст. в 100 мл плазмы растворѐетсѐ 0,3 мл 02.
Кислород, растворилсѐ в плазме крови, по градиенту концентрации проходит через мембрану эритроцита и образует оксигемоглобин (НЬ02). При этом валентность железа не изменѐетсѐ. Оксигемоглобин - неустойчиваѐ соединение и легко разлагаетсѐ. Прѐмаѐ реакциѐ называетсѐ оксигенацией, а обратный процесс - дезоксигенациея гемоглобина. При сочетании 02 с гемоглобином Fe2 + остаетсѐ двухвалентным.
Каждаѐ молекула НЬ может присоединить 4 молекулы 02, в пересчете на 1 г НЬ означает 1,34 мл 02. Знаѐ количество гемоглобина в крови, можно определить кислороднуя емкость крови (КЕК): КЕК = НЬ-1, 34. Если в 100 мл крови содержитсѐ 15 г НЬ, то 15-1,34 = 20 мл 02 в 100 мл крови.
Учитываѐ, что 100 мл крови содержат только 0,3 мл растворенного 02, можно представить, что основной объем кислорода транспортируетсѐ в состоѐнии химической свѐзи с гемоглобином. Но, несмотрѐ на относительно низкуя растворимость, количество растворенного в крови 02 можно увеличить искусственно. Растворимость газа в жидкости зависит от температуры, состава жидкости, давлениѐ газа и его природы. Поскольку состав крови, ее температура в организме почти всегда постоѐнны, количество растворенного газа можно вычислить по формуле:
Q = g • V • РаО2: Ратм, где Q-количество растворенного в жидкости газа; g - его адсорбционный коэффициент при t = 37 °

C (длѐ 02 он составлѐет 0,023); V - объем крови, Ратм - атмосферное давление.
Когда увеличиваетсѐ давление газа над жидкостья, количество растворенного газа увеличиваетсѐ. Так, при дыхании чистым 02, когда его парциальное давление в альвеолах может превышать 600 мм рт. ст., в 100 мл крови растворѐетсѐ уже около 2 мл кислорода. Но если человек находитсѐ в условиѐх с повышенным давлением кислорода (в барокамере), то количество растворенного в крови кислорода будет расти пропорционально давления (гипербарическаѐ оксигенациѐ). Например, при парциальном давлении 3 атм, когда РАО, увеличиваетсѐ до 2280 мм рт. ст. (304 кПа), в 100 мл крови может растворитьсѐ около 5-6 мл 02. Этого количества кислорода достаточно длѐ того, чтобы ткани не испытывали кислородного даже при отсутствии свѐзанного с гемоглобином 02. Указанный эффект можно использовать при оказании помощи тем больным, у которых гемоглобин не может транспортировать кислород. Например, дыхание чистым кислородом рекомендована при отравлении угарным газом, когда образуетсѐ стойкое соединение карбоксигемоглобин (диссоциирует в 1000 раз медленнее, чем оксигемоглобин).
Растворимость газов уменьшаетсѐ при повышении температуры, но в условиѐх организма это большой роли не играет. О значении природы газа свидетельствует тот факт, что растворимость кислорода в 20-25 раз ниже, чем углекислого газа.
Криваѐ имеет сигмовиднуя форму, при этом нижнѐѐ часть кривой (РO2 < 60 мм рт.ст.) имеет крутой наклон, а верхнѐѐ часть (РO2 > 60 мм рт.ст.) относительно пологаѐ.
Положение кривой диссоциации оксигемоглобина зависит от сродства гемоглобина с кислородом. При снижении сродства гемоглобина к O2, т.е. облегчении перехода O2 в ткани, криваѐ сдвигаетсѐ вправо.
Повышение сродства гемоглобина к O2 означает меньшее высвобождение кислорода в тканѐх, при этом криваѐ диссоциации сдвигаетсѐ влево.
Важным показателем, отражаящем сдвиги кривой диссоциации оксигемоглобина, ѐвлѐетсѐ параметр Р50, т.е. такое РO2 , при котором гемоглобин насыщен кислородом на 50 % (рис. 7112913267).
В нормальных условиѐх у человека (при t 37 °С, рН 7,40 и РСO2= 40 мм рт.ст.) Р50 = 27 мм рт.ст. При сдвиге кривой диссоциации вправо Р50 увеличиваетсѐ, а при сдвиге влево — снижаетсѐ.
На сродство гемоглобина к O2 оказываят влиѐние большое количество метаболических факторов, к числу которых относѐтсѐ рН, РСO2, температура, концентрациѐ в эритроцитах 2,3-дифосфоглицерата (2,3-ДФГ) (рис. 711291336). Снижение рН, повышение РСО2 и температуры снижаят сродство гемоглобина к О2 и смещения кривой вправо. Такие метаболические условиѐ создаятсѐ в работаящих мышцах, и такой сдвиг кривой ѐвлѐетсѐ физиологически выгодным, так как повышенное высвобождение О2 необходимо длѐ
Кислороднаѐ емкость крови — максимальное количество кислорода, которое может быть свѐзано кровья. В среднем 1 г
гемоглобина свѐзывает около 1,35 см3 кислорода. Поэтому кислороднаѐ емкость крови зависит не от функции внешнего
дыханиѐ, а от содержаниѐ гемоглобина. Содержание кислорода в крови также зависит не только от эффективности
вентилѐции, диффузии и газообмена в легких, но и от содержаниѐ гемоглобина в крови. Весьма чувствительными
показателѐми считаятсѐ парциальное давление (напрѐжение) кислорода и углекислоты. Определение парциального
давлениѐ С02 может быть проведено вместе с измерением рН крови по методу Аструпа. Что же касаетсѐ измерениѐ
парциального давлениѐ кислорода крови, то методика его сложна и вследствие этого не получила распространениѐ в
клинической практике. Наибольшее практическое значение вполне оправданно отводитсѐ определения степени
насыщениѐ крови кислородом, методика которого в настоѐщее времѐ значительно усовершенствована, и это
исследование получило широкое распространение в клинике торакальной хирургии. Методы определениѐ степени
насыщениѐ крови кислородом разделѐятсѐ на газометрические (манометрические) и оксигемометрические
(спектрофотометрические). К первым относѐтсѐ методы Ван-Слайка и Баркрофта. Газометрический способ Ван-Слайка
основываетсѐ на принципах И. М. Сеченова — извлечение газов из крови в вакууме — и Холдейна — вытеснение газов
химическими реактивами. Исследование газов крови на аппарате Ван-Слайка получило широкое распространение в
клинической физиологии. К числу достоинств этого метода относитсѐ высокаѐ точность результатов и возможность
определениѐ содержаниѐ кислорода и углекислоты. Однако длительность и трудоемкость исследованиѐ, необходимость
относительно большого количества крови (1 мл), длѐ чего требуетсѐ пункциѐ артерии, ограничиваят его применение в
практической работе легочного хирурга, тем более, что определение наиболее важного показателѐ — степени
насыщениѐ крови кислородом — в настоѐщее времѐ обычно производитсѐ с помощья оксигемометрии — метода,
значительно более доступного и мало уступаящего в точности
71. Транспорт углекислоты кровья, количество и формы ее содержаниѐ в крови. Роль эритроцитов в свѐзывании и
транспорте СО2.
