

Нейрохирургия
.pdfморфологические изменения - отражение цитологических процессов, сопровождающих усиленный синтез белка.
Участок аксона дистальнее места перерезки отмирает. В течение нескольких дней этот участок и все синаптические окончания аксона подвергаются разрушению. Миелиновая оболочка аксона тоже дегенерирует, ее фрагменты захватываются фагоцитами. Однако клетки нейроглии, образующие миелин, не погибают. Эта последовательность явлений получила название уоллеровской дегенерации.
Если поврежденный аксон обеспечивал единственный или основной синаптический вход к нервной или к эффекторной клетке, то постсинаптическая клетка может подвергнуться дегенерации и погибнуть. Хорошо известный пример - атрофия волокон скелетной мышцы после нарушения их иннервации мотонейронами. Регенерация аксона После дегенерации поврежденного аксона у многих нейронов может отрастать
новый аксон. На конце проксимального отрезка аксон начинает ветвиться [спрутинг (sprouting) - разрастание]. В ПНС новообразованные ветви растут вдоль исходного пути погибшего нерва, если, конечно, этот путь доступен. В период уоллеровской дегенерации шванновские клетки дистальной части нерва не только
выживают, но и пролиферируют, выстраиваясь рядами там, где проходил погибший нерв. «Конусы роста» регенерирующего аксона прокладывают свои пути между рядами шванновских клеток и в конечном итоге могут достигать своих мишеней, реиннервируя их. Затем аксоны ремиелинизируются шванновскими клетками. Скорость регенерации ограничивается скоростью медленного аксонного транспорта, т.е. примерно 1 мм/сут.
Регенерация аксонов в ЦНС имеет некоторые отличия: клетки олигодендроглии не могут наметить путь для роста ветвей аксона, поскольку в ЦПС каждый олигодендроцит миелинизирует множество аксонов (в отличие от шванновских клеток в ПНС, каждая из которых снабжает миелином только один аксон).
Важно отметить, что химические сигналы по-разному действуют на регенерационные процессы в ЦНС и ПНС. Дополнительное препятствие регенерации аксонов в ЦНС - глиальные рубцы, формируемые астроцитами. Синаптический спрутинг, обеспечивающий «реусиление» существующих нейрональных токов и образование новых полисинаптических связей, обусловливает пластичность нейрональной ткани и формирует механизмы, участвующие в восстановлении нарушенных неврологических функций. Трофические факторы Важную роль в развитии ишемического повреждения ткани мозга играет уровень его трофического обеспечения.
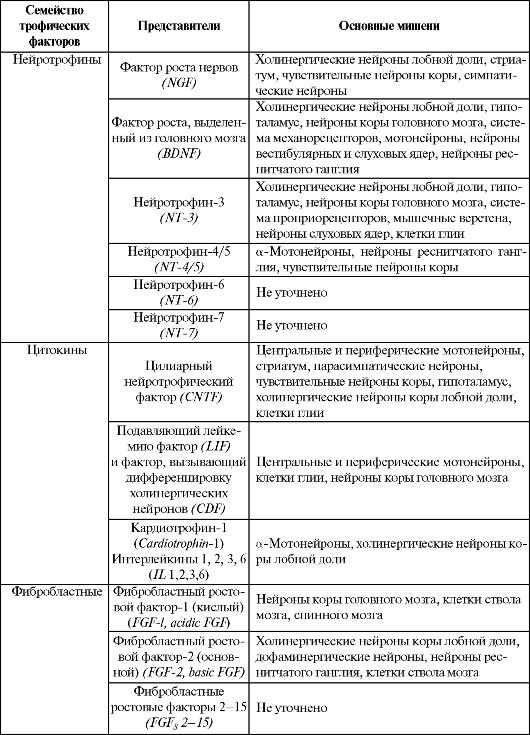
Нейротрофические свойства присущи многим протеинам, в том числе и структурным белкам (например, S1OOβ). В то же время максимально их реализуют факторы роста, которые представляют разнородную группу трофических факторов, состоящую по меньшей мере из 7 семейств, - нейротрофины, цитокины, фибробластные факторы роста, инсулинзависимые факторы роста, семейство трансформирующего фактора роста 31(TGF-J3I), эпидермальные факторы роста и другие, в том числе ростовой протеин 6 (GAP-6)4, тромбоцитзависимый фактор роста, гепаринсвязанный нейротрофический фактор, эритропоэтин, макрофагальный колониестимулирующий фактор и др. (табл. 1.2).
