

Ответы на экзамен по микробиологии 2022 года
.pdf3.Лиазы – участвуют в реакциях негидролитического расщепления с образованием двойных связей или присоединение по двойным связям (гликолиз, брожение, ЦТК).
4.Изомеразы – превращение органических соединений в их изомеры (органические кислоты, аминокислоты, углеводы).
5.Лигазы (синтетазы) – соединение двух молекул с образованием новой химической связи (реакции биосинтеза белков, жиров, углеводов).
6.Гидролазы – катализируют реакции расщепления белков, жиров и углеводов с участием воды (расщепление веществ на более простые соединения). К гидролазам относятся бета-лактамазы (катализируют расщепление бета-лактамного кольца у пенициллинов, цефалоспоринов, монобактамов, карбапенемов) и эстеразы (воздействуют на эритромицин и другие антибиотики близкие к нему по структуре). Многие гидролазы являются ферментами агрессии:
o гиалуронидаза – расщепляет гиалуроновую кислоты соединительной ткани; o нейраминидаза – нейраминовую кислоту слизистых оболочек;
o коллагеназа – коллаген мышечных волокон;
o лецитиназа – лецитин мембран эритроцитов и мышечных волокон; o протеиназа – иммуноглобулины.
Классификация ферментов по месту действия:
1.Эндоферменты (функционируют внутри клетки): оксидоредуктазы, трансферазы, лиазы, изомеразы, лигазы.
2.Экзоферменты (выделяются в окружающую среду): гидролазы.
Классификация ферментов по генетическому контролю:
1.Конститутивные – синтезируются в течение всей жизни микроорганизма.
2.Индуцибельные – синтезируются только в присутствии соответствующего субстрата. Например, лактаза (бета-галактозидаза) синтезируется только при наличии в среде лактозы.
3.Репрессибельные – синтез ферментов подавляется избыточным накоплением продуктов реакции.
Ферментативный спектр является таксономическим признаком, характерным для семейства, рода и в некоторых случаях для видов. Поэтому определением спектра ферментативной активности пользуются при установлении таксономического положения бактерий. Наличие экзоферментов можно определить при помощи дифференциально-диагностических сред. Для идентификации бактерий разработаны специальные тест-системы, состоящие из набора дифференциально-диагностических сред.
13. Спирохеты, их морфология и биологические свойства. Патогенные для человека виды.
Спирохеты – тонкие длинные извитые грамотрицательные бактерии. Они состоят из наружной мембранной клеточной стенки, которая окружает цитоплазматический цилиндр. Поверх наружной мембраны располагается прозрачный чехол гликозаминогликановой природы. Под наружной мембраной располагаются фибриллы, закручивающиеся вокруг цитоплазматического цилиндра, придавая бактериям винтообразную форму. Фибриллы прикреплены к концам клетки и направлены навстречу друг другу. Число и расположение
31
фибрилл варьируют у разных видов. Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение. При этом спирохеты образуют петли, завитки, изгибы, которые названы вторичными завитками. Спирохеты плохо воспринимают красители. Обычно их окрашивают по Романовскому-Гимзе или серебрением. В живом виде спирохеты исследуют с помощью фазово-контрастной или темнопольной микроскопии.
Спирохеты представлены тремя родами, патогенными для человека: Treponema, Borrelia, Leptospira.
Трепонемы имеют вид тонких штопорообразно закрученных нитей с 8-12 равномерными мелкими завитками. Вокруг протопласта трепонем расположены 3-4 фибриллы (жгутики). В цитоплазме имеются цитоплазматические филаменты. Патогенными представителями являются T. pallidum – возбудитель сифилиса, T. pertenue – возбудитель тропической болезни
– фрамбезии. Имеются и сапрофиты – обитатели полости рта человека, ила водоёмов.
Боррелии, в отличие от трепонем, более длинные, имеют по 3-8 крупных завитков и 7-20 фибрилл. К ним относятся возбудитель возвратного тифа (B. recurrentis) и возбудители болезни Лайма (B. burgdorferi) и других заболеваний.
Лептоспиры имеют завитки неглубокие и частые в виде закрученной верёвки. Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв S или С; имеют две осевые фибриллы. Патогенный представитель L. interrogans вызывает лептоспироз при попадании в организм с водой или пищей, приводя к кровоизлияниям и желтухе.
14. Грибы, их морфология и особенности биологии. Принципы систематики.