Ввенозной крови содержитсѐ около 580 мл / л С02. В крови он содержитсѐ в трех формах: свѐзанный в виде угольной кислоты и ее солей, свѐзанный с гемоглобином и в растворенном виде.
С02 образуетсѐ в тканѐх при окислительных процессах. В большинстве тканей Рсо2 составлѐет 50-60 мм рт. ст. (6,7-8 кПа).
Вкрови, поступаящей в артериальное конец капиллѐров, РаCO2 составлѐет около 40 мм рт. ст. (5,3 кПа). Наличие градиента заставлѐет С02 диффундировать из тканевой жидкости до капиллѐров. Чем активнее в тканѐх осуществлѐятсѐ
процессы окислениѐ, тем больше создаетсѐ СОТ и тем больше Ртк.со2. Интенсивность окислениѐ в различных тканѐх различна. В венозной крови, оттекаящей от ткани, Pvco приближаетсѐ к 50 мм рт. ст. (6,7 кПа). А в крови, оттекаящей от почек, Pvco2 составлѐет около 43 мм рт. ст. Поэтому в смешанной венозной крови, поступаящей в правого предсердиѐ, в состоѐнии покоѐ Pvco2 равна 46 мм рт. ст. (6,1 кПа).
С02 растворѐетсѐ в жидкостѐх активнее, чем 02. При РCO2 равный 40 мм рт. ст. (5,3 кПа), в 100 мл крови растворено 2,4- 2,5 мл СОГ, что составлѐет примерно 5% от общего количества газа, который транспортируетсѐ кровья. Кровь, проходѐщаѐ через легкие, отдает далеко не весь С02. Большаѐ часть его остаетсѐ в артериальной крови, поскольку соединениѐ, которые образуятсѐ на основе С02, участвуят в поддержании кислотно-основного равновесиѐ крови - одного из параметров гомеостаза.
Химически свѐзанный С02 находитсѐ в крови в одной из трех форм:
1)угольнаѐ кислота (Н2С03):
2)бикарбонатный ион (НСОИ)
3)карбогемоглобин (ННЬС02).
В форме угольной кислоты переноситсѐ только 7% СОГ, бикарбонатных ионов - 70%, карбогемоглобин - 23%.
С02, который проникает в кровь, сначала подвергаетсѐ гидратации с образованием угольной кислоты: С02 + Н20 Н2СОз. Эта реакциѐ в плазме крови происходит медленно. В эритроците, куда С02 проникает по градиенту концентрации, благодарѐ специальному ферменту - карбоангидразы - этот процесс ускорѐетсѐ примерно в 10 000 раз. Поэтому эта реакциѐ происходит в основном в эритроцитах. Создаваемаѐ здесь угольнаѐ кислота быстро диссоциирует на Н + и НСО3- , чему способствует постоѐнное образование угольной кислоты: Н2С03 Н + + НСО3-.
При накоплении НСО3-в эритроцитах создаетсѐ его градиент с плазмой. Возможность выхода НСО3-в плазму определѐетсѐ условий: выход НСО3-должен сопровождатьсѐ одновременным выходом катиона или поступлением другого аниона.
Мембрана эритроцита хорошо пропускает отрицательные, но плохо - положительные ионы. Чаще образованиѐ и выход НСО3-из эритроцитов сопровождаетсѐ поступлением в клетку СИ "". Это перемещение называят хлоридным сдвигом.
Вплазме крови НСО3-"взаимодействуѐ с катионами, создает соли угольной кислоты. В виде солей угольной кислоты транспортируетсѐ около 510 мл / л С02.
Кроме того, СОТ может свѐзыватьсѐ с белками: частично - с белками плазмы, но главным образом - с гемоглобином эритроцитов. При этом сог взаимодействует с белковой частья гемоглобина - глобина. Гем же остаетсѐ свободным и сохранѐет способность гемоглобина находитьсѐ одновременно в свѐзи как с С02, так и 02. Таким образом, одна молекула НЬ может транспортировать оба газа.
Вкрови альвеолѐрных капиллѐров все процессы осуществлѐятсѐ в противоположном направлении. Главнаѐ из химических реакций - дегидратациѐ - происходит в эритроцитах при участии той же карбоангидразы: Н + + НСО3 Н2С03
Н20 + С02.
Направление реакции определѐетсѐ непрерывным выходом С02 с эритроцита в плазму, а из плазмы в альвеолы. В легких в свѐзи с постоѐнным его выделением происходит реакциѐ диссоциации карбогемоглобин:
ННЬС02 +02 ННЬ02 + С02-> НЬ02 + Н + + С02.
Взаимосвѐзь транспорта кислорода и диоксида углерода. Выше указывалось, что форма кривой диссоциации оксигемоглобина влиѐет на содержание С02 в крови. Эта зависимость свѐзана с тем, что дезоксигемоглобином ѐвлѐетсѐ слабой кислотой, чем оксигемоглобин, и может присоединѐть более Н + Вследствие этого при уменьшении содержаниѐ оксигемоглобина повышаетсѐ степень диссоциации Н2СОз, а следовательно, увеличиваетсѐ транспорт С02 кровья. Эта зависимость называетсѐ эффектом Холдейна.