Наиболее сильное трофическое влияние на все основные процессы жизнедеятельности нейронов оказывают нейротрофины - регуляторные белки нервной ткани, которые синтезируются в ее клетках (нейронах и глии). Они действуют локально - в месте высвобождения и особенно интенсивно индуцируют ветвление дендритов и рост аксонов в направлении клеток-мишеней.
К настоящему времени наиболее изучены три нейротрофина, близких друг другу по структуре: фактор роста нервов (NGF), фактор роста, выделенный из головного мозга (BDNF), и нейротрофин-3 (NT-3).
Таблица 1.2. Современная классификация нейротрофических факторов

В
развивающемся организме они синтезируются клеткой-мишенью (например, мышечным веретеном), диффундируют по направлению к нейрону, связываются с молекулами рецепторов на его поверхности.
Связанные с рецепторами факторы роста захватываются нейронами (т.е. подвергаются эндоцитозу) и транспортируются ретроградно в сому. Там они могут воздействовать непосредственно на ядро, изменяя образование ферментов, ответственных за синтез нейромедиаторов и рост аксонов. Различают две формы рецепторов к факторам роста - низкоаффинные рецепторы и высокоаффинные рецепторы тирозинкиназы, с которыми связывается большинство трофических факторов.
В результате аксон достигает клетки-мишени, устанавливая с ней синаптический контакт. Факторы роста поддерживают жизнь нейронов, которые в их отсутствие не могут существовать.
Трофическая дизрегуляция является одной из универсальных составляющих патогенеза повреждения нервной системы. При лишении трофической поддержки зрелых клеток развивается биохимическая и функциональная дедифференциация нейронов с изменением свойств иннервируемых тканей. Трофическая дизрегуляция сказывается на состоянии макромолекул, принимающих участие в мембранном электрогенезе, активном ионном транспорте, синаптической передаче (ферменты синтеза медиаторов, постсинаптические рецепторы) и эффекторной функции (мышечный миозин). Ансамбли дедифференцированных центральных нейронов создают очаги патологически усиленного возбуждения, запускающего патобиохимичсские каскады, которые ведут к гибели нейронов по механизмам некроза и апоптоза. Напротив, при достаточном уровне трофического обеспечения регресс неврологического дефицита после ишемического повреждения мозга часто наблюдается даже при оставшемся морфологическом дефекте, изначально его вызвавшем, что свидетельствует о высокой приспособляемости мозговой функции. Установлено, что в развитии недостаточности трофического обеспечения принимают участие изменения калиевого и кальциевого гомеостаза, избыточный синтез оксида азота, который блокирует фермент тирозинкиназу, входящий в активный центр трофических факторов, дисбаланс цитокинов. Одним из предполагаемых механизмов является аутоиммунная агрессия против собственных нейротрофинов и структурных нейроспецифических белков, обладающих трофическими свойствами, которая становится возможной в результате нарушения защитной функции гематоэнцефалического барьера.
ГЛАВА 2. КРАТКИЕ СВЕДЕНИЯ ПО АНАТОМИИ НЕРВНОЙ СИСТЕМЫ
2.1. Спинной мозг
Спинной мозг (medulla spinalis) расположен в позвоночном канале. На уровне I шейного позвонка и затылочной кости спинной мозг переходит в продолговатый, а книзу тянется до уровня I-II поясничных позвонков (рис. 2.1). По длиннику спинного мозга выделяют шейное и пояснично-крестцовое утолщения. Спинной мозг состоит
из 31 - 32 сегментов: 8 шейных (CI - CIV), 12 грудных (ТhI-ТhXII), 5 поясничных (LI-LV), 5 крестцовых (SI-SV) и 1-2 рудиментарных копчиковых (СоI - СоII). Шейное утолщение
соответствует сегментам СV-ТhI и обеспечивает иннервацию верхних конечностей. Пояснично-крестцовое утолщение соответствует сегментам LI-SI-II и иннервирует нижние конечности. Ниже поясничного утолщения спинной мозг истончается,
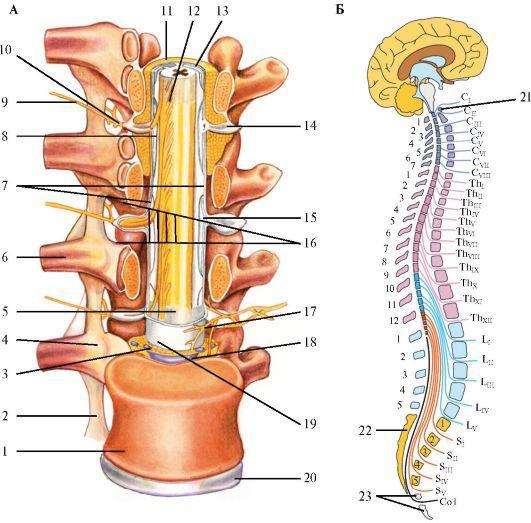
образуя мозговой конус, который заканчивается концевой нитью, доходящей до копчиковых позвонков.
