

Биохимия в таблицах, схемах и графиках
.pdfДля активного действия панкреатической липазы необходимы следующие условия:
-рН, близкое к нейтральной среде;
-желчные кислоты, эмульгирующие жиры;
-белок колипаза, синтезируемый в поджелудочной железе и секретируемый вместе с панкреатической липазой.
3.Образование смешанных мицелл и всасывание продуктов гидролиза. Продукты гидролиза липидов – жирные кислоты с длинным
углеводородным радикалом, β-моноацилглицерины, холестерол, а также соли желчных кислот образуют в просвете кишечника структуры, называемые смешанными мицеллами. Смешанные мицеллы построены таким образом, что гидрофобные части молекул обращены внутрь мицеллы, а гидрофильные – наружу, поэтому мицеллы хорошо растворяются в водной фазе содержимого тонкого кишечника. Стабильность мицелл обеспечивается в основном солями желчных кислот. Мицеллы сближаются со щеточной каймой клеток слизистой оболочки тонкого кишечника, и липидные компоненты мицелл диффундируют через мембраны внутрь клеток. Вместе с продуктами гидролиза липидов всасываются жирорастворимые витамины А,D, Е, К и соли желчных кислот.
ЖЕЛЧНЫЕ КИСЛОТЫ
желчные кислоты выполняют 2 основные функции:
-эмульгирование жира
-всасывание жирных кислот.
Вкишечник желчные кислоты поступают в составе желчи в виде конъюгатов с глицином и таурином. После переваривания и всасывания желчные кислоты возвращаются через воротную вену в печень, совершая такой цикл до 10 раз в сутки. Этот цикл называется энтерогепатическая
циркуляция или кишечно-печеночная циркуляция желчных кислот.
61

Постоянным компонентом желчи является холестерин. Как и желчные кислоты, он подвергается обратному всасыванию, но некоторое количество желчных кислот и холестерина теряются с калом. Для восполнения потери желчных кислот, выводимых с фекалиями, постоянно происходит синтез желчных кислот из холестерина.
62

4. Ресинтез жиров в слизистой оболочке тонкого кишечника.
Первая стадия ресинтеза жиров – реакция активации жирных кислот:
НSКоА + RСООН + АТФ → R-СО~КоА + АМФ + Н4Р2О7.
Реакция катализируется ферментом ацил~КоА-синтетазой. В ресинтезе жиров участвуют не только жирные кислоты, всосавшиеся из кишечника, но и жирные кислоты, синтезированные в организме, поэтому по составу реснтезированные жиры отличаются от жиров, полученных с пищей.
В клетках эпителия тонкой кишки из жиров, образовавшихся в результате ресинтеза, а также из эфиров холестерола, жирорастворимых витаминов, поступивших с пищей, формируются липопротеиновые комплексы – хиломикроны (ХМ). ХМ далее доставляют жиры в периферические ткани.
ТРАНСПОРТ ЛИПИДОВ
Липиды, в частности жиры, не растворяются в водных фазах организма, поэтому транспорт липидов кровью и лимфой осуществляется в виде комплексов с белками и фосфолипидами, которые называются липопротеинами.
Липопротеины - это сферические частицы, в которых можно выделить гидрофобную сердцевину, состоящую из триглицеридов (ТАГ) и эфиров холестерина (ЭХС) и амфифильную оболочку, в составе которой – фосфолипиды, гликолипиды и белки (апобелки).
СТРОЕНИЕ ЛИПОПРОТЕИНОВ
Липопротеины разделяются методом ультрацентрифугирования соответственно их плотности на четыре основных типа:
63