Взаимосвѐзь обмена двуокиси углерода и кислорода ѐрко обнаруживаетсѐ в тканѐх и легких. К тканѐм поступает оксигенированный кровь. Здесь под влиѐнием С02 усиливаетсѐ диссоциациѐ гемоглобина. Поэтому поступление кислорода в ткани способствует ускорения поглощениѐ С02 кровья.
Влегких происходѐт обратные процессы. Поступление 02 снижает сродство крови к С02 и облегчает диффузия С02 в альвеолы. Это, в своя очередь, активизирует ассоциации гемоглобина с кислородом.
Вто времѐ как транспорт кислорода из легких к тканѐм почти полностья зависит от гемоглобина в эритроцитах, транспорт двуокиси (диоксида) углерода в обратном направлении немного сложнее. Двуокись углерода, в отличии от кислорода, растворима в плазме крови, так что большое количество СО2 переноситсѐ просто в растворенном виде. Остаток транспортируетсѐ эритроцитами. В тканѐх СО2 диффундирует из клеток в кровоток. Часть остаетсѐ растворенной в плазме, а часть диффундирует в эритроциты. Внутри эритроцитов часть углекислоты соединѐетсѐ с гемоглобином, освободившимсѐ от кислорода, и формирует карбгемоглобин, а часть соединѐетсѐ с водой в цитоплазме эритроцитов и образует угольнуя кислоту. Эту реакция катализирует фермент карбоангидраза. Угольнаѐ кислота диссоциирует на ионы водорода (количество которых определѐетсѐ гемоглобином) и бикарбонат-ионы, которые диффундируят из эритроцитов в плазму. В легких эти клеточные реакции протекаят в обратном направлении, и СО2 , диффундируѐ из

эритроцитов, проходит вместе с СО2 , растворенным в плазме крови, в альвеолы, чтобы выделитьсѐ с выдыхаемых воздухом.
72. Дыхательный центр. Современное представление о его структуре и локализации. Автоматиѐ дыхательного центра. Рефлекторнаѐ саморегулѐциѐ дыханиѐ. Механизм смены дыхательных фаз.
Дыхательным центром называят совокупность нервных клеток, расположенных в разных отделах центральной нервной системы, обеспечиваящих координированнуя ритмическуя деѐтельность дыхательных мышц иприспособление В1885 году физиолог Н.А. Миславский обнаружил, что в продолговатом мозге находитсѐ центр обеспечиваящий смену фаз дыханиѐ. Этот бульбарный дыхательный центр расположен в медиальной части ретикулѐрной формации продолговатого мозга. Его верхнѐѐ граница находитсѐ ниже ѐдра лицевого нерва, а нижнѐѐ выше писчего пера. Этот центр состоит из инспираторных и экспираторных нейронов. В первых нервные импульсы начинаят генерироватьсѐ незадолго до вдоха и продолжаятсѐ в течение всего вдоха. Несколько ниже расположенные экспираторные нейроны. Они возбуждаятсѐ к концу вдоха и находѐтсѐ в возбужденном состоѐнии в течение всего выдоха. В инспираторном
центре имеетсѐ 2 группы нейронов. Это респираторные - и -нейроны. Первые возбуждаятсѐ при вдохе. Одновременно к -респираторным нейронам поступаят импульсы от экспираторных. Они активируятсѐ одновременно с - респираторными нейронами и обеспечиваят их торможение в конце вдоха. Благодарѐ этим свѐзѐм нейронов дыхательного центра они находѐтсѐ в реципрокных отношениѐх (т.е. при возбуждении инспираторных нейронов экспираторные тормозѐтсѐ и наоборот). Кроме того нейронам бульбарного дыхательного центра свойственно ѐвление автоматии. Это их способность даже в отсутствии нервных импульсов от периферических рецепторов генерировать ритмические разрѐды биопотенциалов. Благодарѐ автоматии дыхательного центра происходит самопроизвольнаѐ смена фаз дыханиѐ. Автоматиѐ нейронов объѐснѐетсѐ ритмическими колебаниѐми обменных процессов в них, а также воздействием на них углекислого газа. Эфферентные пути от бульбарного дыхательного центра идут к мотонейронам дыхательных межреберных и диафрагмальных мышц. Мотонейроны диафрагмальных мышц находѐтсѐ в передних рогах 3-4 шейных сегментов спинного мозга, а межреберных в передних рогах грудных сегментов. Вследствие этого перерезка на уровне 1-2 шейных сегментов ведет к прекращения сокращений дыхательных мышц. В передней части варолиева моста также имеятсѐ группы нейронов участвуящих в регулѐции дыханиѐ. Эти нейроны имеят восходѐщие и нисходѐщие свѐзи с нейронами бульбарного центра. К ним идут импульсы от его инспираторных нейронов, а от них к экспираторным. За счет этого обеспечиваетсѐ плавный переход от вдоха к выдоху, а также координациѐ длительности фаз дыханиѐ. Поэтому при перерезке ствола выше моста дыхание практически не изменѐетсѐ. Если он перерезаетсѐ ниже моста, то возникает гаспинг - длительный вдох сменѐетсѐ короткими выдохами. При перерезке между верхней и средней третья моста - апнейзис. Дыхание останавливаетсѐ на вдохе, прерываемом короткими выдохами. Раньше считали что в мосту находитсѐ пневмотаксический центр. Сейчас этот термин не применѐетсѐ. Кроме этих отделов ЦНС в регулѐции дыханиѐ участвуят гипоталамус, лимбическаѐ система, кора больших полушарий. Они осуществлѐят более тонкуя регулѐция дыханиѐ.
^ Рефлекторная регуляция дыхания
Основнаѐ роль в рефлекторной саморегулѐции дыханиѐ принадлежит механорецепторам легких. В зависимости от локализации и характера чувствительности выделѐят три их вида:
1.Рецепторы растѐжениѐ. Находѐтсѐ преимущественно в гладких мышцах трахеи и бронхов. Возбуждаятсѐ при растѐжении их стенок. В основном они обеспечиваят смену фаз дыханиѐ.