Сегментом называется участок спинного мозга, которому принадлежит одна пара спинномозговых корешков (передних и задних). Передний корешок спинного мозга содержит двигательные волокна, задний корешок - чувствительные волокна.
Соединяясь в области межпозвонкового узла, они образуют смешанный спинномозговой нерв.
Спинной мозг несколько короче позвоночного канала. В связи с этим в верхних отделах спинного мозга его корешки идут горизонтально. Затем, начиная с грудного отдела, они несколько спускаются книзу, прежде чем выйти из соответствующих межпозвоночных отверстий. В нижних отделах корешки идут прямо вниз, формируя конский хвост.
На поверхности спинного мозга видны передняя срединная щель, задняя срединная борозда, симметрично расположенные передние и задние латеральные борозды. Между передней срединной щелью и передней латеральной бороздой располагается передний канатик (funiculus anterior),между передней и задней латеральными бороздами - боковой канатик (funiculus lateralis), между задней латеральной бороздой и задней срединной бороздой - задний канатик (funiculus posterior), который в шейной части спинного мозга делится неглубокой промежу-
Рис.
2.1. Строение спинного мозга. А - Расположение спинного мозга в спинномозговом канале. C - Шейные спинномозговые нервы (СI-СVII, синие); Th - Грудные спинномозговые нервы (ThI-ThXII, фиолетовые); L - Поясничные спинномозговые нервы (LI-LV, бирюзовые); S - Крестцовые спинномозговые нервы (SI-SV, светлозеленые); Co - Копчиковые спинномозговые нервы (Со, серые); 1 - тело позвонка; 2 -
межпоперечная связка; 3 - эпидуральные вены; 4 - реберно-позвоночный сустав; 5 - паутинная оболочка; 6 - ребро; 7 - зубчатая связка; 8 - вентральный корешок; 9 - спинномозговой нерв; 10 - задняя ветвь (кожа и мышцы спины); 11 - эпидуральное пространство; 12 - мягкая мозговая оболочка; 13 - субарахноидальное пространство; 14 - спинальный ганглий; 15 - корешковый рукав; 16 - межпозвонковое отверстие; 17 - менингеальная ветвь; 18 - задняя продольная связка; 19 - твердая мозговая оболочка; 20 - межпозвоночный диск, фиброзное кольцо; 21 - атлант; 22 - крестец; 23 - копчик. Б - Сегменты спинного мозга и спинномозговые нервы
точной бороздой на тонкий пучок (fasciculus gracilis), прилежащий к задней срединной борозде и расположенный кнаружи от него клиновидный пучок(fasciculus cuneatus). Канатики содержат проводящие пути, из передней латеральной борозды выходят передние корешки, в области задней боковой борозды в спинной мозг входят задние корешки.
На поперечном срезе (см. рис. 2.2) в спинном мозге выделяются серое вещество, расположенное в центральных отделах спинного мозга, и белое вещество, лежащее на его периферии. Серое вещество на поперечном срезе напоминает по форме бабочку с раскрытыми крыльями или букву «Н». В сером веществе спинного мозга выделяют более массивные, широкие и короткие передние рога и более тонкие, удлиненные задние рога. В грудных отделах выявляется боковой рог, который меньше выражен в поясничном и шейном отделах спинного мозга. Правая и левая половины спинного мозга симметричны и соединены спайками из серого и белого вещества. Кпереди от центрального канала находится передняя
серая спайка (comissura grisea anterior), далее передняя белая спайка (comissura alba anterior); кзади от центрального канала последовательно располагаются задняя серая спайка (comissura grisea posterior) и задняя белая спайка (comissura alba posterior).
