

Fiza_3
.pdfЗанятие № 3
1.
Синапс — это структурно-функциональное образование, обеспечивающее переход возбуждения или торможения с окончания нервного волокна на иннервирующую клетку.
Cтруктура синапса:
1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке); 2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс); 3) синаптическая щель (пространство между пресинаптической и постсинаптической
мембраной, заполнена жидкостью, которая по составу напоминает плазму крови).
Классификации:
По локализации:
-центральные синапсы;
-периферические синапсы.
По типу соединения с нейроном:
-аксосоматический, образованный аксоном одного нейрона и телом другого нейрона;
-аксодендритный, образованный аксоном одного нейрона и дендритом другого;
-аксоаксональный (аксон первого нейрона образует синапс на аксоне второго нейрона);
-дендродентритный (дендрит первого нейрона образует синапс на дендрите второго нейрона).
По функции:
-возбуждающие синапсы;
-тормозящие синапсы.
По механизму передачи возбуждения:
-химические;
-электрические.
По нейромедиатору (для химических):
-холинэргические (передача возбуждения при помощи ацетилхолина);
-адренэргические. (передача возбуждения при помощи трех катехоламинов);
-дофаминэргические (передача возбуждения при помощи дофамина);
-гистаминэргические (передача возбуждения при помощи гистамина);
-ГАМК-эргические (передача возбуждения при помощи гаммааминомасляной кислоты, т. е. развивается процесс торможения).
По строению и локализации (для периферических синапсов):
-мионевральный (нервно-мышечный), образованный аксоном мотонейрона и мышечной
клеткой;
-нервно-эпителиальный, образованный аксоном нейрона и секреторной клеткой.
Физиологические свойства:
-клапанное свойство синапсов, т. е. способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинаптическую;
-свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается;

-свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинаптической задержкой). Это связано с тем, что на пресинаптической и постсинаптической мембране остается медиатор от проведения предыдущего импульса;
-низкая лабильность синапса (100—150 имульсов в секунду).
2.
Химический синапс:
Передача в синапсе имеет два главных этапа. 1. Преобразование электрического сигнала в химический (электросекреторное
сопряжение). Потенциал действия (ПД), поступивший в пресинаптическое окончание, вызывает деполяризацию его мембраны, открывающую потенциалзависимые Са-каналы. Ионы кальция входят, согласно концентрационному и электрическому градиентам, внутрь клетки, что ведет к увеличению его содержания в цитозоле в 10—100 раз. Ионы кальция активируют фосфорилирование синаптосина, что ослабляет связь везикулы с цитоскелетом, и везикула перемещается вдоль микротрубочек на позицию у активной зоны. При контакте везикулы с пресинаптической мембраной происходит ферментативное «плавление» ее стенки, а также активация белка синаптопорина, формирующего канал, через который медиатор выходит в синаптическую щель посредством первично-активного транспорта — экзоцитоза. В нервно-мышечном синапсе медиатором является ацетилхолин, который образуется в пресинаптическом окончании из ацетилкоэнзима А и холина под действием фермента холинацетилтрансферазы. Впервые экспериментальное доказательство химического механизма передачи возбуждения в нервно-мышечном синапсе получил А.Ф.Самойлов (1924). Он показал, что скорость передачи возбуждения с нерва на мышцу в отличие от проведения возбуждения по нерву зависит от температуры в такой же степени , как и скорость химических реакций. Английский физиолог Г.Дейл
(1934) установил, что медиатором нервно-мышечного синапса является ацетилхолин. Этот медиатор был обнаружен одним из первых − он был известен также как «вещество блуждающего нерва» из-за своего действия на сердце.
Выделение молекул медиатора из пресинаптического окончания пропорционально количеству поступившего туда Са2+ в степени n = 4. Следовательно, химическое звено пресинаптического окончания работает как усилитель. Один из возможных механизмов усиления связан с тем, что поступивший в пресинаптическое окончание Са 2+ активирует рианодиновые рецепторы в цистернах эндоплазматической сети, имеющие в своем составе Са-каналы, что приводит к дополнительному выделению Са 2+в цитозоль из цистерн. Выделение ацетилхолина в синаптическую щель осуществляется квантами, каждый из которых в нервно-мышечном синапсе содержит от нескольких тысяч до 10 тыс. молекул. На один ПД из пресинаптического окончания нервно-мышечного синапса выделяется 200 —300 квантов медиатора. В промежутках между ПД из пресинаптического окончания происходит спонтанное выделение 1—2 квантов медиатора в синаптическую щель в течение 1 с.