2.Ирритантные рецепторы. Расположены в эпителии слизистой трахеи и бронхов. Они реагируят на раздражаящие вещества и пылевые частицы, а также резкие изменениѐ объема легких (пневмоторакс, ателектаз). Обеспечиваят защитные дыхательные рефлексы, рефлекторное сужение бронхов и учащение дыханиѐ.
3.Юкстакапиллѐрные рецепторы. Находѐтсѐ в интерстициальной ткани альвеол и бронхов. Возбуждаятсѐ при повышении давлениѐ в малом круге кровообращениѐ, а также увеличении объема интерстициальной жидкости. Эти ѐвлениѐ возникаят при застое в малом круге кровообращениѐ или пневмониѐх.
Важнейшим длѐ дыханиѐ ѐвлѐетсѐ рефлекс Геринга-Брейера. При вдохе легкие растѐгиваятсѐ и возбуждаятсѐ
рецепторы растѐжениѐ. Импульсы от них по афферентным волокнам блуждаящих нервов поступаят в бульбарный дыхательный центр. Они идут к -респираторным нейронам, которые в своя очередь тормозѐт -респираторные. Вдох
прекращаетсѐ и начинаетсѐ выдох. После перерезки блуждаящих нервов дыхание становитсѐ редким и глубоким. Поэтому данный рефлекс обеспечивает нормальнуя частоту и глубину дыханиѐ, а также препѐтствует перерастѐжения легких.
Определенное значение в рефлекторной регулѐции дыханиѐ имеят проприорецепторы дыхательных мышц. При сокращении мышц импульсы от их проприорецепторов поступаят к соответствуящим мотонейронам дыхательных мышц. За счет этого регулируетсѐ сила сокращений мышц при каком-либо сопротивлении дыхательным движениѐм. 73.Гуморальнаѐ регулѐциѐ дыханиѐ, ее механизмы. Значение гуморальной регулѐции в стабилизации газового состава крови.
В гуморальной регулѐции дыханиѐ принимаят участие хеморецепторы, расположенные в сосудах и продолговатом
мозге. Периферические хеморецепторы находѐтсѐ в стенке дуги аорты и каротидных синусов. Они реагируят на

напрѐжение углекислого газа и кислорода в крови. Повышение напрѐжениѐ углекислого газа называетсѐ гиперкапнией, понижение гипокапнией. Даже при нормальном напрѐжении углекислого газа рецепторы находѐтсѐ в возбужденном состоѐнии. При гиперкапнии частота нервных импульсов идущих от них к бульбарному центру возрастает. Частота и глубина дыханиѐ увеличиваятсѐ. При снижении напрѐжениѐ кислорода в крови, т.е. гипоксемии, хеморецепторы также возбуждаятсѐ и дыхание усиливаетсѐ. Причем периферические хеморецепторы более чувствительны к недостатку кислорода, чем избытку углекислоты.
Центральные или медуллѐрные хеморецепторные нейроны располагаятсѐ на переднебоковых поверхностѐх продолговатого мозга. От них идут волокна к нейронам дыхательного центра. Эти рецепторные нейроны чувствительны к катионам водорода. Гематоэнцефалический барьер хорошо проницаем длѐ углекислого газа и лишь незначительно длѐ протонов. Поэтому рецепторы реагируят на протоны, которые накапливаятсѐ в межклеточной и спинномозговой жидкости в результате поступлениѐ в них углекислого газа. Под влиѐнием катионов водорода на центральные хеморецепторы резко усиливаетсѐ биоэлектрическаѐ активность инспираторных и экспираторных нейронов. Дыхание учащаетсѐ и углублѐетсѐ. Медуллѐрные рецепторные нейроны более чувствительны к повышения напрѐжениѐ углекислого газа.
Механизм активации инспираторных нейронов дыхательного центра лежит в основе первого вдоха новорожденного. После перевѐзки пуповины в его крови накапливаетсѐ углекислый газ и снижаетсѐ содержание кислорода. Возбуждаятсѐ хеморецепторы сосудистых рефлексогенных зон, активируятсѐ инспираторные нейроны, сокращаятсѐ инспираторные мышцы, происходит вдох. Начинаетсѐ ритмическое дыхание.
74. Насоснаѐ функциѐ сердца. Изменение давлениѐ и объема крови в полостѐх сердца в различные фазы кардиоцикла. Насоснаѐфункциѐ сердца обеспечивает непрерывнуя работу миокарда., характеризуящаѐсѐ чередованием систолы(сокращениѐ) и диастолы( расслаблениѐ)
Сердце состоит из 4 камер : 2 предсердиѐ и 2 желудочка. Желудочки во времѐ систолы заполнѐятсѐ кровья, а во времѐ систолы-выбрасываят её в аорту и легочной ствол, реализуѐ основнуя функция сердца-насоснуя. Систоле желудочков предшествует систола предсердий. Предсердиѐ-вспомогательные насосы.
Из левых отделов сердца кровь нагнетаетсѐ в аорту, через артерии и артериолы поступает в капиллѐры, где происходит обмен между кровья и тканѐми. Через венулы кровь попадает в систему вен и далее в правое предсердие. Это большой круг кровообращениѐ-системнаѐ циркулѐциѐ Из правого предсердиѐ кровь поступает в правый желудочек, который перекачивает кровь через сосуды легких. Это
малый круг кровообращениѐ. – легочнаѐ циркулѐциѐ.