В передних рогах спинного мозга находятся крупные двигательные нейроны, аксоны которых идут в передние корешки и иннервируют поперечнополосатую мускулатуру шеи, туловища и конечностей. Первичные чувствительные клетки располагаются в спинномозговых (межпозвоночных) узлах, представляющих собой утолщения задних корешков. Такая нервная клетка имеет один отросток, который, отходя от нее, делится на две ветви. Одна из них идет на периферию, где получает раздражение от рецеп-
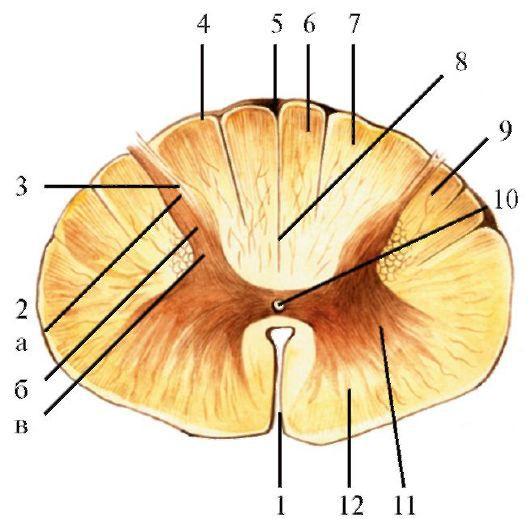
Рис.
2.2. Поперечный срез спинного мозга (схема).
1 - передняя срединная щель; 2 - задний рог: а - верхушка; б - головка; в - шейка; 3 - студенистое вещество; 4 - задний канатик; 5 - задняя срединная борозда; 6 - тонкий пучок; 7 - клиновидный пучок; 8 - задняя срединная перегородка; 9 - боковой канатик; 10 - центральный канал; 11 - передний рог; 12 - передний канатик торов кожи, мышц, сухожилий или внутренних органов, а по другой ветви эти импульсы передаются в спинной мозг. В зависимости от вида раздражения и
проводящего пути, по которому оно передается, афферентные волокна, входящие в спинной мозг через задний корешок, могут оканчиваться на клетках задних рогов либо непосредственно проходят в белое вещество спинного мозга.
Таким образом, клетки передних рогов осуществляют двигательные функции, клетки задних рогов - чувствительные, в боковых рогах локализуются сегментарные вегетативные клетки. На уровне сегментов СVIII-LIII в боковых рогах располагаются клетки симпатической нервной системы, на уровне сегментов SII-SIV локализован спинальный парасимпатический центр, обеспечивающий регуляцию функции тазовых органов.
Белое вещество спинного мозга состоит из волокон восходящих и нисходящих проводящих путей (рис. 2.3), осуществляющих взаимосвязь как различных уровней спинного мозга между собой, так и всех вышележащих отделов ЦНС со спинным мозгом.
В передних канатиках спинного мозга расположены проводящие пути, участвующие в осуществлении двигательных функций:
1) передний корково-спинномозговой (пирамидный) путь
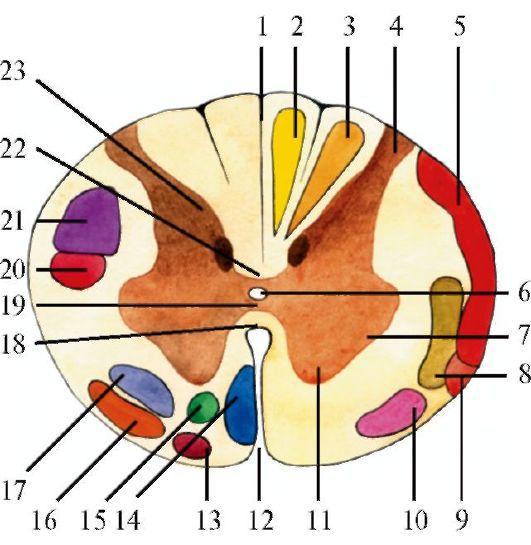
Рис.
