

Морфология растений / Нухимовский Е.Л. Основы биоморфологии семенных растений. III. Теория интегральной соматической эволюции
.pdf350 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
что-то новое в феносоматическом обиталище для генов — истинных хозяев и властелинов жизни на Земле, а возможно и где-то ещё.
В нашей работе уже не раз (т. 1, 2) обращалось внимание на то, что тело биоморфы, будучи одним миром для множества ресурсов, закономерно наполняющих его собой, образуют кроме того ещё несколько дополняющих друг друга систем (тел), например, геносому (генетически организованное тело в одном мире-теле биоморфы) и феносому (фенетически организованное тело в том же мире-теле биоморфы, которое является
атрибутивным обиталищем, образно говоря, «футляром» для генов). В этом обзоре мы обратим внимание ещё на две, взаимодополняющие друг друга, системы и тоже в одном мире-теле биоморфы (любой): вегетатив-
ной системе и системе размножения.
Б |
43 |
А |
Д |
|
|
10 |
|
у
2,15, |
31 |
19,21 |
|
 71
71
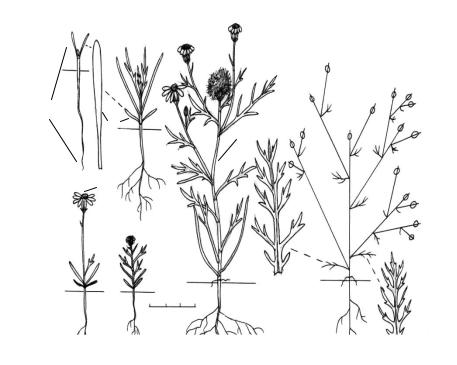
Рис. 204. Биографический портрет Crepis senecioides Delile [сем. Asteraceae
(там же, что и рис. 203, 10 III)].
Биоморфотип: надземно-двусемядольные (семядоли обратнояйцевидные, закруглённые, 4–6 мм дл., 2,5–3 мм шир., сидячие) полурозеточные очерёднолистные семидетерминант- но-репродуцирующие (полузакрытые, верхушки побегов закрываются открытыми бракте-
озными соцветиями — жёлтоцветковыми корзинками) однодомные стержнекорневые травянистые эвриэфемеры.
Единое генетическое содержание составляет преемственную основу отдельных биоморф (и организмов, и синорганизмов). По большому счёту, каждая отдельная биоморфа — это один соматический (телесный) мир, в котором на единой преемственной основе происходит интеграционное развитие определённого набора вещественных и пространственных ресурсов, который, пока существует (бытует) биоморфа, должен постоянно пополняться притоком новых ресурсов. В окружающем мире ресурсы не свободны; они всегда кому-то или чему-то принадлежат, поэтому за них как-либо приходится бороться в соответствии с универсальной стратегией

Глава 7. Статусное разнообразие органов семенных фитобиоморф |
351 |
Жизни (т. 1, гл. 3), т.е. стратегии наступательно-оборонительного жизне-
сохранения, и с использованием разнообразных тактических средств, из которых в приоритете тактика наступления, а в целом комбинированная тактика, которая в нашей работе рассматривается как агрессивно- компромиссно-камуфляжная экспансия.
А |
Б |
Д |
|
||
|
43 |
|
|
10 |
|
у |
2,15,19,21 |
|
1,14,18 
20,28 |
|
у |
|
|
|
|
|
71 |
|
|
Рис. 205. Биографический портрет Senecio vernalis Waldst. et Kit. [сем. Asteraceae (там же, что и рис. 199, 17 III)].
Биоморфотип: надземно-двусемядольные (семядоли продолговато-линейные, закруг-
лённые, 8–15 мм дл., 1–2мм шир., почти сидячие) полурозеточные или безрозеточные (при затенении) очерёднолистные семидетерминантно-репродуцирующие [полузакрытые, на
верхушках побегов развиваются открытые брактеозные соцветия — корзинки; «корзинки собраны в общее щитковидное соцветие, с краевыми язычковыми пестичными и срединными трубчатыми обоеполыми цветками; язычковые цветки в 1,5–2 раза длиннее обёртки, жёлтые» (Губанов и др., 2004, с. 480)] однодомные стержнекорневые травянистые эвриэфемеры.