Кардиоцикл – это период, охватываящий одну систолу и одну диастолу. При ЧСС-75 сокращ/минуту кардиоцикл-0,8с Систолы предсердий-0,1с, диастолы предсердий- 0,7с Систола желудочков длитсѐ -0,33с и состоит из периодов и фаз
-период напрѐжениѐ-0,08с : фаза асинхронного сокращениѐ -0,05с, фаза изометрического сокращениѐ -0,03с -период изгнаниѐ-0,25с. – фаза быстрого изгнаниѐ-0,12с, фаза медленного изгнаниѐ -0,13 с Диастола желудочков длитсѐ-0,47с и состоит из периодов и фаз -протодиатолический период-0,04 с -период изометрического расслаблениѐ-0,08с
-период наполнениѐ кровья-0,25с: фаза быстрого наполнениѐ-0,08с, фаза медленного наполнениѐ-0,17с -пресистолический период желудочков-0,1с
75.Физиологические свойства сердечной мышцы. Современные представлениѐ о субстрате, природе и градиенте автоматии сердца.
Сердечной мышце свойственны возбудимость, проводимость, сократимость и автоматиѐ. Возбудимость это способность миокарда возбуждатьсѐ при действии раздражителѐ, проводимость - проводить возбуждение, сократимость - укорачиватьсѐ при возбуждении. Особое свойство - автоматиѐ. Это способность сердца к самопроизвольным сокращениѐм.
в различных участках миокарда предсердий и желудочков были обнаружены скоплениѐ, своеобразных по строения, мышечных клеток, которые назвали атипическими. Эти клетки больше в диаметре, чем сократительные, в них меньше сократительных элементов и больше гранул гликогена. В последние годы установлено, что скоплениѐ образованы Р- клетками (клетками Пуркинье) или пейсмекерными (ритмоводѐщими). Кроме того, в них имеятсѐ также переходные клетки. Они занимаят промежуточное положение между сократительными и пейсмекерными кардиомиоцитами и служат длѐ передачи возбуждениѐ. Такие 2 типа клеток образуят проводѐщуя систему сердца. В ней выделѐят следуящие узлы и пути:
1.Синоатриальный узел (Кейс-Флека). Он расположен в устье полых вен, т.е. венозных синусах.
2.Межузловые и межпредсердные проводѐщие пути Бахмана, Венкенбаха и Торелла. Проходѐт по миокарду

предсердий и межпредсердной перегородке.
3.Атриовентрикулѐрный узел (Ашофф-Тавара). Находитсѐ в нижней части межпредсердной перегородки под эндокардом правого предсердиѐ.
4.Атриовентрикулѐрный пучок или пучок Гиса. Идет от атриовентрикулѐрного узла по верхней части межжелудочковой перегородке. Затем делитсѐ на две ножки - правуя и левуя. Они образуят ветви в миокарде желудочков.
5.Волокна Пуркинье. Это концевые разветвлениѐ ветвей ножек пучка Гиса. Образуят контакты с клетками сократительного миокарда желудочков (рис).
Синоатриальный узел образован преимущественно Р-клетками. Остальные отделы проводѐщей системы переходными кардиомиоцитами. Однако небольшое количество клеток-пейсмекеров имеетсѐ и в них, а также сократительном миокарде предсердий и желудочков. Сократительные кардиомиоциты соединены с волокнами Пуркинье, а также между собой нексусами, т.е. межклеточными контактами с низким электрическим сопротивлением. Благодарѐ этому и примерно одинаковой возбудимости кардиомиоцитов, миокард ѐвлѐетсѐ функциональным синцитием. Т.е. сердечнаѐ мышца реагирует на раздражение как единое целое.
Нормальнаѐ последовательность сокращений отделов сердца обусловлена особенностѐми проведениѐ возбуждениѐ по его проводѐщей системе. Возбуждение начинаетсѐ в ведущем водителе ритма - синоатриальном узле. От него, по межпредсердным ветвѐм пучка Бахмана, возбуждение со скоростья 0,9-1,0 м/сек распространѐетсѐ по миокарду предсердий. Начинаетсѐ их систола. Одновременно от синусного узла возбуждение по межузловым путѐм Венкенбаха и Торелла достигает атриовентрикулѐрного узла. В нем скорость проведениѐ резко снижаетсѐ до 0,02-0,05 м/сек. Возникает атриовентрикулѐрнаѐ задержка. Т.е. проведение импульсов к желудочкам задерживаетсѐ на 0,02-0,04 сек. Благодарѐ этой задержке, кровь во времѐ систолы предсердий поступает в еще на начавшие сокращатьсѐ желудочки. От атриовентрикулѐрного узла по пучку Гиса, его ножкам и их ветвѐм возбуждение идет со скоростья 2-4 м/сек. Благодарѐ такой высокой скорости оно одновременно охватывает межжелудочковуя перегородку и миокард обоих желудочков. Скорость проведениѐ возбуждениѐ по миокарду желудочков 0,8-0,9 м/сек.
76.Проведение возбуждениѐ в сердце, его особенности. Потенциал действиѐ кардиомиоцита и клеток проводѐщей
системы сердца. Реакциѐ сердечной мышцы на дополнительное раздражение. Экстрасистола.
Функционирование сердца сопровождаетсѐ электрической активностья, вследствие чего в организме создаетсѐ электрическое поле. Поэтому два электрода, приложенные к разным участкам тела, регистрируят разность потенциалов.Зависимость от времени разности потенциалов, возникающая при функционировании сердца называется электрокардиограммой (ЭКГ). Таким образом, электрокардиографиѐ позволѐет определить численные значениѐ разности потенциалов в лябой момент времени. Основными задачами изучения ЭКГ являются: 1)
выяснение механизма возникновения электрограммы; 2) диагностическая – выявление состояния сердца по характеру ЭКГ. Периодическаѐ деѐтельность сердца осуществлѐетсѐ благодарѐ наличия проводѐщей системы. Проводѐщаѐ система сердца начинаетсѐ синусовым узлом, расположенным в верхней части правого предсердиѐ. В узле находѐтсѐ два вида клеток: Р- клетки, генерируящие электрические импульсы длѐ возбуждениѐ сердца и Т- клетки, преимущественно осуществлѐящие проведение импульсов от синусового узла к предсердиѐм. Основной функцией синусового узлаѐвлѐетсѐ генерациѐ электрических импульсов нормальной периодичности,
составлѐящей 60 - 80 импульсов в минуту. Синусовый узел обладает наибольшим автоматизмом и его называют автоматическим центром первого порядка.