Грибы – многоклеточные или одноклеточные гетеротрофные эукариотические микроорганизмы (эукариоты) с клеточной стенкой (отсутствует она лишь у зооспор и вегетативных клеток некоторых примитивных грибов). Они имеют хорошо оформленное ядро, митохондрии и вакуоли. Грибная клетка содержит одно или несколько ядер. Клеточная стенка грибов состоит на 80-90% из полисахаридов (45% приходится на хитин) и 10-20% приходится на белки и липиды. Соотношение химических ингредиентов клеточной стенки грибов определяет их вирулентность. Строение клеточной стенки определяет выбор антимикотических средств.
Все грибы являются гетеротрофными организмами. Минеральные вещества гриб способен усваивать из окружающей среды, однако органические он должен получать в готовом виде. Грибы не способны усваивать крупные частички пищи, поэтому всасывают исключительно жидкие вещества через всю поверхность тела, при этом огромная площадь поверхности мицелия оказывается весьма выгодной.
Выделяют два типа роста грибов – дрожжевой и мицелиальный (гифальный).
Основа вегетативного тела плесневых грибов – мицелий, состоящий из ветвящихся гиф. У простейших грибов – зачаточный мицелий, представленный единичной клеткой с
32
тонкими гифообразными отростками. У низших грибов (Mucor) – мицелий одноклеточный, обильно ветвящийся, со множеством ядер. У высших грибов (Penicillium, Aspergillus) мицелий разделён перегородками (септами) на отдельные клетки. Гифы плотно переплетаясь образуют ложную ткань. Мицелий обычно дифференцируется на две функционально различные части: субстратный (вегетативный), служащий для прикрепления к субстрату (питательная среда, ткани человека, животных), поглощения и транспортировки воды и растворённых в ней веществ. Его функция – питание микроорганизма. И воздушный (репродуктивный), поднимающийся над субстратом и образующий органы размножения.
Более высоко организованными считаются дрожжи и дрожжеподобные грибы. У них мицелий отсутствует, тело в вегетативном состоянии представляет округлую клетку. Дрожжеподобные грибы образуют псевдомицелий. Он отличается от мицелия плесневых грибов тем, что не имеет общей оболочки и перегородок, а состоит из длинных клеток, образующихся путём последовательного бокового или концевого почкования.
Важнейшей особенностью грибов является диморфизм. Диморфизм – способность расти в двух морфологических формах: дрожжевой или мицелиальной. Трансформация гриба затрудняет диагностику.
Инфекционными агентами грибов считаются споры и конидии (бесполые споры). Заболевания, вызванные грибами, обозначают термином микозы.
Грибам присуще 3 типа размножения: вегетативное, бесполое, половое.
Вегетативное размножение. На начальном этапе инфекционного процесса, когда необходимо быстро колонизировать поражаемую ткань, гриб размножается почкованием или частями мицелия. Наблюдается фрагментация гиф на отдельные клетки, каждая из которых даёт начало новому организму. При снижении влажности среды, исчерпании питательного субстрата (питательная среда, ткани человека и животных), многие грибы начинает «искать» новый источник питательных веществ и приступают к бесполому размножению.
Бесполое размножение осуществляется с помощью экзо- и эндоспор. Для облегчения распространения споры выносятся над субстратом, формируя репродуктивный мицелий. Большинство патогенных грибов размножаются экзоспорами или конидиями. Конидии образуются по бокам или на концах гиф. Морфологические особенности конидий лежат в основе классификации грибов.
Например, у мукора споры формируются эндогенно в специализированных органах – спорангиях, расположенных на вершине спорангиеносца. При разрыве спорангия споры высвобождаются, попадают в благоприятные условия и прорастают. У аспергилла конидиеносцы несептированные, в форме лейки, у пеницилла – септированные в виде кисти рук.
33
При неблагоприятных условиях гриб формирует хламидоспоры. Хламидоспора – гифальная клетка увеличенных размеров, покрытая плотной оболочкой. Она необходима для сохранения гриба. Образование хламидоспор затрудняет лечение микозов.
Половое размножение. Даёт преимущества грибам: возникают неидентичные особи, способные лучше адаптироваться к условиям обитания и осваивать новые экологические ниши.
15. Морфология, ультраструктура и химический состав вирусов. Принципы классификации.
Вирусы – своеобразная форма жизни, неклеточные биологические объекты, способные репродуцироваться только внутри живых клеток организма (в цитоплазме или ядре клетки), относятся к царству Vira.
Вирусы – автономные генетические структуры. Отличаются дизъюнктивным (разобщённым) способом репродукции (размножения), это означает, что в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом.
Вирусы располагаются на границе жизни. С неживой природой их роднит:
форма кристалла,
отсутствие метаболизма вне клетки хозяина.