Главной миссией в биоморфном развитии является завоевание веществ и пространства с последующим преобразованием их в разнообразные соматические структуры. Мало завоевать необходимые ресурсы и заполнить своими новообразованными структурами как-либо аннексированное у кого-то пространство, обеспечивая вегетативное развитие биоморф
посредством линейного роста осевых структур (стеблей, корней) и путём развёртывания аппендикулярных органов, в первую очередь листьев, для переработки завоёванных вещественных ресурсов. Вся соль (суть) проблемы жизнеразвития фитобиоморф (шире и других биоморф) в том, что оно не может быть вечным и всегда вегетативные наступления ограниче-
352 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
ны во времени, поскольку все носители жизни дискретны и по их генетическим программам они обязаны неизбежно когда-то умирать. Вполне по-
нятно, что принцип неизбежного отмирания вегетативных структур, тем более без образования органов размножения, уже первобытную жизнь на Земле устроить не мог. Наступательный порыв жизни в вегетативном развитии должен был не прекращаться и до, и после смерти родительских биоморф. Предковые фитобиоморфы довольно быстро, методом проб и ошибок, стали на путь производства системы размножения, которая нередко могла ещё долго рассеивать потомков в пространстве и после смерти родительских организмов; потомки часто отрываются от зрелых, но уже неживых репродуктивных структур. Великим завоеванием в эволюции стало создание системы размножения, которая поначалу была примитивной (простое деление и т.п.), но торопиться было некуда, и, в конце концов, сработал бинарный принцип жизнесохранения, в котором решающую роль приобрели две системы: вегетативного развития (личная) и система размножения (неличная).
А |
Б |
Д |
|
01к
10 |
2,15,20,21 |
|
 у
у
43
пп
Рис. 206. Биографический портрет Senecio glaucus L. [сем. Asteraceae (там же, что и рис. 197, 25 III)].
Биоморфотип: надземно-двусемядольные (семядоли продолговато-линейные, закруг-
лённые, 8–15 мм дл., 1–2 мм шир., почти сидячие) безрозеточные, редко полурозеточные, очерёднолистные семидетерминантно-репродуцирующие (полузакрытые, верхушки побегов закрывают открытые брактеозные соцветия — корзинки жёлто-астроцветковые, а раз-
ветвлённые побеги нередко формируют метёлку из корзинок) однодомные стержнекорневые травянистые эвриэфемеры.
Глава 7. Статусное разнообразие органов семенных фитобиоморф |
353 |
||
|
Б |
|
Д |
|
|
|
|
А |
|
|
|
|
|
|
39 |
10 |
у |
|
|
01к |
|
|
|
1кс |
|
|
|
43 |
41 |
|
|
|
|
||
|
|
2,15,20,21 |
|
2кс
 71
71
Рис. 207. Биографический портрет Medicago blancheana Boiss. [сем. Fabaceae (Израиль, окрестности города Бет-Шемеш, открытые горные склоны, 20 III)].
Биоморфотип:
15 мм дл., 2–3 мм шир., на коротких черешках 2–3 мм) безрозеточные очерёднолистные преиндетерминантно-репродуцирующие (приоткрытые, путём эфемерно-моноподиального
нарастания формируются сборные соцветия — открытые фрондозные кисти, в которых собраны все пазушные открытые брактеозные кисти из нескольких цветков, но только 1, реже 2 цветка таких кистей превращаются в бобы) однодомные стержнекорневые травянистые эвриэфемеры.