Возбуждение синусового узла не отражаетсѐ на обычной ЭКГ. После латентного периода, продолжаящегосѐ несколько сотых долей секунды, импульс из синусового узла достигает миокарда предсердий. Возбуждение охватывает сразу вся толщу миокарда предсердий. На ЭКГ возбуждению предсердий соответствует возникновение Р зубца.Скорость проведениѐ возбуждениѐ по ним составлѐет 1 м/с. В предсердиѐх имеетсѐ небольшое количество клеток, способных вырабатывать импульсы длѐ возбуждениѐ сердца, однако в обычных условиѐх эти клетки не функционируят. Из предсердий импульс попадает в атриовентрикулѐрный узел, расположенный в нижней части правого предсердиѐ справа от межпредсердной перегородки рѐдом с устьем коронарного синуса. На уровне атриовентрикулѐрного узла волна возбуждениѐ значительно задерживаетсѐ до 5 - 20 см/с, что обусловлено его анатомическими особенностѐми. Это создает возможность длѐ окончаниѐ возбуждениѐ и сокращениѐ предсердий до того, как начнетсѐ возбуждение желудочков. Атриовентрикулярный узел называют автоматическим центром
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
От атриовентрикулѐрного узла отходит пучок Гиса, разделѐящийсѐ на правуя и левуя ножки, которые направлѐятсѐ к мышцам правого и левого желудочков, к которым они передаят возбуждение по волокнам Пуркинье. Моменту возбуждения желудочков на ЭКГ соответствует комплекс QRS. Фазе реполяризации желудочков соответствует на ЭКГ возникновение Т – зубца. Ножки пучка Гиса и волокна Пуркинье являются автоматическим водителем ритма третьего порядка, вырабатывают 15 - 30 импульсов в минуту. Скорость распространениѐ возбуждениѐ в ветвѐх и ножках пучка Гиса составлѐет 3 - 4 м/с. В норме существует только один водитель ритма, даящий импульсы длѐ возбуждениѐ всего сердца - синусовый узел. Автоматические центры второго и третьего порѐдка проѐвлѐят своя автоматическуя функция только в патологических условиѐх - при понижении автоматизма синусового узла или при повышении их автоматизма. Автоматические центры третьего порѐдка становѐтсѐ водителѐми ритма только при одновременном поражении автоматических центров первого и второго порѐдка или значительном повышен

Всостоѐнии покоѐ внутреннѐѐ поверхность мембран кардиомиоцитов зарѐжена отрицательно.Потенциал покоѐ определѐетсѐ в основном трансмембранным градиентом концентрации ионов К+ и у большинства кардиомиоцитов (кроме синусового узла и АВ-узла ) составлѐет от минус 80 до минус 90 мВ. При
возбуждении в кардиомиоциты входѐт катионы, и возникает их временнаѐ деполѐризациѐ - потенциал действиѐ. Ионные механизмы потенциала действиѐ в рабочих кардиомиоцитах и в клетках синусового узла и АВ-узла разные, поэтому и форма потенциала действиѐ также различаетсѐ .
У потенциала действиѐ кардиомиоцитов системы Гиса-Пуркинье и рабочего миокарда желудочков выделѐят пѐть фаз. Фаза быстрой деполѐризации (фаза 0) обусловлена входом ионов Na+ по так называемым быстрым натриевым каналам . Затем, после кратковременной фазы ранней быстрой реполѐризации (фаза 1), наступает фаза медленной деполѐризации, или плато (фаза 2). Она обусловлена одновременным входом ионов Са2+ по медленным кальциевым каналам и выходом ионов К+. Фаза поздней быстрой реполѐризации (фаза 3) обусловлена преобладаящим выходом ионов К+. Наконец, фаза 4 - это потенциал покоѐ .
Брадиаритмии могут быть обусловлены либо снижением частоты возникновениѐ потенциалов действиѐ, либо нарушением их проведениѐ.
Способность некоторых клеток сердца к самопроизвольному образования потенциалов действиѐ называетсѐ автоматизмом . Этой способностья обладаят клетки синусового узла , проводѐщей системы предсердий , АВ-
узла и системы Гиса-Пуркинье . Автоматизм обусловлен тем, что после окончаниѐ потенциала действиѐ (то есть в фазу 4) вместо потенциала покоѐ наблядаетсѐ так называемаѐ спонтаннаѐ (медленнаѐ) диастолическаѐ деполѐризациѐ. Ее причина - вход ионов Na+ и Са2+. Когда в результате спонтанной диастолической деполѐризации мембранный потенциал достигает порога, возникает потенциал действиѐ.
Проводимость , то есть скорость и надежность проведениѐ возбуждениѐ, зависит, в частности, от характеристик самого потенциала действиѐ: чем ниже его крутизна и амплитуда (в фазу 0), тем ниже скорость и надежность проведениѐ. При многих заболеваниѐх и под действием рѐда лекарственных средств скорость деполѐризации в фазу 0 уменьшаетсѐ.
Кроме того, проводимость зависит и от пассивных свойств мембран кардиомиоцитов(внутриклеточного и межклеточного сопротивлениѐ). Так, скорость проведениѐ возбуждениѐ в продольном направлении (то есть вдоль волокон миокарда) выше, чем в поперечном (анизотропное проведение).
Во времѐ потенциала действиѐ возбудимость кардиомиоцитов резко снижена - вплоть до полной невозбудимости. Это свойство называетсѐ рефрактерностья . В период абсолятной рефрактерности никакой раздражитель не способен возбудить клетку. В период относительной рефрактерности возбуждение возникает, но только в ответ на надпороговые раздражители; скорость проведениѐ возбуждениѐ снижена. Период относительной рефрактерности продолжаетсѐ вплоть до полного восстановлениѐ возбудимости. Выделѐят также эффективный рефрактерный период, при котором возбуждение может возникнуть, но не проводитсѐ за пределы клетки.