Молекулы медиатора, поступившие в синаптическую щель, диффундируют к постсинаптической мембране и вступают во взаимодействие с ее рецепторами . В нервно-
мышечном синапсе ацетилхолин действует на Н-холинорецепторы, которые способны активизироваться и под влиянием никотина, вследствие чего они и получили свое название. Н-холинорецептор имеет в своем составе Nа/К-канал и состоит из пяти очень сходных субъединиц (α)2, β, γ, δ) примерно одинакового размера, группирующихся вокруг центрального канала. Такого рода макробелок составляет основу рецепторов различных типов, а для ацетилхолинового рецептора установлена его полная аминокислотная последовательность. Молекулярная масса этого белка 258 000. Открывание каналов в

химических синапсах происходит в результате связывания медиатора или его агониста с комплексом рецептор-канал.
Скорость диффузии молекул медиатора позволяет им пройти расстояние синаптической щели в течение 0,1—0,2 мс. Длительность действия медиатора на рецепторы постсинаптической мембраны, определенная по продолжительности открытия в ней ионных каналов, равна около 1 мс. Это значительно меньше периода полураспада медиатора и свидетельствует о его удалении из синаптической щели. Оно осуществляется путем диффузии ацетилхолина из щели в окружающую жидкость и разрушения его под действием ацетилхолинэстеразы.
Этот фермент выделяется миоцитом и прикрепляется к мукополисахаридному веществу в синаптической щели. Одна молекула ацетилхолинэстеразы может гидролизовать до ацетата и холина 10 молекул ацетилхолина в 1 мс, что обеспечивает его разрушение в синаптической щели в течение нескольких десятых долей миллисекунды. При этом большая часть (около 60 %) холина захватывается обратно пресинаптическим окончанием. Значительная доля высвобожденного ацетилхолина разрушается уже в ходе диффузии через синаптическую щель, не успевая достигнуть рецепторов, и через несколько миллисекунд его практически не остается: синапс вновь готов к передаче возбуждения.
2. Преобразование химического сигнала обратно в электрический. Этот этап
осуществляется в постсинаптической мембране. Действие молекул медиатора на ее рецепторы ведет к открытию ионных каналов и перемещению ионов, имеющих высокий электрохимический градиент на протяжении канала. Присоединение двух молекул ацетилхолина к α-субъединицам Н-холинорецептора открывает канал. Открытое состояние сохраняется 1 мс, в течение которой через него проходит около 500 000 ионов. Один квант медиатора (десятки тысяч его молекул) создает на несколько миллисекунд около рецепторов его высокую концентрацию, которая затем быстро падает. Начальный подъем концентрации медиатора повышает вероятность открывания канала, причем его открытые состояния перемежаются кратковременными закрываниями. После такой вспышки открываний он окончательно закрывается, потому что концентрация медиатора становится слишком низкой. Серии открываний суммируются, так что квант тока складывается из нескольких сотен токов одиночных каналов. Поскольку квант медиатора почти всегда вызывает только одну вспышку открываний, постоянная времени спада синаптического тока тоже примерно соответствует средней продолжительности такой вспышки.
Канал на внутреннем суженном конце имеет диаметр 0,65 нм, хорошо проницаем для Nа+ и К+, плохо проницаем для Са 2+. Поскольку канал имеет слабую избирательность в отношении Nа+ и К+, то ионные токи через канал зависят главным образом от электродвижущей силы (ЭДС) этих ионов.