В кишечнике образуются два типа липопротеинов: хиломикроны - ХМ и в небольшом количестве липопротеины очень низкой плотности - ЛОНП. В составе хиломикронов экзогенные жиры доставляются в органы и ткани.
ЛИПОПРОТЕИНЫ - ТРАНСПОРТНЫЕ ФОРМЫ ЛИПИДОВ В
ОРГАНИЗМЕ ЧЕЛОВЕКА
|
Хиломик- |
ЛОНП |
|
ЛПП |
ЛНП |
ЛВП |
|
роны |
|
|
|
|
|
Состав, %: |
|
|
|
|
|
|
белки |
2 |
10 |
|
11 |
22 |
50 |
ФЛ |
3 |
18 |
|
23 |
21 |
27 |
ХС |
2 |
7 |
|
8 |
8 |
4 |
ЭХС |
3 |
10 |
|
30 |
42 |
16 |
ТАГ |
85 |
55 |
|
26 |
7 |
3 |
|
|
|
|
|
|
|
|
|
|
64 |
|
|
|
|
Транспорт |
|
Транспорт |
Промежуто- |
Транс- |
|
Транспорт |
|
Функция |
липидов |
из |
липидов, |
чная форма |
порт |
|
холесте- |
|
|
клеток |
ки- |
синтезируемы |
превращения |
холесте- |
|
рина из |
|
|
шечника |
|
х в печени |
ЛОНП в ЛНП |
рина |
в |
тканей в |
|
|
|
|
|
|
ткани |
|
печень. |
|
|
|
|
|
|
|
|
Удаление |
|
|
|
|
|
|
|
|
избытка |
|
|
|
|
|
|
|
|
холесте- |
|
|
|
|
|
|
|
|
рина из |
|
|
|
|
|
|
|
|
клеток. |
|
|
|
|
|
|
|
|
Донор |
|
|
|
|
|
|
|
|
апопро- |
|
|
|
|
|
|
|
|
теинов |
|
Место |
Эпителий |
|
Клетки пе- |
Кровь |
Плазма |
|
В |
клет- |
образова- |
тонкой |
|
чени |
|
крови |
|
ках |
пе- |
ния |
кишки |
|
|
|
(из |
|
чени – |
|
|
|
|
|
|
ЛОНП) |
|
ЛВП- |
|
|
|
|
|
|
|
|
предше- |
|
|
|
|
|
|
|
|
ствен- |
|
|
|
|
|
|
|
|
ники |
|
Плотность, |
0,92 – 0,98 |
0,96 – 1,0 |
|
1,00– 1,06 |
|
1,06 – 1,21 |
||
г/мл |
|
|
|
|
|
|
|
|
Диаметр |
Более 120 |
30 - 100 |
|
21 - 100 |
|
7 - 15 |
||
частиц, нм |
|
|
|
|
|
|
|
|
Основные |
В – 48 |
|
В – 100 |
В -100 |
В - 100 |
|
А |
|
аполипоп- |
С ІІ |
|
С – ІІ |
Е |
|
|
С – |
ІІ |
ротеины |
Е |
|
Е |
|
|
|
Е |
|
Примечание. ХМ – хиломикроны; ФЛ – фосфолипиды; ХС – холестерин, ЭХС – эфиры холестерина. Функции аполипопротеинов: В48 – основной белок ХМ; В-100 основной белок ЛОНП и ЛНП, взаимодействует с рецепторами ЛНП; С – ІІ – активатор липопротеинлипазы, переносится с ЛВП на ХМ и ЛОНП в крови; Е – взаимодействует с рецепторами ЛНП.
Существуют также промежуточные формы в метаболизме липопротеинов: хиломикроны остаточные (ХМост), ЛОНП остаточные (или липопротеины средней плотности - ЛСП).
Липопротеинлипаза (ЛП-липаза) - фермент, обеспечивающий потребление экзогенных жиров тканями. ЛП-липаза, располагающаяся в эндотелии сосудов, взаимодействует с хиломикронами кровотока и гидролизует триацилглирины на глицерин и жирные кислоты, которые поступают в клетку. По мере извлечения ТАГ из хиломикронов последние превращаются в остаточные хиломикроны и затем поступают в печень. Жирные кислоты поступают либо в жировую ткань, либо в мышечную.
65

