

Строение и функции отдельных слоёв сетчатки
1. Пигментный (наружный) слой. Пигментные клетки (наружный слой). Содержат пигмент фусцин. Функции пигментного слоя:
-Трофическая функция для фоторецепторных клеток
- Депо витамина А
-Утилизация разрушенных наружных дисков фоторецепторов (1 диск в 40 мин)
- Меланин поглощает фотоны, не попавшие на фоторецептор, препятствуя рассеиванию света.
У альбиносов меланина нет → свет рассеивается во всех направлениях → острота зрения низкая (0.1-0.2). У ночных животных глаза в темноте светятся, т.к. пигментный слой не выражен, фотон света отражается и «гуляет» по сетчатке, возбуждая – приспособительная реакция для лучшего видения в темноте.
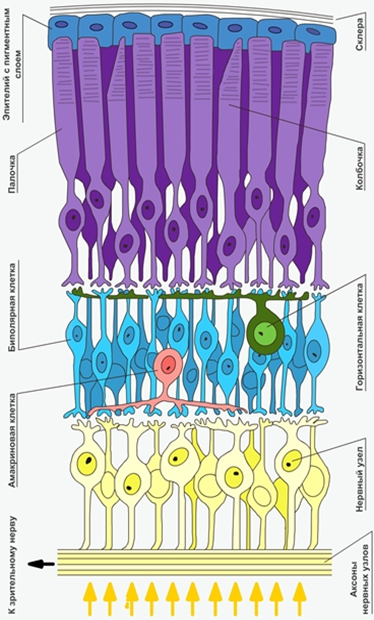
Рис. 8. Расположение клеток слоев сетчатки, относительно попадающих лучей света
2. Слой фоторецепторных клеток.
Ø колбочки: 3 млн.; в центре; длина 0,03 мм
Ø палочки: 100 млн.; на периферии; 0,06 мм.
Плотность упаковки:
колбочки: – 200 000 / мм2 максимально - в центральной ямке (высокая острота зрения)
палочки: в центре – нет, на периферии желтого пятна 60 000 - 5 000 /мм2
В центральной ямке (1 мм2) все слои смещены. Поэтому здесь только один слой – это слой колбочек. Здесь нет: палочек, биполярных, ганглиозных, амакриновых, горизонтальных клеток.
Рис.9. Электронная микрофотография центральной ямки: оранжевым и желтым цветом видны ядра клеток сетчатки.
Порог возбуждения фоторецепторов разный
Ø Палочки низкопороговые – для возбуждения достаточно 1 фотона → обеспечивают «сумеречное» зрение
Ø Колбочки высокопороговые – нужно 30-100 фотонов → дневное зрение
Причина: в наружных сегментах палочек больше дисков → больше мембран → больше ионных каналов.
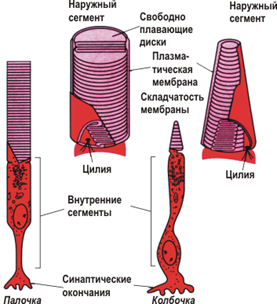
Рис. 10. Отличия в строении фоторецепторных клеток
Фоторецепция начинается с поглощения кванта света молекулой зрительного пигмента – родопсина. Локализован он в мембранах дисков наружного сегмента палочек и колбочек сетчатки, а сам сегмент не что иное, как сохранивший ножку видоизмененный жгутик: в палочках он имеет форму аккуратного цилиндра, а в колбочках – конуса. Внутри жгутика плазматическая мембрана образует многочисленные и регулярно расположенные диски, поставленные перпендикулярно ходу светового луча. Чем длиннее наружный сегмент, тем больше дисков должен пересечь луч и тем выше вероятность поглощения кванта.
Фоторецепция начинается с поглощения кванта света молекулой зрительного пигмента – родопсина. Локализован он в мембранах дисков наружного сегмента палочек и колбочек сетчатки, а сам сегмент не что иное, как сохранивший ножку видоизмененный жгутик: в палочках он имеет форму аккуратного цилиндра, а в колбочках – конуса. Внутри жгутика плазматическая мембрана образует многочисленные и регулярно расположенные диски, поставленные перпендикулярно ходу светового луча. Чем длиннее наружный сегмент, тем больше дисков должен пересечь луч и тем выше вероятность поглощения кванта.
Механизм восприятия световой энергии
Рецептор, воспринимающий фотон света – это молекула родопсина:
опсин (белок ) + ретиналь (альдегид витамина А)
Ø Родопсин палочек распадается при попадании на него волны любой длины, поэтому палочки не способны различать цвета (сумеречное зрение)
Особенность фоторецепторов- в покое (темноте) натриевые каналы удерживаются открытыми благодаря цГМФ → через мембрану наружного сегмента фоторецептора фиксируется постоянный входящий натриевый ток, что приводит к постоянному выделению порций медиатора глутамата и восприятию его биполярными клетками.
Свет → Образование транс-ретиналя → уменьшается количество цГМФ → закрываются Nа+ каналы → гиперполяризация → рецепторный потенциал → прекращается выделение медиатора (глутамата)
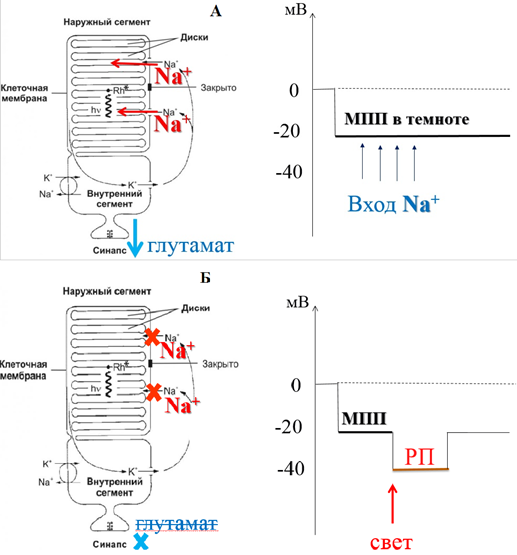
Рис. 11. Механизм генерации рецепторного потенциала в фоторецепторной клетке:
А – состояние фоторецептора в темноте; Б – состояние фоторецептора на свету.
Эффекта влияния фоторецептора на биполярную клетку хватает на 0,2 сек. Потом рецептор нужно сменить. Поэтому глаз должен сместиться – различают следующие типы движений глазных яблок:
- Саккады – быстро, скачкообразно (2-3 прыжка в сек)
- Дрейф – медленно
- Тремор (постоянный): 30-80 Гц
Рецепторный потенциал фоторецепторной клетки– гиперполяризационный, электротонический а значит, градуированный по силе: его амплитуда зависит от количества воспринятых фотонов (по закону силы).
Ø Для палочки:
1 фотон – min РП (-21 мВ)
60 фотонов – max РП (-40 мВ)
Ø Эта градуированность важна для кодирования силы света
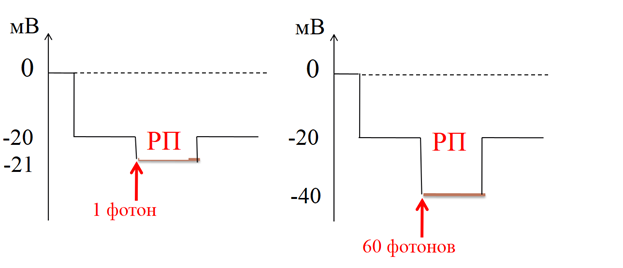
Рис. 12. Зависимость величины РП от силы светового раздражителя.