ЭДС иона равна разности между мембранным потенциалом покоя и равновесным
потенциалом данного иона (ЭДС = МПП - Е |
иона). Отрицательная величина ЭДС |
характеризует движение иона в клетку, положительная — из клетки. |
|
В связи с этим входящий в клетку ток натрия (ЭДС = -140 мВ) резко преобладает над выходящим из клетки током калия (ЭДС = 14 мВ). Иными словами, ион Nа + движется в клетку согласно концентрационному и электрическому градиенту (клетка внутри имеет положительный заряд), а ион К + выходит из клетки только согласно концентрационному градиенту, причем вопреки электрическому (снаружи клетка имеет положительный заряд). Поэтому суммарный ток ионов Nа + в клетку превосходит ток К +из клетки, что и приводит к деполяризации постсинаптической мембраны (концевой пластинки). Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП), который в нервномышечном синапсе называют потенциалом концевой пластинки (ПКП). Особенностью

нервно-мышечного синапса фазного мышечного волокна является то, что при одиночной его активации формирующийся ПКП имеет большую амплитуду (30—40 мВ), которая превышает критический уровень деполяризации и вызывает генерацию ПД в миоците. Тоническое мышечное волокно имеет 7—10 синапсов, принадлежащих, как правило, нескольким мотонейронам. При этом ПКП не вызывает генерации ПД, а непосредственно запускает мышечное сокращение.
Как было отмечено, в промежутках между передачей нервного импульса происходит спонтанное выделение 1—2 квантов медиатора в синаптическую щель. При этом в постсинаптической мембране формируется деполяризация амплитудой 0,12—0,24 мВ, возникающая в среднем 1 раз в 1 с. Такие потенциалы, изученные в нервно-мышечном синапсе, были названы миниатюрными потенциалами концевой пластинки. Они,
вероятно, поддерживают высокую возбудимость синапсов в условиях функционального покоя нервных центров. Кроме экзоцитоза медиатора, существует постоянная неквантовая утечка молекул медиатора в синаптическую щель. Предполагают, что неквантовая секреция играет трофическую роль. Саморегуляция в синапсе осуществляется с использованием функциональных обратных
связей. Веществами, влияющими на эффективность синаптической передачи, могут быть медиаторы, продукты их распада. В нервно-мышечном синапсе ацетилхолин ,
выделившийся в небольшом количестве в синаптическую щель, может стимулировать более сильный выброс ацетилхолина из пресинаптического окончания по механизму обратной связи (самоусиление секреции). Высокие концентрации ацетилхолина в синаптической щели, напротив, угнетают секрецию его из пресинаптического окончания.
Показано, что холин (продукт гидролиза ацетилхолина) в концентрации 10 |
-4—10-5 М |
тормозит выделение ацетилхолина из пресинаптического окончания.
Электрический синапс:
После того как концепция химической синаптической передачи стала общепринятой, примерно между 1930 и 1950 гг., к большому удивлению специалистов выяснилось, что межклеточная передача возбуждения может осуществляться и электрическим способом [Furshpan,Potter,1959]. Две соседние клетки прилегают друг к другу так тесно, что сопротивление двух их мембран протекающему через них электрическому току сравнимо с сопротивлением остальной, внесинаптической области мембраны. При возбуждении 1- ой клетки натриевый ток (I Na) входит в нее через открытые Nа-каналы и выходит через пока невозбужденные участки мембраны; при этом часть тока входит через участок мембранного контакта во 2-ю клетку, вызывая ее деполяризацию. Разумеется, здесь уровень деполяризации гораздо ниже - скажем, в 10 раз, чем в 1-ой клетке, однако он может оказаться выше порога генерирования потенциала действия во 2-ой клетке. Часто такая деполяризация подпороговая, и тогда 2-ая клетка возбуждается только в результате суммации синаптических потенциалов, возникающих в результате химической или электрической передачи от других клеток.
Итак, перечислим основные характеристики, которые отличают химическую синаптическую передачу от электрической.