ТРАНСПОРТНАЯ ФУНКЦИЯ ЛИПОПРОТЕИНОВ
КАТАБОЛИЗМ ЖИРА
Жиры хранятся до момента их использования. Катаболизм жира идет в три этапа:
1.Гидролиз жира до глицерина и жирных кислот (липолиз).
2.Превращение глицерина (вступает в реакции обмена
глюкозы) и жирных кислот (подвергаются β-окислению) в ацетил-КоА.
3.Общий путь – цикл трикарбоновых кислот.
Процесс липолиза известен как МОБИЛИЗАЦИЯ ЖИРА. Мобилизация жира - это реакция гидролиза жира до глицерина и жирных кислот. Это ферментативный процесс. Осуществляют его два фермента:
ЛИПАЗА ЖИРОВОЙ ТКАНИ и МОНОГЛИЦЕРИДЛИПАЗА.
Ключевым ферментом является липаза жировой ткани. Она регулируется гормонами, поэтому часто ее называют
«ГОРМОНЧУВСТВИТЕЛЬНАЯ ЛИПАЗА».
ПУТИ МЕТАБОЛИЗМА ГЛИЦЕРИНА
66
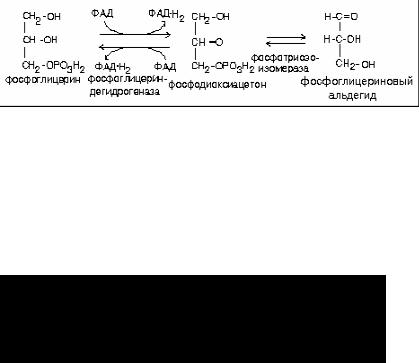
1.Глицерин может быть повторно использован для синтеза жира или других липидов.
2.Глицерин может вступить в обмен углеводов.
В любом случае в первую очередь происходит активация глицерина. Она похожа на активацию углеводов.
РАСПАД ГЛИЦЕРИНА НА ПУТИ К УГЛЕВОДАМ
Если глицерин распадается по пути к углеводам, то происходит дегидрирование.
Для фосфоглицеринового альдегида (ФГА) существует два варианта дальнейших превращений:
1.ФГА может окисляться в аэробном гликолизе до СО2 и Н2О с образованием 21 молекулы АТФ.
2.ФГА может вступить в реакции глюконеогенеза с образованием глюкозы.
ПУТИ ИСПОЛЬЗОВАНИЯ ЖИРНЫХ КИСЛОТ
Жирные кислоты могут вступать в реакции только после активации.
АКТИВНАЯ ФОРМА ЛЮБОЙ ЖИРНОЙ КИСЛОТЫ – АЦИЛ-КОА.
Для активной жирной кислоты, как и для глицерина, возможны два пути метаболических превращений:
1.Синтез жира или других липидов.
2.Катаболизм до Ацетил-КоА. Этот процесс называют β-окисление
67

КАТАБОЛИЗМ ЖИРНЫХ КИСЛОТ
Активация ЖК происходит в цитоплазме, а β-окисление – в митохондриях.
Ацил-КоА не может проходить через мембрану митохондрий. Поэтому имеется специальный механизм транспорта ЖК из цитоплазмы в митохондрию при участии вещества "КАРНИТИН". Во внутренней мембране митохондрий есть специальный транспортный белок, обеспечивающий перенос. Благодаря этому ацилкарнитин легко проникает через мембрану митохондрий.
Если жирная кислота попадает в митохондрию, то она обязательно подвергнется катаболизму до ацетил-КоА
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
Процесс β-окисления является циклическим. За каждый оборот цикла от жирной кислоты отщепляется 2 углеродных атома в виде ацетильного остатка. После этого укороченный на 2 углеродных атома ацил-КоА снова подвергается окислению (вступает в новый цикл реакций β-окисления). Образующийся Ацетил-КоА может дальше вступить в цикл трикарбоновых кислот.
68

Ускорение β-окисления наблюдается во многих тканях, в том числе и в печени при состоянии углеводного голодания и интенсивной мышечной работе. В печени образуется больше Ацетил-КоА, чем ей требуется. Поэтому в печени из Ацетил-КоА синтезируются специальные вещества, которые называются "КЕТОНОВЫЕ ТЕЛА".
69

РЕАКЦИИ СИНТЕЗА КЕТОНОВЫХ ТЕЛ
В норме концентрация кетоновых тел в крови 2 мг/дл.
Ацетон, который образуется при спонтанном (неферментативном) декарбоксилировании ацетоацетата, в организме не используется.
Кетоновые тела - ацетоацетат и бета-гидроксибутират- синтезируются в печени, легко проходят через митохондриальные и клеточные мембраны и поступают в кровь. Кровью они транспортируются во все другие ткани.
УТИЛИЗАЦИЯ КЕТОНОВЫХ ТЕЛ
Происходит в митохондриях (кроме клеток печени). β-гидроксибутират превращается в ацетоацетат, а ацетоацетат вступает в реакцию с промежуточным продуктом ЦТК - сукцинил-КоА.
70