

7_Lektsia_Obmen_lipoproteinov
.pdfОБМЕН ЛИПОРОТЕИНОВ (ЛП)
ЛП – это сложные надмолекулярные комплексы, представляющие собой
водорастворимые функционально-активные белково-липидные молекулы.
Физиологическая роль ЛП
1.Используются клетками в качестве энергетических субстратов,
мембранного материала и предшественников в синтезе биологически важных
соединений.
2.ЛП играют важную роль в гомеостазе ХС: отдельные ЛП осуществляют
«захват» избыточного ХС с поверхности клеток периферических тканей и его
«обратный» транспорт в печень для окисления в желчные кислоты и выведения с желчью.
3. В составе ЛП транспортируются некоторые стероидные гормоны и жирорастворимые витамины.
Строение ЛП
ЛП имеют сферическую форму и состоят из липидов (прдставленных ФЛ, ХС,
ТАГ и ЭХС) и белкового компонента (апобелков).
Основную массу ЛП составляет его ядро, в котором находятся неполярные липиды: ТАГ и ЭХС.
Ядро окружено гидрофильным монослоем (оболочкой), образованным полярными липидами (ФЛ, ХС) и апобелками периферическими и стационарными. Причем ХС встраивается между жирнокислотными радикалами ФЛ.
Периферические апобелки (или динамические) представляют собой небольшие полипептиды, в состав которых входят до 5% углеводов: маноза,
глюкоза, фруктоза, галактоза, галактозамин и сиаловая кислота.
Всю толщу фосфолипидного монослоя пронизывает стационарный апобелок
(или белок-вектор, или интегральный белок). Химические связи между компонентами ЛП носят нековалентный характер.
периферический белок
ФЛ
ХС
интегральный
белок 
ТАГ
ЭХС
МОНОСЛОЙ |
ЯДРО |
Строение ЛП
Апобелки – это оболочки ЛП, нековалентно связанные с ФЛ и ХС, что необходимо, чтобы апобелки поддерживали структурную целостность ЛП,
обеспечивали и участвовали в процессах обмена между ЛП различных классов.
Они отвечают за взаимодействие ЛП с рецепторами. Апобелки формируют белок-липидные комплексы и определяют их специфическую структуру.
Апобелки классифицируются на стационарные и динамические.
Стационарные апобелки являются интегральными, т.е. пронизывают всю толщу фосфолипидного монослоя, и никогда не покидают ЛП. К ним относятся:
В-48 – кишечного происхождения;
В-100 – печеночного происхождения;
А-1 – кишечного и печеночного происхождения.
Динамические апобелки являются периферическими белками. Это непостоянные компоненты ЛП и они способны перемещаться между ЛП различных классов.
К ним относят: СII – печёночного происхождения, Е – кишечного
происхождения, а также синтезируемого в миоцитах, эпителии почек,
надпочечников, астроцитах нейроглии, моноцитах, макрофагах; СIII –
печёночного происхождения, Апо-АII – кишечного и печёночного
происхождения, АIV – печёночного и кишечного происхождения, БПЭХ – белок,
переносящий эфиры ХС (Апо-D).
Функции апобелков
1.Структурообразование.
Стационарные белки связывают липиды и формируют белок-липидные комплексы:
Например, В-48 – акцептируют ТАГ, В-100 – ТАГ и ЭХС, А-1 – ФЛ.
Динамические белки также участвуют в структурообразовании ЛП.
Например, А-IV – акцептирует ХС и транспортирует его к ЛПВП.
2.Векторная функция.
Стационарные белки обеспечивают направленный транспорт ЛП к клеткам-
мишеням, на которых находятся рецепторы к соответствующим белкам.
Динамический АПО-Е обеспечивает взаимодействие стационарных белков с рецепторами клеток-мишеней.
3.Являются кофакторами ферментов, участвующих в обмене ЛП.
СII – кофактор липопротеинлипазы (ЛПЛ), СIII – кофактор печеночной глицерол гидролазы, ее переносчик и ингибитор ЛПЛ, АII – кофактор α- и β-
лецитинхолестеролацилтрансферазы (ЛХАТ), Е – ингибитор ЛПЛ.
Классификации ЛП
Основаны на различиях их физико-химических свойств.