Пожалуй, есть смысл проблему разнообразных бинарных отношений «копнуть» поглубже: давайте «потревожим» и даже оголим бинарий личного (своего) — неличного (не-своего) в соматической организации
семенных фитобиоморф, который предстаёт перед нами, наблюдателями, как два мира-части, или два солитария (две стороны), занявшие в процессе
эволюции строго определённые места в едином теле биоморфы как две части одного интегрального мира — бинария. В чём главная особенность этого бинария? В соматической организации семенных фитобиоморф он представлен с одной стороны вегетативной основой биоморфы, которую я предлагаю называть кратко — «вегета» (от лат. vegetus — крепкий) се-
менного растения, а с другой стороны (второй солитарий единого бинария) — репродуктивной (собственно половое развитие) и (или) вицерепродуктивной (в случае с бесполой репродуктивностью) надстройкой, развивающейся на и над вегетативной основой, т.е., в первую очередь, за счёт ресурсов, добытых и произведённых ею (вегетой); такую «надстройку» можно назвать особенным термином — «изидой» (система полового и

354 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
бесполого размножения в теле биоморфы). Эту терминологию желательно достойным образом обосновать; почему именно предложенные здесь сло- ва-этикетки будут более всего уместны для отражения понятийной части
соответствующих терминов.
|
Б |
41 |
Д |
|
|
|
|
|
А |
1,14,20 |
|
|
|
|
|
01к |
10 |
пп |
|
|
39 |
||
|
|
 2,15,20,21
2,15,20,21
2к
71
Рис. 208. Биографический портрет Medicago polymorpha L. [сем. Fabaceae (там же, что и рис. 207, 20 III)].
Жизненные циклы биоморф и их биоморфотип во многом подобны тому, что приведено на рисунке 207 в отношении биоморф Medicago blancheana.
Термин, в моём понимании, — «это концептуальная динамическая система, характеризующаяся единством слова-этикетки (слова-символа) и
понятия (объяснения этого слова в смысле приемлемом для отражения явления-вещи, процесса определённого рода) [Нухимовский, 1997, с. 5].
Слово «вегета», на мой взгляд, уместно подчёркивает его привязку именно к вегетативной основе всего растительного организма и синорганизма, для которых её (вегеты) ростовая активность, на самом деле, придаёт фитобиоморфам силу, свежесть, крепость соматического развития. В качестве дублёра синонима может быть использовано сборное слово «корненабег» (vegeta). Дело в том, что сама вегета, будучи организована по бинарному
принципу, является интегральной силовой (ростовой) структурой в соматической организации семенных фитобиоморф, в первую очередь, предназначена для того, чтобы покорять (завоёвывать) пространство, на которое она буквально набегает (атакует его) своей листостебельной вегетативной частью — набегом, или рейдом (англ. «raid» означает «набег»), стремясь,
посредством ростовой активности, овладеть некоторыми его фрагментами,

Глава 7. Статусное разнообразие органов семенных фитобиоморф |
355 |
чтобы превратить их в места расположения своего тела и некоторых, например, репродуктивных, частей всего организма и одновременно ещё и потребить полезные для развития вещества, рассеянные в таком пространстве (рис. 211).
Набег общий (рейд) — листостебельная вегетативная основа всей
растительной биоморфы (целостная набеговая система) нередко дифференцируется на нижнюю часть (основание) и заметно обозначаемые в пространстве, в разной мере олиственные осевые структуры, восходящие от основания — главный и (или) парциальные (боковые или придаточные) — комплектующие набеги, или рейдеры (англ.).
Б
|
А |
Д |
|
|
|
01к |
|
у |
|
|
Рис. 209. Биографический портрет Helianthemum ledifolium (L.) Miller
[сем. Cistaceae (там же, что и рис. 197, 27 III)].
Биоморфотип: надземно-двусемядольные (семядоли эллиптические, закруглённые, 5–7
мм дл., 2,5–3,5 мм шир., на коротких черешках 1–3 мм) безрозеточные очерёднолистные и супротивнолистные (в нижней части растений) детерминантно-репродуцирующие (закрытые, на верхушках побегов развиваются закрытые фрондулёзно-брактеозные эфемерно-
полисимподиальные соцветия из жёлтолепестковых обоеполых цветков) стержнекорневые травянистые эвриэфемеры.