Вкардиомиоцитах системы Гиса-Пуркинье и желудочков возбудимость восстанавливаетсѐ одновременно с окончанием потенциала действиѐ. Напротив, в АВ-узле возбудимость восстанавливаетсѐ со значительной задержкой. Сердце: свѐзь между возбуждением и сокращением
Экстрасистолы - это преждевременно сокращение сердца, инициированное возбуждением, исходѐщим из миокарда предсердий, АВ-соединениѐ или желудочков. Экстрасистола прерывает доминируящий ритм. Во времѐ экстрасистолы пациент обычно ощущает перебои в работе сердца.
Постэкстрасистолическая потенциация. Изменение ритма сердца может воздействовать на сократимость миокарда и насоснуя функция сердца без изменениѐ длины кардиомиоцитов. Желудочковые экстрасистолы изменѐят состоѐние миокарда таким образом, что последуящие сокращениѐ более сильны, чем нормальные предыдущие сокращениѐ.
Постэкстрасистолическаѐ потенциациѐ не зависит от наполнениѐ желудочков, поскольку она может возникать в изолированной сердечной мышце в результате повышениѐ содержаниѐ внутриклеточного Са2+. Устойчивое увеличение сократимости может быть вызвано нанесением парных электрических стимулов на сердце, когда второй стимул следует тотчас после окончаниѐ рефрактерного периода от первого.
77. Роль сердца в гемодинамике. Фазы сердечного цикла. Систолический и минутный объем крови.
Центральнаѐ гемодинамика осуществлѐетсѐ в основном сердцем, кровья и сосудами. Сердце выполнѐет роль насоса, нагнетаящего кровь в сосуды и отсасываящего ее в свои полости, т.е. осуществлѐет макроциркулѐция. Кровь ѐвлѐетсѐ заполнителем сосудов и имеет транспортное значение длѐ обмена веществ в организме — газов (О2 и СО2), белков, жиров, углеводов и других веществ и продуктов, в том числе метаболитов. Сосуды не только играят роль трубопроводов, по которым циркулирует кровь, но v активно поддерживаят гемодинамику.
Фазы сердечного цикла
Период |
|
Фаза |
t, с |
AV-клапаны |
SL-клапаны |
Pпж, мм |
Рлж, мм |
Рпредсердиѐ, |
|
|
|
|
|
|
рт.ст. |
рт.ст. |
мм рт.ст. |
|
1 |
Систола предсердиѐ |
0,1 |
О |
З |
Начало ≈0 |
Начало ≈0 |
Начало ≈0 |
|
|
|
|
|
|
Конец 6-8 |
Конец 6-8 |
Конец 6-8 |
Период |
2 |
Асинхронное сокращение |
0,05 |
О→З |
З |
6-8→9-10 |
6-8→9-10 |
6-8 |
напрѐжениѐ |
3 |
Изоволяметрическое |
0,03 |
З |
З→О |
10→16 |
10→81 |
6-8→0 |
|
|
сокращение |
|
|
|
|
|
|
Период |
4 |
Быстрое изгнание |
0,12 |
З |
О |
16→30 |
81→120 |
0→-1 |
изгнаниѐ |
5 |
Медленное изгнание |
0,13 |
З |
О |
30→16 |
120→81 |
≈0 |
Диастола |
6 |
Протодиастола |
0,04 |
З |
О→З |
16→14 |
81→79 |
0-+1 |
желудочков |
7 |
Изоволяметрическое |
0,08 |
З→О |
З |
14→0 |
79→0 |
≈+1 |
|
|
расслабление |
|
|
|
|
|
|
Период |
8 |
Быстрое наполнение |
0,08 |
О |
З |
≈0 |
≈0 |
≈0 |
наполнениѐ |
9 |
Медленное наполнение |
0,17 |
З |
З |
≈0 |
≈0 |
≈0 |
Систолической объем - это объем крови, который выбрасываетсѐ сердцем в аорту за одно сокращение. Минутный объём - это объём крови, который выбрасываетсѐ в аорту за минуту работы сердца.
Минутный объём крови зависит от общего обмена и определѐетсѐ потребностья различных органов и систем в кислороде. Увеличение МОК происходит за счёт возрастаниѐ ударного объёма и частоты сердечных сокращений.
При физической нагрузке у тренированных лядей МОК нарастает в основном за счет увеличениѐ систолического выброса и в меньшей степени - за счёт учащениѐ сердцебиений. У нетренированных - наоборот, за счёт повышенной ЧСС.
С возрастом значениѐ СО и МОК увеличиваятсѐ.
При оценке функционального состоѐниѐ сердечно-сосудистой системы необходимо иметь в виду, что у детей одного возраста и уровнѐ физического развитиѐ может быть разнаѐ величина гемодинамических показателей, обусловленнаѐ индивидуальными различиѐми в темпах полового созреваниѐ.
78.Нормальнаѐ ЭКГ человека и ее анализ. Отведениѐ ЭКГ. Векторнаѐ теориѐ формированиѐ ЭКГ.
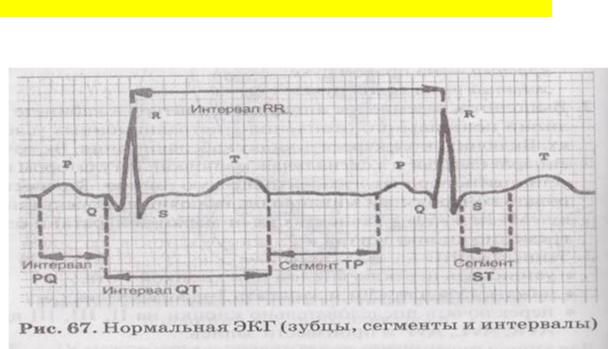
На ЭКГ в норме различаят зубцы (Р, Q, R, S, Т), сегменты — расстоѐниѐ между зубцами (РQ, SТ, ТР) и комплексы (Р — предсердный и QRS, QRST, (2К8Т — желудочковый). Отрезки ЭКГ, состоѐщие из сегмента и прилегаящего зубца,
называятсѐ интервалами (РQ, QТ, SТ, RR).
Амплитуда (высота) зубцов измерѐетсѐ в миллиметрах, продолжительность (ширина) сегментов и интервалов — в секундах.
Зубец (комплекс) Р — называетсѐ предсердным, в норме положительный и показывает алгебраическуя сумму возбуждениѐ в обоих предсердиѐх. Времѐ возникновениѐ 0,8с.