I. Классификация, основанная на плотности ЛП и их выделении в процессе
препаративного ультрацентрифугирования:
1.ХМ (хиломикроны)
2.ЛПОНП (ЛП очень низкой плотности)
3.ЛППП (ЛП промежуточной плотности)
4.ЛПНП (ЛП низкой плотности)
5.ЛПВП (ЛП высокой плотности).
II. Классификация, основанная на поведении ЛП в электрическом поле при электрофорезе на бумаге:

1.ХМ
2.ЛПОНП (пре β-ЛП)
3.ЛПНП (β-ЛП)
4.ЛПВП (α-ЛП)
Плотность ЛП определяется соотношением апопротеины/липиды; чем больше это соотношение, тем выше плотность.
Плотность при электорофорезе зависит от содержания АПО-протеинов и полярных липидов.
III. Классификация по составу в зависимости от соотношения компонентов:
1. ЛП, богатые ТАГ ХМ, ЛПОНП
2.ЛП, богатые Хс  ЛПНП
ЛПНП
3.ЛП, богатые ФЛ  ЛПВП
ЛПВП
Ферменты, участвующие в метаболизме ЛП
1.Липопротеинлипаза (ЛПЛ) гидролизует сложноэфирные связи в α-положении ТАГ (1 и 3 связи), входящих в структуру ХМ и ЛПОНТ. ТАГ расщепляется до ДАГ и ЖК, и β-МАГ. ЛПЛ локализуется на эндотелии капилляров. Гепарин вызывает высвобождение ЛПЛ во время приема пищи. Кофактором ЛПЛ является АПО-С II.
Место синтеза: жировая ткань, сердечная и скелетные мышцы, почки, легкие,
лактирующая молочная железа, селезенка.
2. Печёночная глицеролгидралаза (ПГГ) также гидролизует сложноэфирные связи в α-положении ТАГ (1 и 3 связи), входящих в структуру ЛППП, ЛПНП (при
малой доступности субстрата) до ДАГ, β-МАГ и ЖК.
Локализуется на эндотелии синусовых капилляров печени. ПГГ транспортируется по крови в комплексе с С-III, который одновременно является её кофактором.
3. Лецитинхолестеролацилтрансфераза (ЛХАТ) катализирует реакцию этерификации ХС, путём переноса остатка ВЖК из второго положения ФХ
(лецитин) ЛП на гидроксильную группу ХС с образованием ЭХС и лизолецитина:

ХС + ФХ(лецитин) |
ЛХАТ в крови |
ЭХС + лизолецетин |
|
Существует 2 изоформы ЛХАТ: α и β.
Α-ЛХАТ катализирует этерификацию ХС полиненасыщенных ЖК на кишечном пуле ЛПВП.
Β-ЛХАТ катализирует этерификацию ХС олеиновой кислотой на печеночном пуле ЛПВП.
Кофактором и коферментом обеих изоформ ЛХАТ является А-II.
Обе изоформы (α и β) ЛХАТ синтезируются гепатоцитами и секретируются в кровь, где они образуют прочный комплекс с ЛПВП.
Этапы формирования ЛП
Метаболизм ЛП различных классов осуществляется по единому сценарию и складывается из 4 стадий:
1.Образование насцентных («новорождённых») ЛП
Насцентные ЛП – это незрелые ЛП-комплексы. Они являются продуктом ассоциации липидов и стационарных апобелков.
2.Трансформация насцентных ЛП в зрелые (плазменные) ЛП
Зрелые (плазменные) ЛП – это сформировавшиеся ЛП. После секреции насцентных ЛП в кровь, они соединяются с динамическими апобелками, получая возможность активировать ферменты, осуществляющие их метаболизм.
3. Ферментативная деградация зрелых (плазменных) ЛП с образованием ремнантных ЛП (старых)
Ремнантные ЛП – это плазменные ЛП, которые подверглись в кровотоке воздействию специфических ферментов с последующим качественным и количественным изменением их структуры.
4.Поглощение ремнантных ЛП клетками тканей путем рецептор-
опосредованного эндоцитоза.
