

14. Регуляция дыхания, Дыхательный центр (Легаллуа, Флуранс, h.А. Миславский). Современные представления о локализации и строении дыхательного центра.
Дыхательный центр - совокупность нейронных ансамблей, расположенных на разных этажах центральной нервной системы, и обеспечивающих управление внешним дыханием.
Автоматический дыхательный центр - совокупность нейронов специфических (дыхательных) ядер продолговатого мозга, способных генерировать дыхательный ритм.
Функции ДЦ: моторная или двигательная(адаптирует дыхание к метаболическим потребностям организма, приспосабливает дыхание к поведенческим реакциям (бег, ходьба и др.), осуществляет интеграцию дыхания с др. функциями ЦНС.) , гомеостатическая (Поддерживает нормальные величины рО2 , рСО2 и рН в крови и внеклеточной жидкости. Регулирует дыхание при изменении t0 тела, адаптирует дыхательную функцию к изменению газовой среды (пониженное и повышенное атмосферное давление).
Уровни организации ДЦ:
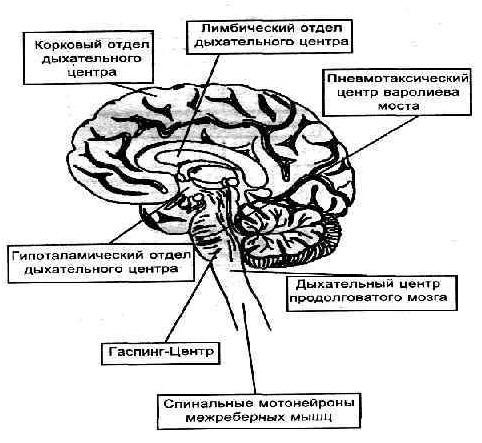
Французские ученые С. Легаллуа (1812) и М. Флурано (1842) используя метод перерезок пришли к выводу, что ДЦ локализован в продолговатом мозге, а введение внего иглы приводило к смерти животного.
Н.А. Миславский (1885) показал, что ДЦ — парное образование, имеющее инспираторные и экспираторные отделы.
Обнаружены некоторые особенности в работе дыхательного центра. При спокойном дыхании активна только небольшая часть дыхательных нейронов и, следовательно, в дыхательном центре есть резерв нейронов, который используется при повышенной потребности организма в кислороде. Установлено, что между инспираторными и экспираторными нейронами дыхательного центра существуют функциональные взаимосвязи. Они выражаются в том, что при возбуждении инспираторных нейронов, обеспечивающих фазу вдоха, деятельность экспираторных нервных клеток заторможена и наоборот. Таким образом, одной из причин ритмичной, автоматической деятельности дыхательного центра являются взаимосвязанные функциональные отношения между вдыхательными и выдыхательными нейронами.
+Существуют и другие представления о локализации и организации дыхательного центра, которые поддерживают ряд советских и зарубежных физиологов. Предполагают, что в продолговатом мозге локализованы центры вдоха, выдоха и судорожного дыхания. В верхней части моста мозга (варолиев мост) находится пневмотаксический центр, который контролирует деятельность расположенных ниже центров вдоха и выдоха и обеспечивает правильное чередование циклов дыхательных движений.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III-IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах грудных сегментов спинного мозга (III-XII)
15. Нервно-рефлекторный механизм регуляции дыхания. Роль центральных и периферических рецепторов в регуляции дыхания (Гейманс). Значение афферентной импульсации от механорецепторов легких в саморегуляции дыхания (рефлексы Геринга- Брейера).
Рефлекторная регуляция дыхания – Осуществляется благодаря тому, что нейроны дыхательного центра имеют связи с многочисленными механорецепторами дыхательных путей и альвеол легких и рецепторов сосудистых рефлексогенных зон. В легких человека находятся следующие типы механорецепторов: рецепторы растяжения гладких мышц дыхательных путей; ирритантные, или быстроадаптирующиеся, рецепторы слизистой оболочки дыхательных путей; J-рецепторы.