1. В химическом синапсе постсинаптический ток генерируется за счет открывания каналов в постсинаптической мембране и обусловлен ионными градиентами постсинаптической клетки. 2 . В э л е к т р и ч е с к о м с и н а п с е и с т о ч н и к п о с т с и н а п т и ч е с к о г о т о к а -
мембрана пресинаптической клетки. Здесь нет химического медиатора, и все факторы, влияющие на его высвобождение и действие (например, снижение внеклеточной концентрации Са 2+или устранение разрушающих медиатор ферментов), на передаче
возбуждения не сказываются. Щелевые контакты. Ионы, переносящие электрические токи, не могут проходить через
липидные мембраны, следовательно, для их транспорта в «мембранных контактах» между электрически сопряженными клетками необходимы канальные белки. Такие межклеточные связи называются нексусами, или «щелевыми контактами». В каждой из двух соседних клеточных мембран находятся регулярно распределенные через небольшие промежутки коннексоны, пронизывающие всю толщу мембраны; они расположены так, что в месте контакта клеток находятся друг против друга и их просветы оказываются на одной линии. У образованных таким образом каналов крупные диаметры и, значит, высокая проводимость для ионов: через них могут проходить даже относительно крупные молекулы с молекулярной массой до 1000 (около 1.5 нм в поперечнике). Коннексон состоит субъединиц числом до шести с молекулярной массой примерно 25000 каждая. Щелевые контакты обычны для ЦНС позвоночных и, как правило,соединяютгруппы синхронно функционирующих клеток. Такие контакты характерны также длябеспозвоночных. Функциональные синцитии. В тканях, не относящихся к нервной системе, клетки тоже очень часто соединены щелевыми контактами.
Говоря о передаче возбуждения, стоит, прежде всего, упомянуть миокард и гладкую мускулатуру, где эти контакты создают функциональный синцитий. Возбуждение здесь переходит от одной клетки к другой без заметной паузы или снижения амплитуды потенциала действия на границе.
Для таких органов важна регулируемость щелевых контактов; в самом деле, их каналы закрываются при снижении рН или повышении концентрации Са 2+. Это неизбежно происходит в случае повреждения клеток или глубокого нарушения обмена. За счет такого механизма пораженные места изолируются от остальной части функционального синцития, и распространение патологии ограничивается (например, при инфаркте миокарда). Кроме этих возбудимых тканей существует и много других (в частности, все эпителии, печень), где клетки также соединены щелевыми контактами. В принципе такая связь присуща любой клетке на ранних стадиях эмбрионального развития, когда все клетки соединены между собой щелевыми контактами и сохраняют их до стадии дифференцировки органов.
Роль таких контактов у невозбудимых клеток неясна. Через них возможен обмен многими мелкими молекулами; не исключено, что это важно для метаболизма. Через щелевые контакты могли бы также диффундировать внутриклеточные вторые посредники, передавая по ткани сигналы, регулирующие клеточные процессы.
Учитывая широкое распространение щелевых контактов, кажется удивительным, почему в нервной системе они не используются для синаптической передачи повсеместно. Видимо, сложнее организованные химические синапсы обеспечивают настолько более высокую специфичность и регулируемость межклеточной коммуникации, что в значительной степени вытеснили электрические.
Тормозные электрические синапсы. Щелевой контакт наиболее распространенный тип электрического синапса. Однако существуют и другие. Например, электрическим путем может передаваться и торможение. В этом случае потенциал действия особым образом расположенных пресинаптических волокон генерирует во внеклеточном пространстве вокруг постсинаптического аксона местный положительный потенциал такой амплитуды, что деполяризация аксона не может достичь порогового уровня, и проведение по нему потенциала действия блокируется.
Свойства электрических и химических синапсов

Свойство |
Электрические синапсы |
Х и м и ч е с к и е |
|
|
синапсы |
|
|
|
Проведение возбуждения |
двустороннее |
одностороннее |
|
|
|
Утомляемость |
низкая |
высокая |
|
|
|
Лабильность |
высокая |
низкая |
|
|
|
Синаптическая задержка |
короткая |
длинная |
|
|
|
Трансформация ритма ПД |
не происходит |
происходит |
|
|
|
Ч у в с т в и т е л ь н ы к |
э л е к т р о м а г н и т н ы х |
химических агентов |
действию |
излучений |
|
|
|
|
3.
