

Биохимия в таблицах, схемах и графиках
.pdfБИОСИНТЕЗ ЖИРНЫХ КИСЛОТ
Процесс осуществляется в различных частях клетки. В цитоплазме синтезируются насыщенные жирные кислоты с углеродной цепью до С16 (пальмитат).
Субстратом для синтеза жирной кислоты служит ацетил-СоА, образующийся в результате окисления пирувата. Использование пирувата в синтезе пальмитата включает следующие метаболические пути:
-окисление глюкозы в процессе гликолиза до пирувата в цитозоле,
-окислительное декарбоксилирование пирувата в митохондриях и последующая конденсация ацетил-СоА с оксалоацетатом с образованием цитрата.
Далее цитрат перемещается в цитозоль, где распадается на ацетил-СоА и оксалоацетат. Существуют механизмы транспорта АцетилКоА из митохондрий в цитоплазму.
71
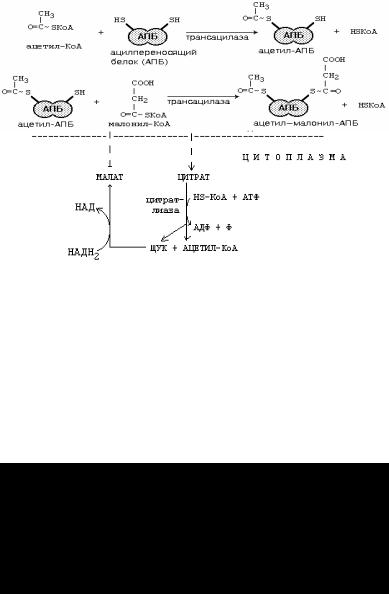
Условием для выхода Ацетил-КоА из митохондрии в цитоплазму является хорошее обеспечение клетки АТФ. Если АТФ в клетке мало, то Ацетил-КоА расщепляется до СО2 и Н2О.
Первая реакция в биосинтезе жирной кислоты - это карбоксилирование ацетил-СоА и превращение его в малонил-СоА. Реакция катализируется ацетил-СоА-карбоксилазой, коферментом которой является биотин. Затем следуют повторяющиеся циклы из шести реакций.
Катализирует весь процесс пальмитилсинтетаза - полифункциональный белок - фермент, имеющий одну полипептидную цепь, упакованную в два домена.
РЕАКЦИИ СИНТЕЗА ЖИРНЫХ КИСЛОТ.
72
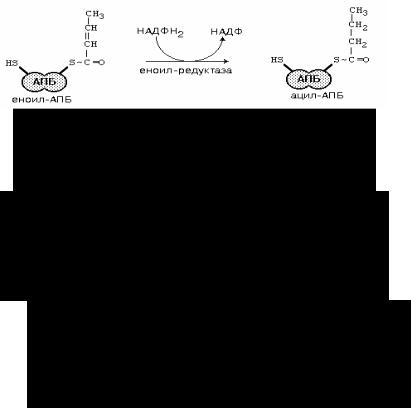
Очередной цикл синтеза начинается с присоединения новой молекулы малонил-СоА к одной из SH-групп активных центров доменов.
Каждый цикл увеличивает длину цепи на два углеродных атома. Когда цепь достигает длины 16 углеродных атомов, тиоэфирная связь гидролизуется и пальмитат освобождается.
Суммарное уравнение синтеза пальмитата (7 циклов):
Ацетил-СоА+7Малонил-КоА+14(NADPH+Н+) → пальмитат + 8HS-CoA+
7CO2+14NADP++7H2O
В митохондриях происходит дальнейшее наращивание цепи, а в ретикулуме насыщенные жирные кислоты превращаются в ненасыщенные, и также происходит удлинение цепи.
73
МЕТАБОЛИЗМ ХОЛЕСТЕРИНА И ЖЕЛЧНЫХ КИСЛОТ
Функции холестерина:
1)холестерин является предшественником в синтезе других стероидов: желчных кислот, стероидных гормонов, витамина D3;
2)холестерин входит как структурный компонент в состав мембран всех клеток.
Существует два пути поступления холестерина:
1)из пищи животного происхождения (экзогенный холестерин;
2)синтез в печени (эндогенный холестерин).
Кроме печени в небольшом количестве холестерин может синтезироваться в клетках кишечника и кожи.
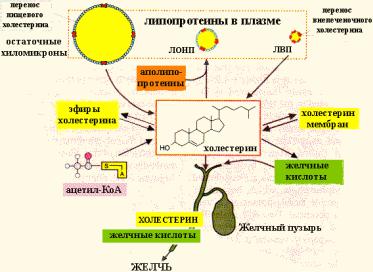
ПУТИ ИСПОЛЬЗОВАНИЯ ХОЛЕСТЕРИНА
БИОСИНТЕЗ ХОЛЕСТЕРИНА
Процесс происходит в цитозоле клетки.