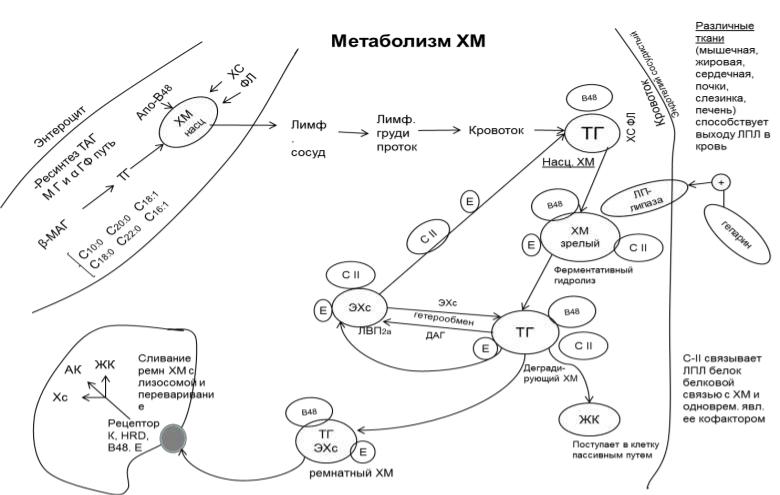
Метаболизм ХМ.
Функции ХМ
1.Транспортируют вновь синтезированные (ресинтезированные) ТАГ
(этерифицированные экзогенными насыщенными С16:0, С18:0, С20:0 ,С22:0 и
моноеновыми С18:1, С16:1 Ж.К) из кишечника к периферическим тканям и печени.
2. В составе ХМ транспортируются жирорастворимые витамины, поступающие в организм с пищей.
Формирование насцентных ХМ
После приёма пищи и всасывания в энтероцитах происходит ресинтез ТАГ из β-
МАГ (β-МАГ- путь). Глицерол этерифицируется насыщенными и моноевыми ЖК
(С16:0, С18:0, С20:0 ,С22:0 , С18:1, С16:1 ).
В-48 синтезируется в энтероцитах и связывает ТАГ, образуя белок-липидный комплекс, который покрывается монослоем ФЛ. Образовавшиеся насцентные ХМ содержат 90-95% ТАГ, 2% ФЛ и по 1-2% белка, ХС и ЭХС.
Созревание ХМ
Сформированные насцентные ХМ из энтероцита секретируются в лимфу (т.к.,
имея большие размеры, они не могут проходить через эндотелиальные клетки кровеносных сосудов), по лимфатическим коллекторам попадают в грудной лимфатический проток и далее в кровь, где происходит их созревание.
В процессе созревания ХМ приобретают динамические апобелки апо С-II и Апо Е, донором которых являются ЛПВП2α.
Ферментативная деградация ХМ и образование ремнантных ХМ
Зрелые ХМ током крови транспортируются к тканям, где на поверхностях капилляров подвергаюся гидролизу Е-гепаринзависимой ЛПЛ. (Она синтезируется клетками жировой ткани, сердечной и скелетными мышцами,
легкими). Апо-С II связывает ЛПЛ белок-белковой связью с ХМ и одновременно является ее кофактором. ЛПЛ гидролизует ТАГ гидрофобного ядра ХМ в α-
положении, расщеляя их на ЖК, ДАГ и β-МАГ.
ВЖК покидают ХМ и пассивным путем поглощаются клетками тканей и органов.
Т.о., перемещаясь по кровотоку, ХМ отдают часть ЖК тканям и постепенно
деградируют. Деградированные ХМ вступают во взаимодействие с ЛПВП2α и
отдают им часть ДАГ взамен забирая ЭХС. Процесс обмена ДАГ на ЭХС называется гетерообменом.
В результате гетерообмена изменяется липидный состав ХМ, повышается их плотность, отщепляются с поверхности динамические апобелки (апо-СII) и
переходят на ЛПВП2α , зрелые плазменные ХМ превращаются в ремнантные. Апо Е-остается.
Поглощение ремнантных ХМ гепатоцитами
Изменение липидного состава ядра ХМ приводит к изменению конформации АПО-В-48. Он взаимодействует с АПО Е, формируя лиганд, который связывается с рецептором Апо В-48/Е гепатоцита. Связавшись с рецептором, находящимся на гепатоците, ремнантные ХМ подвергаются эндоцитозу. Попав в гепатоцит,
ремнантный ХМ сливается с лизосомой и под действием лизосомальных ферментов распадается на мономеры: ДАГ и МАГ до глицерола и ЖК, белки - до АК, ЭХС - до ХС и ЖК. Т.о. ХМ доставляют часть пищевых липидов в печень.