Сходство вирусов с живой природой:
самовоспроизводство и наследственность,
изменчивость,
тропность (способность избирательно поражать определённые клетки).
Отличия вирусов от прокариотической клетки:
Вирусы мелких размеров, измеряются в нанометрах, проходят через бактериальные фильтры.
Наличие одного типа нуклеиновой кислоты (ДНК или РНК).
Отсутствие клеточного строения.
Не способны расти и бинарно делиться.
Вирусы – облигатные внутриклеточные паразиты на генетическом уровне.
Не имеют белоксинтезирующих и энергетических систем.
Отличаются дизъюнктивным способом репродукции.
Принципы классификации вирусов:
1.Тип нуклеиновой кислоты:
a.дезоксирибовирусы (ДНК-содержащие),
b.рибовирусы (РНК-содержащие).
2.Форма вириона и тип симметрии.
3.Наличие суперкапсидной оболочки.
4.Особенности репродукции (жизненный цикл вируса).
5.Антигенные свойства.
6.Механизм передачи возбудителей.
34
7.Круг восприимчивых хозяев.
8.Патогенность вируса.
Вирусы существуют в двух морфологических вариантах:
внутриклеточная форма – вирус,
внеклеточная форма – вирион.
Вирион – конечная стадия развития вируса. Полноценная вирусная частица, находящаяся вне живой клетки.
Формы вирионов:
палочковидная (вирус табачной мозаики),
пулевидная (вирус бешенства),
нитевидная (филовирусы – вирус Эбола),
сферическая (вирус гриппа, кори, ВИЧ),
кубоидальная – форма икосаэдра (аденовирусы, вирус полиомиелита, гепатитов А и Е),
сперматозоидная (бактериофаги).
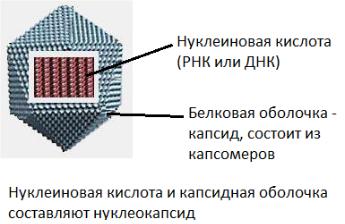
Структура вирусов
Просто устроенные вирионы
Функции капсида:
защитная;
скелетная (придаёт прочность, жёсткость конструкции);
адгезивная (избирательно взаимодействует с рецепторами клеток);
антигенная;
35
иммуногенная.
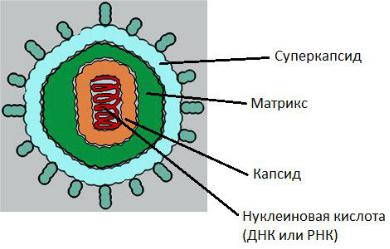
Сложно устроенные вирионы
Нуклеокапсид сложно организованных вирионов покрыт внешней оболочкой – суперкапсидом.
Суперкапсидная оболочка – биологическая мембрана клеточного происхождения; состоит из липидного бинарного слоя со встроенными белками (представлена мембранами ЦПМ, ядра, ЭПС клетки хозяина).
Матрикс – белковый компонент, локализован между суперкапсидом и капсидом.
Геном вирусов представлен различными видами нуклеиновых кислот: двунитевыми, однонитевыми, линейными, кольцевыми, фрагментированными.
Среди РНК-содержащих вирусов различают:
Вирусы с положительным геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК).
Вирусы с отрицательным геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Белки вирусов:
Структурные белки капсида – простые белки, обладающие способностью к самосборке. Они складываются в геометрически правильные структуры, в которых различают несколько типов симметрии: спиральный, кубический, смешанный.
Функциональные белки – ферменты.
1.Ферменты репликации и транскрипции: ДНК-полимераза и РНК-полимераза.
2.Вирус-индуцированные ферменты, участвуют в адгезии, проникновении вирусной нуклеиновой кислоты в клетку хозяина и выходе из неё образовавшихся вирионов: нейраминидаза, лизоцим, АТФ-аза.
Липиды и углеводы есть в суперкапсидной оболочке сложноустроенных вирусов.
16. Взаимодействие вируса с клеткой. Фазы жизненного цикла. Понятие о персистенции вирусов и персистентных инфекциях.
Взаимодействие вируса с клеткой хозяина (типы взаимодействия):
1. Продуктивная вирусная инфекция (происходит репродукция вируса, а клетки погибают).
36
2.Абортивная вирусная инфекция (репродукции вируса не происходит, а клетка восстанавливает нарушенную функцию).
3.Латентная вирусная инфекция (идёт репродукция вируса, а клетка сохраняет свою функциональную активность).
4.Вирус-индуцированная трансформация (клетка, инфицированная вирусом, приобретает новые, ранее не присущие ей свойства).