Центральные хеморецепторы (нервные клетки стволовой части мозга) регистрируют в межклеточной жидкости мозга pH и PСО2, они особенно чувствительны к увеличению PСО2 (гиперкапнии), а часть из них к уменьшению pH (ацидозу). Существенно, что центральные хеморецепторы расположены кнутри от гематоэнцефалического барьера, т.е. они отделены от крови в системе общей циркуляции (в частности, находятся в более кислой среде).
Периферические хеморецепторы(каротидные и аортальные тельца) регистрируют в артериальной крови pH, PО2 (PaО2) и PСО2, они особенно чувствительны к уменьшению PО2 (гипоксемии) и в меньшей степени — к увеличению PСО2 (гиперкапнии) и уменьшению pH (ацидозу).
Каротидный синус — расширение просвета внутренней сонной артерии тотчас у места её ответвления от общей сонной артерии. В стенке артерии области расширения присутствуют многочисленные барорецепторы, регистрирующие значения АД и передающие эту информацию в ЦНС по нервным волокнам, проходящим в составе синусного нерва (Херинга) — ветви языкоглоточного нерва.
Каротидное тельце – находится в области бифуркации общей сонной артерии. Клубочек каротидного тельца состоит из 2–3 клеток типа I (гломусные клетки), окружённых поддерживающими клетками (тип II). Клетки типа I образуют синапсы с терминалями афферентных нервных волокон. Каротидное тельце состоит из скоплений клеток (клубочков, гломусов), погружённых в густую сеть кровеносных капилляров (интенсивность перфузии телец наибольшая в организме, в 40 раз больше перфузии головного мозга). Каждый клубочек содержит 2–3 хемочувствительные гломусные клетки, образующие синапсы с терминальными разветвлениями нервных волокон синусного нерва — ветви языкоглоточного нерва. В тельцах также находятся нервные клетки симпатического и парасимпатического отделов вегетативной нервной системы. На этих нейронах и на гломусных клетках заканчиваются преганглионарные симпатические и парасимпатические нервные волокна, а на гломусных клетках также постганглионарные нервные волокна из верхнего шейного симпатического узла [терминали этих волокон содержат светлые (ацетилхолин) или гранулярные (катехоламины) синаптические пузырьки]. Гломусные клетки связаны друг с другом при помощи щелевых контактов, их плазмолемма содержит потенциалозависимые ионные каналы, клетки могут генерировать ПД и содержат разные синаптические пузырьки, содержащие ацетилхолин, дофамин, норадреналин, вещество P и метионин–энкефалин. Механизм регистрации DPО2, DPСО2 и DpH окончательно не установлен, но приводит к блокаде K+ каналов, что вызывает деполяризацию плазмолеммы гломусных клеток, открытию потенциалозависимых Ca2+ каналов, внутриклеточному увеличению [Ca2+] и секреции нейромедиаторов.
Аортальные (парааортальные) тельца рассыпаны по внутренней поверхности дуги аорты и содержат гломусные хемочувствительные клетки, образующие синапсы с афферентами блуждающего нерва.
Рефлексы Геринга- брейера
Раздувание легких у наркотизированного животного рефлекторно тормозит вдох и вызывает выдох. Перерезка блуждающих нервов устраняет рефлекс.
Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются волокнами блуждающего нерва.
Рефлекс Геринга — Брейера контролирует глубину и частоту дыхания. У человека он имеет проявляется при дыхательных объемах свыше 1 л (например, при физической нагрузке). Его физиологическое значение состоит в предотвращении перерастяжения лёгких . У новорожденных рефлекс Геринга — Брейера четко проявляется только в первые 3—4 дня после рождения
16. Гуморальный механизм регуляции дыхания. Влияние на дыхательный центр газового состава крови (опыт Фредерика), рH. некоторых электролитов и биологически активных веществ. Механизм первого вдоха новорожденного.