Метаболизм ЛПОНП
Липиды, синтезированные в гепатоцитах, переносятся от печени к периферическим тканям. Этими транспортными формами являются ЛПОНП и ЛПНП.
Функции ЛПОНП
1.Транспортируют эндогенные ТАГ, эстерифицированные эндогенными насыщенными и моноеновыми ЖК из печени к периферическим тканям.
2.Являются предшественниками ЛПНП.
Формирование насцентных ЛПОНП
Синтез насцентных ЛПОНП начинается в печени. В печени синтезируются ТАГ из глицерола и ЖК. В эстерификации ТАГ принимают участие ЖК двух пулов:
а) поступившие в печень в составе ХМ – экзогенные (С16:0, С18:0, С20:0, С22:0)
б) синтезированные de novo из ацетил-SКоА углеводного происхождения – эндогенные (С16:0, С18:0). В гепатоцитах синтезируется стационарный белок Апо-
В-100, который связывает ТАГ, образуя комплекс. Затем этот комплекс покрывается гидрофильной оболочкой, состоящей из монослоя ФЛ.
Образовавшиеся т.о. насцентные ЛОНП содержат (%): ТАГ – 50-70, ФЛ – 13-20,
белок – 5-12, ХС и ЭХС – по 8.
Созревание ЛПОНП
Сформированные насцентные (т.е. новорожденные) ЛПОНП секретируются в кровоток, где они приобретают динамические белки Апо-С-II и Апо-Е (донором которых является ЛПВП2α) и становятся зрелыми. Апо-С-II необходим для дальнейшей деградации.
Ферментативная деградация ЛПОНП и образование ремнантных ЛПОНП
Зрелые ЛПОНП транспортируются к тканям ( миокард, скелетные мышцы, почки,
легкие), где на поверхности капилляров повергаются гидролизу гепаринзависимой ЛПЛ. ЛПЛ гидролизует ТАГ наружного слоя ЛПОНП в α-
положении (1 и 3), расщепляя их на ЖК, ДАГ и β-МАГ. ЖК пассивным путем поступают в клетки тканей, где служат основным энергетическим субстратом.
Метаболизм ХМ
Т.о., перемещаясь по кровотоку, ЛПОНП отдают часть ЖК тканям и постепенно деградируют. Деградированные ЛПОНП вступают во взаимодействие с ЛПВП2α и отдают им часть ДАГ, взамен забирая ЭХС.
Происходит гетерообмен. В результате гетерообмена изменяется липидный состав ЛПОНП, и с их поверхности отщепляется Апо-СII,
переходя на ЛПВП2α, а Апо-Е остается на ЛПОНП. В результате образуется ремнантный ЛПОНП.
Поглощение ремнантных ЛПОНП гепатоцитами
Ремнантные ЛПОНП делятся на 2 потока (пула):
а) «крупные» ремнантные ЛПОНП содержат в своем составе АПО В-100
и АПО-Е. «Крупные» ремнантные ЛПОНП связываются с рецепторами АПО В-100/Е гепатоцита, затем подвергаются эндоцитозу. Попав в гепатоцит, «крупные» ремнантные ЛПОНП сливаются с лизосомой и под действием лизосомальных ферментов распадаются на мономеры:
ДАГ и β-МАГ – до глицерола и жирных кислот (ЖК), ЭХС – до ХС и ЖК, белки – до аминокислот (АК).
б) «мелкие» ремнантные ЛПОНП содержат в своем составе АПО В-100
и очень небольшое количество АПО-Е.
Метаболизм и функции ЛПНП
«Мелкие» ремнантные ЛПОНП являются ЛППП, которые служат предшественниками ЛПНП (мелкие ЛПНОП (они же ЛППП) → ЛПНП).
За счет небольшого содержания АПО Е их «не узнают» В-100/Е-
рецепторы гепатоцитов, и они продолжают циркулировать в крови,
подвергаясь дальнейшему ферментативному гидролизу.
ЛППП подвергаются гидролизу под действием ПГГ, кофактором и переносчиком которой является СIII. ПГГ гидролизует труднодоступные ТАГ в α-положении (1 и 3 связь) ядра ЛППП до ДАГ, β-МАГ и ВЖК.