5.Интегративная инфекция – интегрированная (встроенная) в клеточный геном вирусная ДНК называется провирусом. В этом случае образование дочерних вирусных частиц не происходит, вирусные и клеточные гены передаются по наследству дочерним клеткам при делении.
Жизненный цикл вируса (стадии):
1.Адсорбция вириона на клетке. Адсорбция осуществляется за счёт специфического взаимодействия антирецептора вириона с комплементарными рецепторами мембраны клетки хозяина.
2.Проникновение в цитоплазму (пенетрация).
a.У просто устроенных вирусов осуществляется путём эндоцитоза – образуется эндоцитарная вакуоль, в которую заключается вирион.
b.У сложно устроенных вирусов проникновение в цитоплазму осуществляется путём слияния клеточной мембраны с суперкапсидной оболочкой.
3.Депротеинизация (раздевание). Осуществляется клеточными ферментами, разрушающими капсид. При этом вирусный геном освобождается от белков. Вирус на время как бы исчезает (эта стадия называется эклипс).
4.Синтез вирусных компонентов. На этой стадии синтезируются вирусные белки и нуклеиновые кислоты.
5.Сборка вирионов. Может происходить в цитоплазме, в ядре, на внутренней поверхности клеточной мембраны или на мембране ядра клетки хозяина.
6.Выход вирионов из клетки.
a.Сформировавшиеся безоболочечные (просто устроенные) вирионы выходят из клетки путём взрыва. В результате наблюдается деструкция клетки – это литический тип репродукции (пример, вирус полиомиелита).
b.Сформировавшиеся оболочечные (сложно устроенные) вирионы выходят из клетки методом почкования. В результате не наблюдается деструкция клетки – это нелитический тип репродукции (вирус гепатита В).
Вирусная персистенция – сохранение вируса в функционально активном состоянии в клетках организма или культур ткани за пределами тех сроков, которые характерны для острой инфекции. Соответственно, инфекции, обусловленные феноменом вирусной персистенции, называют персистентными вирусными инфекциями. Как правило, они протекают при менее выраженных по сравнению с острой инфекцией клинических проявлениях, либо вовсе без них.
Выделяют три группы персистентных вирусных инфекций: хронические, латентные и медленные.
37
Хронические инфекции отличаются от двух других тем, что присутствие вируса определяется относительно просто с помощью лабораторных методов; к их числу относятся, например, хронические формы вирусного гепатита В. При латентных инфекциях, типичных, в частности, для герпесвирусов, возбудитель маскирован в тканях, и его выявление возможно при обострениях. При медленных инфекциях вирусная персистенция приходится на чрезвычайно длительный (несколько лет) инкубационный период, предшествующий медленно развивающемуся заболеванию.
17. Принципы и методы лабораторной диагностики вирусных инфекций. Методы культивирования вирусов.
Для лабораторной диагностики вирусных инфекций используются различные методы.
Световая микроскопия позволяет обнаружить характерные вирусные включения, а электронная микроскопия – сами вирионы, и по особенностям их строения диагностировать соответствующую инфекцию (например, ротавирусную).
Для выделения вирусов используют заражение лабораторных животных, куриных эмбрионов или культуры тканей.
Первичную идентификацию выделенного вируса до уровня семейства можно провести с помощью:
определения типа нуклеиновой кислоты,
особенностей строения (электронная микроскопия),
размера вириона (фильтрование через мембранные фильтры с порами),
наличия суперкапсидной оболочки (проба с эфиром),
гемагглютининов (реакция гемагглютинации),
типа симметрии нуклеокапсида (электронная микроскопия).
Существенное значение для идентификации вирусов (до рода, вида, внутри вида) имеет также изучение их антигенного строения, которое проводится в реакции вирусонейтрализации с соответствующими иммунными сыворотками. Сущность этой реакции состоит в том, что после обработки гомологичными антителами вирус утрачивает свою биологическую активность (нейтрализуется) и клетка хозяина развивается так же, как и неинфицированная вирусом. Об этом судят по отсутствию цитопатического действия, цветной пробе, результатам реакции торможения гемагглютинации (РТГА), отсутствию изменений при заражении куриных эмбрионов, выживаемости чувствительных животных.
Методы иммунодиагностики. Они реализуются в самых разнообразных реакциях иммунитета:
радиоизотопный иммунный анализ (РИА),
иммуноферментный анализ (ИФА),
реакция иммунофлюоресценции (РИФ),
реакция связывания комплемента (РСК),
реакция пассивной гемагглютинации (РПГА),
реакции торможения гемагглютинации (РТГА) и другие.