В ростовой деятельности набегам всецело помогает другая атрибутивная часть вегеты (единой вегетативной системы в соматической организации семенной фитобиоморфы) — это, как известно, корни. Среди корней различаются, согласно классической морфологии, главный и парциальные [боковые и (или) придаточные] корни. В вегете нередко бывает
удобно вычленять основные детали, или комплектующие части (фрагменты её интегральной организации), разные по их организационным особенностям. В частности, основание общего набега (рейда), рейдеры (главный и парциальные набеги), корни (главный и парциальные, включая придаточные) — это всё составляющие части вегеты (рис. 211), что можно так-

356 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
же обозначать ещё и одним словом, например, — «вегетикулы» (уменьшительное к слову «вегета», что подчёркивает соответствующий суффикс
— «cula») или иначе, как синоним, — «вегетаторы» [в словаре И.Х. Дворецкого (1976) дан такой перевод: лат. «vegetätor» — придающий силу, вносящий бодрость].
|
Б |
В |
|
|
А |
2к |
|
|
|
1кс |
|
|
у |
|
|
38 |
Д |
|
|
|
2,15,20,21 |
|
40,70 |
|
1к |
81 |
9к |
|
пп |
|
Рис. 210. Биографический портрет Helianthemum vesicarium Boiss. [сем.
Cistaceae (там же, что и рис. 197, 27 III)].
Биоморфотип: надземно-двусемядольные (семядоли широкоэллиптические, закруглён-
ные, 4–6 мм дл., 3–4 мм шир., резко клиновидно суженные в черешки 2–5 мм) безрозеточные супротивно-очерёднолистные (супротивность листорасположения больше проявляется в нижней части побегов) детерминантно-репродуцирующие (закрытые, образуются верху-
шечные одиночные розовые цветки, которые нередко перевершиниваются приростами также закрытыми одиночными цветками, т.е. проявляется эфемерно-полисимподиальное
нарастание соцветий) однодомные эусимподиальные (в вегетативной побеговой основе, где ежегодно после отмирания верхних и средних частей побегов сохраняются короткие надземные нижние части) стержнекорневые полукустарнички.
Итак, вегета (вегетативная система, или корненабег) — это соматическая система, состоящая только из вегетативных структур, которые, нередко даже, способны вторгаться в осевую организацию соцветий; в основе организации вегет всегда оси с разнообразными аппендикулярными компонентами [их натура листостебельная и (или) корневая]. Нередко при
анализе вегетативной системы как вегеты бывает не столь принципиально подчёркивать качественную принадлежность осевых структур (стеблевую, корневую или совместную). Главное для набегов и их наиболее активных составных частей (рейдеров), основных возмутителей спокойствия в соматической организации семенных фитобиоморф то, что они склонны, весьма уверенно и масштабно буквально рвать на части («куски») пространство (теперь, пожалуй, понятно почему в телах фитобиоморф «прописались» рейдеры, образно говоря, набеговые «приставы»), чтобы находить место в нём и для себя, и для структур иного типа, в первую очередь, тех, которые


 кн
кн 

 пг
пг рд
рд 
 кш
кш пн(а)
пн(а)
358 |
ОСНОВЫ БИОМОРФОЛОГИИ СЕМЕННЫХ РАСТЕНИЙ |
А
Д
 рл
рл
Бип
пп
фс 
ф |
кс |
|
Рис. 212*. Семенное возобновление и вегетативное разрастание после завершения очередной фазы цветения-плодоношения Chlorophytum
comosum (Thunb.) Jaques.
Б — всходы, Д — взрослое растение, рл — розеточные листья родительского растения, ип
— интеркалярные побеги с соцветиями, кс — открытые соцветия-кисти, ф — фугетты (ро-
зетки листьев на концах осей бывших соцветий, фс — сборная система фугетт, предваряющая вегетативное размножение, или фуга.