рО2 и рСО2 в артериальной крови поддерживается на достаточно стабильном уровне, несмотря на значительные изменения потребления О2 и выделение СО2.
• Гипоксия и понижение рН крови (ацидоз) вызывают усиление вентиляции (гипервентиляция), а гипероксия и повышение рН крови (алкалоз) — понижение вентиляции (гиповентиляция) или апноэ. Контроль за нормальным содержанием во внутренней среде организма О2, СО2 и рН осуществляется периферическими и центральными хеморецепторами.
• Адекватным раздражителем для периферических хеморецепторов является уменьшение рО2 в артериальной крови, в меньшей степени увеличение рСО2 и рН.
• Для центральных хеморецепторов — увеличение концентрации Н+ во внеклеточной жидкости мозга.
ОПЫТ:
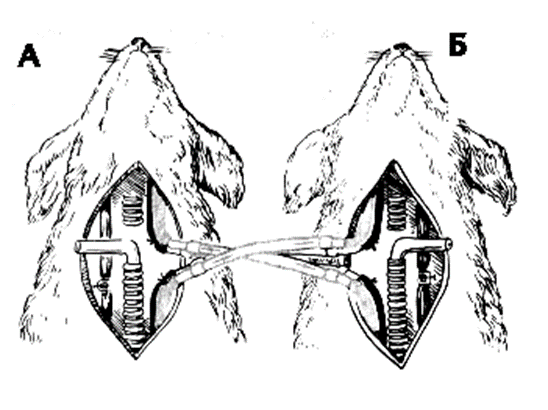
У двух собак, находившихся под наркозом, перерезали и соединяли перекрестно сонные артерии и отдельно яремные вены. После такого соединения и перевязки позвоночных артерий голова первой собаки снабжалась кровью второй и наоборот. Если у первой собаки перекрывали трахею и вызывали таким путем асфиксию, то у второй собаки развивалось гиперпноэ- увеличение легочной вентиляции. У первой же собаки, несмотря на увеличение в крови напряжения углекислоты и снижение напряжения кислорода через некоторое время наступалоапноэ- прекращение дыхательных движений. Это объясняется тем, что в сонную артерию первой собаки поступает кровь второй собаки, у которой в результате гипервентиляции снижается содержание углекислоты в артериальной крови. Уже тогда было установлено, что регуляция дыхания происходит путем обратной связи: отклонения в газовом составе артериальной крови приводят путем воздействия на дыхательный центр такие изменения дыхания, которые уменьшают эти отклонения.
Пережатие трахеи у собаки А вызывает одышку у собаки Б. Одышка у собаки Б вызывает замедление и остановку дыхания у собаки А
Механизм первого вдоха новорожденного.
Первый вдох - Происходит в момент пережатия пуповины, вследствие резкой стимуляции хеморецепторов благодаря, быстрому накоплению в крови CO2 и нарастающей гипоксии. Факторами, вызывающими первый вдох новорожденного являются:
1. Появление в крови таких стимуляторов дыхательного центра как избыток углекислого газа, низкая рН и недостаток кислорода. Наиболее быстро всё это нарастает при прекращении кровообращения через пуповину.
2. Мощный поток афферентных импульсов от рецепторов кожи (температурных, тактильных) и проприорецепторов, в процессе родов и сразу после рождения. Эти импульсы активируют ретикулярную формацию ствола мозга, что повышает возбудимость дыхательного центра.
3. Устранение раздражения рецепторов, расположенных в области ноздрей. Данные рецепторы посылают в дыхательный центр сигналы о наличии жидкости у входа в полость носа и предотвращают вдох (рефлекс «ныряльщика»). Поэтому сразу при появлении головки плода из родовых путей, акушеры удаляют слизь и околоплодные воды из полости носа.
Большое значение при этом имеют тактильные и температурные раздражители, повышающие активность центрального дыхательного механизма, в ходе движения плода по родовым путям.