При использовании методов серодиагностики обязательным является исследование парных сывороток. При этом четырёхкратное нарастание титра антител во второй сыворотке в
38
большинстве случаев служит показателем протекающей или свежеперенесённой инфекции. При исследовании одной сыворотки, взятой в острой стадии болезни, диагностическое значение имеет обнаружение антител класса Ig М, свидетельствующее об острой инфекции.
С помощью молекулярно-генетических методов (ПЦР) выявляют персистирующие вирусы, находящиеся в клиническом материале, с трудом обнаруживаемые или не обнаруживаемые другими методами.
Для культивирования вирусов используют культуры клеток, куриные эмбрионы и чувствительных лабораторных животных. Эти же методы используют и для культивирования риккетсий и хламидий – облигатных внутриклеточных бактерий, которые не растут на искусственных питательных средах.
Культуры клеток:
Первичные клеточные культуры. Получают методом трипсинизации эмбриональных тканей, выдерживают не более 5-10 пассажей после выделения из тканей.
Полуперевиваемые культуры клеток. Получают из диплоидных клеток эмбриона человека (сохраняют при пассажах диплоидный набор хромосом), выдерживают 50-60 пассажей.
Перевиваемые культуры клеток. Получают из опухолевых клеток. Сохраняют жизнеспособность в процессе неограниченного числа пересевов (десятки лет).
Феномены репродукции вирусов:
Цитопатогенное действие вирусов (ЦПД).
Образование внутриклеточных включений (скопления вирусов или их компонентов в цитоплазме или ядре клетки).
Образование «бляшек».
Реакция гемадсорбции и гемагглютинации.
«Цветная» реакция.
18.Вирусы бактерий – фаги. Взаимодействие фага с бактериальной клеткой. Умеренные и вирулентные бактериофаги. Профаг. Лизогения. Фаговая конверсия. Применение фагов в биотехнологии, микробиологии и медицине.
Бактериофаги – вирусы бактерий. Открыты в 1917 году французским микробиологом Д’Эррелем.
Бактериофаги специфичны, то есть лизируют определенные виды бактерий. Отсюда их названия: дизентерийный бактериофаг, стафилококковый бактериофаг.
Структура бактериофагов
Размеры бактериофагов колеблются от 20 нм до 200 нм. Как все вирусы, содержат ДНК, или РНК, и белковый капсид. Чаще всего встречаются и лучше изучены бактериофаги, имеющие форму сперматозоида или головастика. Они имеют:
двунитевую ДНК;
головку кубического типа симметрии;
воротничок;
39
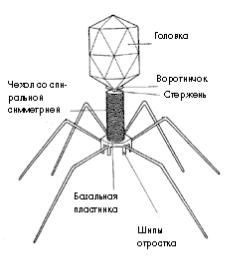
хвостовой отдел спирального типа симметрии, снаружи сократительный чехол, заканчивающийся базальной пластинкой;
от базальной пластинки отходят белковые нити – фибриллы.
Взаимодействие бактериофага с бактериальной клеткой
Продуктивная инфекция. Вирулентные (вегетативные) фаги репродуцируются в бактериальной клетке и вызывают её лизис.
Интегрированная инфекция. Умеренный фаг (профаг) встраивается в клеточный геном, вступает в своеобразный симбиоз с микробной клеткой. Лизогения – встраивание умеренного фага (профага) в клеточный геном.
Умеренные бактериофаги (особенности):
В неактивной стадии профага бактериальная клетка сохраняет жизнеспособность и всё потомство заражённой клетки содержит профаг.
Культура, заражённая умеренным фагом, называется лизогенной.
Умеренный фаг может активироваться в результате воздействия физических и химических факторов.
В результате активации профага происходит синтез новых фаговых частиц (взрыв репродукции) и гибель бактериальной клетки.
Фаговая конверсия (лизогенная конверсия)
Геном профага может придавать бактерии новые свойства. Это явление называется фаговой конверсией.
Конвертироваться могут морфологические, культуральные, биохимические, антигенные, токсические и др. свойства бактерий.
Пример: наличие профага в дифтерийной палочке обусловливает её способность продуцировать дифтерийный экзотоксин.
Практическое применение бактериофагов
Диагностика: определение вида микроорганизма и источника инфекции.
Эпидемиологическая оценка количества патогенных бактерий по наличию специфических бактериофагов.
Получение вакцинных штаммов с использованием умеренных бактериофагов.
Профилактика и лечение заболеваний, лечение дисбактериозов.
40