2.3. Поперечный срез спинного мозга на уровне верхнегрудного отдела. Проводящие пути (схема). 1 - задняя срединная перегородка; 2 - тонкий пучок; 3 - клиновидный пучок; 4 - задний рог; 5 - задний спинно-мозжечковый путь; 6 - центральный канал; 7 - боковой рог; 8 - латеральный спинно-таламический путь; 9 - передний спинно-мозжечковый путь; 10 - передний спинно-таламический путь; 11 - передний рог; 12 - передняя срединная щель; 13 - оливоспинномозговой путь; 14 - передний корково-спинномозговой (пирамидный) путь; 15 - передний ретикулярноспинномозговой путь; 16 - преддверно-спинномозговой путь; 17 - бульборетикулярно-спинномозговой путь; 18 - передняя белая спайка; 19 - серая спайка; 20 - красноядерно-спинномозговой путь; 21 - латеральный корковоспинномозговой (пирамидный) путь; 22 - задняя белая спайка; 23 - грудной столб (столб Кларка)
(неперекрещенный), идущий в основном от двигательной области коры большого мозга и заканчивающийся на клетках передних рогов;
2)преддверно-спинномозговой (вестибулоспинальный) путь, идущий от латерального вестибулярного ядра той же стороны и заканчивающийся на клетках передних рогов;
3)покрышечно-спинномозговой путь, начинающийся в верхних холмиках четверохолмия противоположной стороны и заканчивающийся на клетках передних рогов;
4)передний ретикулярно-спинномозговой путь, идущий от клеток ретикулярной формации мозгового ствола той же стороны и заканчивающийся на клетках
переднего рога. Кроме того, вблизи от серого вещества проходят волокна, осуществляющие связь различных сегментов спинного мозга между собой.
Вбоковых канатиках спинного мозга располагаются как двигательные, так и чувствительные пути. К двигательным путям относятся:
1) латеральный корково-спинномозговой (пирамидный) путь (перекрещенный), идущий от двигательной области коры большого мозга и заканчивающийся на клетках передних рогов противоположной стороны; 2) руброспинальный путь, идущий от красного ядра и заканчивающийся на клетках передних рогов противоположной стороны;
3) ретикулярно-спинномозговые пути, идущие преимущественно от гигантоклеточного ядра ретикулярной формации противоположной стороны и заканчивающиеся на клетках передних рогов; 4) оливоспинномозговой путь, соединяющий нижние оливы с двигательным нейроном переднего рога.
К афферентным, восходящим проводникам бокового канатика относятся:
1) задний (дорсальный, неперекрещенный) спинно-мозжечковый путь, идущий от клеток заднего рога и заканчивающийся в коре мозжечка; 2) передний (перекрещенный) спинно-мозжечковый путь, идущий от клеток задних рогов и заканчивающийся в черве мозжечка;
3) латеральный спинно-таламический путь, идущий от клеток задних рогов и заканчивающийся в таламусе. Кроме того, в боковом канатике проходят спиннопокрышечный, спиноретикулярный, спинооливный пути.
Взадних канатиках спинного мозга располагаются афферентные тонкий и клиновидный пучки. Волокна, входящие в них, начинаются в межпозвонковых узлах и заканчиваются соответственно в ядрах тонкого и
клиновидного пучков, располагающихся в нижнем отделе продолговатого мозга. Таким образом, на уровне спинного мозга замыкаются рефлекторные дуги: возбуждение, приходящее по волокнам задних корешков, подвергается анализу и передается на клетки переднего рога для реализации ответа. Рефлекс может осуществляться при наличии трех звеньев:
• афферентной части, в которую входят рецепторы и проводящие пути, передающие раздражение в нервные центры;
• центральной части рефлекторной дуги, где формируется реакция;
• эффекторной части, реализующей реакцию через скелетную мускулатуру, гладкие мышцы и железы.
Спинной мозг, таким образом, является одним из первых этапов, на котором осуществляются анализ и синтез раздражений как из внутренних органов, так и от рецепторов кожных покровов, мускулатуры, слизистых оболочек. Наряду с этим спинной мозг передает приходящие афферентные импульсы в вышележащие отделы.
Сегментарный вегетативный аппарат спинного мозга участвует в реализации вегетативно-трофических влияний, симпатической иннервации мышц глаза, регуляции деятельности тазовых органов (мочеиспускания и дефекации). Периферические нервы спинного мозга Корешки спинного мозга имеют сегментарное распределение и разделяются на
задние (несущие чувствительные волокна) и передние (несущие двигательные волокна). Вблизи от спинного мозга по ходу заднего корешка расположен чувствительный межпозвонковый узел (спинномозговой, или спинальный), в котором находятся первые чувствительные нейроны всех систем поверхностной и глубокой чувствительности. За спинномозговым узлом на уровне каждого сегмента передний и задний корешки сливаются в общий ствол, образуя смешанный спинномозговой нерв (рис. 2.4).