Особенности морфогенеза биоморф Chlorophytum comosum — это наглядный пример того,
как далеко может зайти компромисс во взаимоотношениях между вегетативными и репродуктивными структурами. Такой тип поведения семенных фитобиоморф можно обозначить как суперадаптированное (обеспеченное) пострепродуктивное побегоотрастние.
В отношении другого термина — «изида» предлагаю следующее обоснование. Представляется весьма заманчивой идея использовать для обозначения второго типа основных органов высшей статусной категории имя богини Изиды: «Исида, Изида — в древнеегипетской мифологии — богиня жизни, покровительница плодородия и материнства» (Сл. иностр. слов, 1964). Прецеденты к построению научных терминов подобным образом есть, например, широко известный сейчас термин «флора» взят из древнеримской мифологии. Флора — «богиня цветов, весны и юности». Насколько моё предложение рационально, покажет время, а уже сейчас я
Глава 7. Статусное разнообразие органов семенных фитобиоморф |
359 |
готов начать испытание «изиды» на прочность как нового ботанического термина в смысле системы репродуктивного (полового) и (или) вицерепродуктивного (бесполого) размножения. В качестве синонима термина «изида», на мой взгляд, вполне может подойти слово «генерата» [сокращение к широко известному слову «генеративный» (лат. generare — порождать, производить; generatio — рождение)]. Когда я предлагаю новые
термины, то нередко сопровождаю их синонимами, поскольку в научной практике со временем, по разным причинам, смогут привиться далеко не
все терминологические слова-этикетки.
Вершиной изидогенерации стала в эволюции система репродуктивного размножения (репродуктив). Бесполое размножение (вицерепродуктность) наблюдается у семенных фитобиоморф в соцветиях (вицерепродук-
тивах) немногих видов, например, Poa bulbosa L., Polygonum viviparum L.,
когда в их отдельных цветках или в соцветиях (целиком) без полового процесса образуются диаспоры особого типа, как результат процесса бесполого (вицерепродуктивного) размножения, которые уместно обозначать общим, только для них, образным, очень кратким термином «пигмы» (англ. pygmy –карлик, малорослый), например, «выводковые почечки»,
«выводковые побеги»; изиды в моём понимании могут иметь репродуктивное или вицерепродуктивное развитие (рис. 211). Все другие формы размножения относим к вегетативному размножению. Все проявления вегетативного размножения я осознанно не включаю в систему изидного (репродуктивного и вицерепродуктивного) размножения, поскольку они качественно отличаются друг от друга. Изида может состоять из одного цветка (простая) или из нескольких цветков (сложная). У кустистых растений общая изида нередко состоит из нескольких комплектующих изид
— главной и (или) парциальных (рис. 211).
Рейдеры (комплекстующие набеги) могут, таким образом, развивать на своих верхушках или изиды комплектующие (главные и парциальные), или специальные органы, адаптированные к вегетативному размножению,
—фугетты. Так, например, клубни Solanum tuberosum L., Aetheorhiza bulbosa (L.) Cass., вегетативные розеточные побеги на верхушках рейдеров
—остаточных осей соцветий, как-либо завершающих или уже завершивших своё репродуктивное развитие у Chlorophytum comosum (Thunb.) Jaques и др. (рис. 27, 212), — это всё фугетты.
Рейды (общие набеги) развивают и располагают на себе изиду общую, включающую всю систему репродуктивного и (или) вицерепродуктивного размножения, когда такие структуры образуются в организации семенной фитобиоморфы.
Таким образом, на верхушках рейдеров могут образовываться не только комплектующие изиды, но и специализированные органы вегетативного размножения — фугетты, что в процессе эволюции сыграло важное значение в преодолении противоречий, точнее в смягчении противоречий и усилении компромиссов между личным и неличным в организа-
ции семенных фитобиоморф. Так у Chlorophytum comosum (Liliaceae) от-
крытые отплодоносившие соцветия долго не отмирают и не несут угрозы для набеговой системы, которая успевает насытить рейдеры ростовой ак-