Молекула холестерина целиком "собирается" из ацетил-СоА. Скорость синтеза холестерина зависит от количества экзогенного холестерина, то есть поступающего с пищей. При поступлении 2-3 г холестерина в сутки синтез эндогенного холестерина подавляется.
74
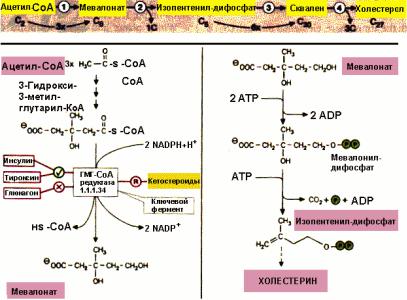
Фермент гидроксиметилглутарил-СоА-редуктаза играет главную роль в регуляции синтеза холестерина. Холестерин подавляет синтез ГМГ-СоА-редуктазы и таким образом по механизму отрицательной обратной связи снижает скорость своего синтеза.
ТРАНСПОРТ ХОЛЕСТЕРИНА
В печени пул холестерина составляется из синтезированного холестерина самими клетками и поступившего из остаточных хиломикронов. Этот пул холестерина существует не только для собственных нужд печени, но и для снабжения других тканей. Холестерин печени вместе с жирами, синтезированными из глюкозы, включается в ЛОНП и таким образом транспортируется кровью. После гидролиза жиров ЛП-липазой образуются ЛОНП остаточные. Эти липопротеины либо поглощаются печенью, либо превращаются в ЛНП.
 Почти во всех клетках имеются рецепторы для апоВ100. Поэтому ЛНП фиксируются на поверхности клеток. При этом наблюдается переход холестерина в клеточные мембраны. Таким образом, ЛНП способны снабжать холестерином клетки тканей.
Почти во всех клетках имеются рецепторы для апоВ100. Поэтому ЛНП фиксируются на поверхности клеток. При этом наблюдается переход холестерина в клеточные мембраны. Таким образом, ЛНП способны снабжать холестерином клетки тканей.
Транспортируют холестерин из тканей в печень липопротеины высокой плотности (ЛВП). В кровеносном русле к ЛВП присоединяется
75
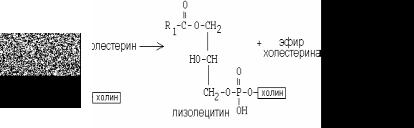
белок-фермент лецитинхолестеринацилтрансфераза (ЛХАТ). АпоА является активатором ЛХАТ.
ЛХАТ катализирует следующую реакцию:
Реакция важна, потому что образующийся эфир холестерина является очень гидрофобным веществом и сразу переходит в ядро ЛВП – так, при контакте с мембранами клеток ЛВП удаляют из них избыток холестерина. Дальше ЛВП идут в печень, там разрушаются, и избыток холестерина удаляется из организма.
Нарушение соотношения между количеством ЛНП, ЛОНП и ЛВП может вызывать задержку холестерина в тканях. Это приводит к атеросклерозу. Поэтому ЛНП называют атерогенными липопротеинами,
а ЛВП - антиатерогенными липопротеинами.
ЭЙКОЗАНОИДЫ
Биологически активные вещества, синтезируемые большинством клеток из полиеновых жирных кислот, содержащих 20 углеродных атомов.
Эйкозаноиды разделяют на классы: а) простагландины (включая простациклины); б) тромбоксаны; в) лейкотриены.
БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ОСНОВНЫХ ТИПОВ ЭЙКОЗАНОДОВ
Эйкозаноиды |
Основное место |
Основное биологическое действие |
|||
|
снтеза |
|
|
|
|
РGЕ2 |
Большинство |
Расслабляет |
гладкую |
мускулатуру, |
|
|
тканей, |
особенно |
расширяет |
сосуды, |
инициирует |
|
почки |
|
родовую активность |
|
|
|
|
|
|
|
|
РGF2α |
Большинство |
Сокращает |
гладкую |
мускулатуру, |
|
|
тканей |
|
сужает |
сосуды, |
бронхи, |
|
|
|
стимулирует сокращение матки |
||
|
|
|
|
|
|
|
|
76 |
|
|
|
РGI2 |
Сердце, |
клетки |
Уменьшает |
агрегацию |
||
|
эндотелия сосудов |
тромбоцитов, расширяет сосуды, в |
||||
|
|
|
клетках-мишенях |
увеличивает |
||
|
|
|
образование сАМФ |
|
||
ТХА2 |
Тромбоциты |
|
Стимулирует |
агрегацию |
||
|
|
|
тромбоцитов, сужает сосуды, в |
|||
|
|
|
клетках |
уменьшает образование |
||
|
|
|
сАМФ |
|
|
|
ТХВ2 |
Тромбоциты |
|
Сужает сосуды |
|
||
LТВ4 |
Клетки белой крови, |
Стимулирует |
хемотаксис |
и |
||
|
клетки эпителия |
агрегацию лейкоцитов |
|
|||
LТС4→LТD4 |
Клетки белой крови, |
Стимулирует расширение сосудов, |
||||
|
альвеолярные |
|
увеличивают их |
проницаемость, |
||
|
макрофаги |
|
вызывают |
сокращение бронхов. |
||
|
|
|
Основные компоненты медленно |
|
||
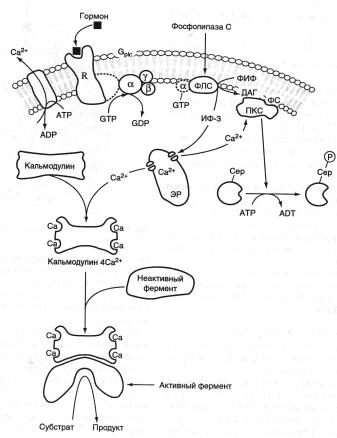
ТРАНСМЕМБРАННАЯ ПЕРЕДАЧА СИГНАЛА
Мембраны способны воспринимать (наличие рецептора) и передавать внутрь клетки сигналы из внешней среды.
Внеклеточными химическими сигналами могут быть гормоны нейромедиаторы, эйкозаноиды или другие сигнальные молекулы.
Гормоны – это молекулы, которые вырабатываются специализированными клетками, секретируются в кровь в ответ на изменение какого-либо специфического параметра внутренней среды организма и оказывают влияние на метаболизм и функциональное состояние клеток мишеней.
По механизму передачи сигнала в клетку различают гормоны, взаимодействующие с мембранными рецепторами, и гормоны, передающие сигнал через внутриклеточные рецепторы.
Взаимодействие большинства гормонов, эйкозаноидов и др. с рецептором приводит к активации внутриклеточных регуляторных систем, в частности аденилатциклазной и инозитолфосфатной.
АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА
Включает 5 мембранных белков: рецептор активатора (RS), рецептор ингибитора (Ri), GS (стимулирующий белок) и Gi (ингибирующий белок), фермент аденилатциклазу (АЦ) и цитозольный фермент протеинкиназу А – ПКА (цАМФ-зависимую протеинкиназу).
77

ИНОЗИТОЛФОСФАТНАЯ СИСТЕМА
Включает 3 основных мембранных белка: R (рецептор), фосфолипазу С и Gрlс – белок, активирующий фосфолипазу С, а также белки и ферменты мембран цитозоля, участвующие в связывании и транспорте Са2+ .
78
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Нуклеиновые кислоты - гетерополимеры, их мономерами являются мононуклеотиды.
Роль нуклеиновых кислот
1.ДНК: хранение генетической информации.
2.РНК:
а) хранение генетической информации у некоторых вирусов; б) реализация генетической информации: м-РНК – матричная РНК, т-РНК (транспортная), р-РНК (рибосомальная);
79
в) некоторые молекулы РНК способны катализировать реакции гидролиза 3’,5’- фосфодиэфирной связи в самой молекуле РНК. Такие РНК называют рибозимами.
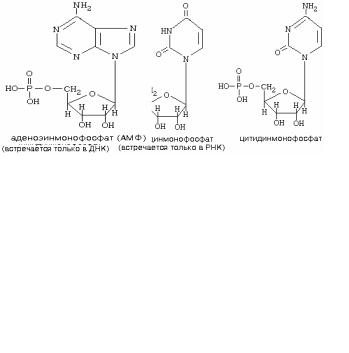
Мононуклеотид состоит из азотистого основания + рибоза у РНК (или дезоксирибоза у ДНК) - вместе они составляют нуклеозид, и остатка фосфорной кислоты.
НОМЕНКЛАТУРА НУКЛЕОТИДОВ
Азотистое основание |
Нуклеозид |
Нуклеотид |
||||
аденин |
аденозин |
Аденозинмонофосфат (АМФ) |
||||
гуанин |
гуанозин |
Гуанозинмонофосфат (ГМФ) |
||||
урацил |
уридин |
уридинмонофосфат (УМФ) |
||||
тимин |
тимидин |
тимидинмонофосфат (ТМФ) |
||||
цитозин |
цитидин |
цитидинмонофосфат (ЦМФ) |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
80