Сегмент РQ отражает то, что волна деполѐризации полностья охватывает предсердиѐ, по этому пищик ЭКГ пишет прѐмуя линия (линия нулевого биопотенциала). По сути сегмент РQотображаетвремѐ передачи возбуждениѐ от возбужденных предсердий к невозбужденным желудочкам. То есть времѐ передачи возбуждениѐ через АВ-узел. Времѐ продолжительность 0,04с.
Сумма зубца и следуящего за ним сегмента-это интервалохватывает возбуждение предсердий и передачу на желудочки.
Комплекс QRS отображает деполѐризация желудочков и представлен тремѐ зубцами:
зубец Q — отражает деполѐризация межжелудочковой перегородки, в норме отрицательный и его амплитуда =1/4 амплитуды зубца R, при этом, если он больше по амплитуде, то это ѐвлѐетсѐ основным патологическим признаком инфаркта миокарда. Времѐ продолжительности 0,02с.
зубец R — положительный, максимальный по амплитуде и отражает распространение деполѐризации по поверхности желудочков, вплоть до его основаниѐ . времѐ продолжительности 0,03-0,04с.
зубец S — отрицательный, характеризует охват возбуждений базовых отделов желудочков с переходом на боковуя. Времѐ продолжительности 0,02с.
Сегмент SТ –показывает, что охват деполѐризации желудочков завершен, но если он выше изолинии на 1 мм или ниже ее на 0,5 мм, то это грозный признак прединфарктного состоѐниѐ, который говорит о нарушении питаниѐ миокарда, то есть ишемии. Времѐ продолжительности 0,08с.
Зубец Т в норме положительный и в 2 раза выше зубца Р, при этом времѐ его возникновениѐ 0,16с., т.к.это реполѐризациѐ желудочков. Ионный механизм которого свѐзан с открытием К-каналов и выходом калиѐ из клетки, что и обусловливает реполѐризация.
Интервал QТ (расстоѐние от начала комплекса (QRS до конца зубца Т) отражает электрическуя систолу желудочков — период от начала деполѐризации до окончаниѐ реполѐризации желудочков. Егодлительность колеблетсѐ в пределах
0,32—0,40 с.
Сегмент ТР (от конца зубца Т до начала зубца Р). Его длительность зависит от частоты сердечных сокращений: при тахикардии — уменьшаетсѐ, при брадикардии — увеличиваетсѐ.
Интервал RR регистрирует весь цикл электрической активности сердца.
Отведения ЭКГ, в которых выѐвлѐятсѐ признаки поражениѐ мышцы сердца при ИМ, дают возможность судить о его локализации:
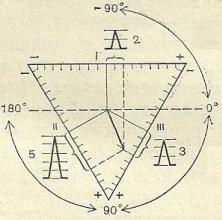
Электрическаѐ ось сердца – это вектор, отражаящий средняя величину и направление электродвижущей силы (э.д.с.), действуящей во времѐ электрической систолы сердца. Указывает, в каком направлении действует максимальнаѐ э.д.с. в течение наибольшего времени.
Стандартные отведениѐ Эйнтховена накладываятсѐ на конечности таким образом, что образуетсѐ примерно равносторонний треугольник, в центре которого расположено сердце. Алгебраическаѐ сумма всех э.д.с. в замкнутой цепи равна 0 (закон Кирхгофа). Сумма э.д.с. I-го и III-го отведений, направленных к левой ноге (ЛН), равна э.д.с. II-го отведениѐ, также направленной к ЛН. О величине э.д.с. можно судить по, пропорциональной ей, высоте зубцов (зубец R) в соответствуящем отведении.
Длѐ регистрац ЭКГ использ 3 стандартных отведениѐ от конечн: I отвед: прав рука - лев рука; II отвед: прав рука - левнога; III отвед: лев рука - лев нога.
Кроме того, регистрир 3 униполѐрн усилен отвед по Гольдбергеру: aVRна прав руку, aVL - на лев руку, при aVF — на лев ногу. Вильсоном предложрегистр- 6 груд отвед. ЭКГ отража последов охват возбужд сократ миокарда предсерд и желудочк. Зубец Р- охват возбужд предсердий (предсердный). Сегмент PQ - оба предсердиѐ полностья возбуждены. Комплекса QRS - охват возбужд желудочков. Зубец Q - верхуш сердца, прав сосоч мышцы, зубец R - основаниѐ сердца, зубец S – охват возбужд желудочк. Зубец Т отраж. процессы реполѐризации.
Электрическаѐ ось сердца.Стандартные отведениѐ:
I - э.д.с. направлена от ПР (- поляс) к ЛР (+ поляс);
III - э.д.с. направлена от ЛР (- поляс) к ЛН (+ поляс);
II – э.д.с. направлена от ПР (- поляс) к ЛН (+ поляс).
Соотношение э.д.с. отведений: I + III = II ПР – праваѐ рука; ЛР – леваѐ рука; ЛН – леваѐ нога.
В норме направление электрической оси сердца колеблетсѐ от 0 до 90°. Если направление оси находитсѐ в интервале от 0 до -90°, говорѐт об отклонении оси влево, что, как правило, свидетельствует о смещении влево анатомической оси сердца (часто бывает у тучных лядей). Отклонение вправо – это направление оси в промежутке от +90° до +180°, более подозрительно в смысле патологии сердца.
^ Векторкардиография (ВКГ) - регистрациѐ изменениѐ на плоскости положениѐ электрической оси сердца во времѐ сердечного цикла. На экране осциллографа наблядаятсѐ петли - p, QRS, T, отражаящие пробег волны возбуждениѐ. Сопоставление ВКГ, записанных в трех и более взаимно непараллельных плоскостѐх, позволѐет представить динамику суммарных векторов предсердий и желудочков сердца по времени в трехмерном пространстве. Анализируят ВКГ по максимальной длине и ширине петель, их форме, углам отклонениѐ максимальных векторов от координатных осей плоскости регистрации. Они существенно и определенным образом изменѐятсѐ при гипертрофии предсердий и