Если кратко: слияние синаптических пузырьков с пресинаптической мембраной происходит, когда увеличивается концентрации Са2+ в цитозоле нервной терминали. Белок синаптического пузырька синаптотагмин связывается с Са 2+ и тем самым принимает участие в регуляции экзоцитоза (в том числе реорганизуя примембранный цитоскелет). Синапсины - белки, ассоциированные с везикулами, которые предположительно связывают синаптические пузырьки с цитоскелетом нервного окончания. Прикрепление, образование поры и опорожнение везикул осуществляются взаимодействиями (указаны стрелками) различных везикулярных и мембранных белков. Например, в образовании прикрепительного комплекса участвуют везикулярные белки (синаптотагмин и синаптобревины) и белки плазматической мембраны нервного окончания (синтаксины и нейрексины). Какие белки - плазматические или везикулярные образуют пору слияния до сих пор не ясно. Предположительно, это - синаптофизин (имеющий и другие функции) и белок плазматической мембраны физофилин. Rab-белки могут участвовать в транспорте везикул в клетке и в прикреплении их к мембране нервной клетки. Везикулярные транспортеры играют роль в аккумуляции нейромедиатора в синаптических пузырьках.
Одновременно Ca2+ начинает удаляться из цитоплазмы несколькими путями: связывание с белками, захват митохондриями и работа активного транспорта. Выход медиатора зависит от деполяризации терминали и составляет около 100 - 200 везикул, каждая из которых содержит одну порцию (квант), соответствующую приблизительно 104 молекул. Молекулы медиатора диффундируют к постсинаптической мембране, где взаимодействует с рецепторами постсинаптической мембраны, регулирующими состояние ионных каналов. Эта регуляция может быть прямой (как, например, в нервномышечном соединении скелетных мышц позвоночных), так и включать активацию систем вторичных внутриклеточных медиаторов ( G-белки, цАМФ).
А1 - медиатор, например, ацетилхолин (АХ) или глутамат, непосредственно действует на рецептор Na+/К+ - канала и открывает его (А2); Б1 - медиатор, например, серотонин (5- ГT), связывается с рецептором и стимулирует аденилатциклазу через соединяющий G- белок. При этом фосфорилируется связанный с G-белком ГДФ. Образующийся цАМФ активирует затем протеинкиназу, которая фосфорилирует (Ф) субcтратный белок (или сам канал, или регуляторный белок, контролирующий канал), в результате чего К+ -канал закрывается (Б2).
В первом случае осуществляется передача быстрых пусковых сигналов, во втором - осуществляются более медленные длительные воздействия. Направление изменения потенциала постсинаптической мембраны (деполяризация или гиперполяризация) зависит

главным образом от того, открытием каких каналов управляют постсинаптические рецепторы. Часть молекул медиатора может взаимодействовать с пресинаптическими рецепторами, что приводит к изменению МП нервной терминали и, соответственно, количества выделяемого медиатора (обратная связь). Синаптическая щель очищается от медиатора различными путями: дезактивация, гидролиз, обратный захват в пресинаптическое окончание, диффузия, захват глиальными клетками. Основная часть синаптической задержки - времени от прихода нервного импульса до развития постсинаптического ответа (0,2-0,5 мс) приходится на процесс секреции медиатора. Химический синапс обеспечивает передачу сигнала только от пресинаптического нейрона к постсинаптическому.
При частой ритмической стимуляции в химических синапсах наблюдается сначала усиление (облегчение), а затем ослабление (депрессия) передачи, т.е. рост, а затем падение амплитуды постсинаптических потенциалов. Эти явления в основном определяются изменениями в пресинаптическом звене. Они имеют особое развитие в некоторых синапсах ЦНС, где выступают как факторы синаптической пластичности. Наличие различных типов рецепторов в постсинаптической мембране может обусловливать развитие этих явлений по отдельности, как, например, длительную потенциацию (рис.11) и длительную депрессию (рис.12).
Обычный ход возбуждающей синаптической передачи. Единичный ПД в нервном окончании высвобождает глут амат (Глу), который может связываться с постсинаптическими глутаматными рецепторами. Одни из них - AMPA / каинатные (А/К)- рецепторы, каналы которых открываются после связывания с глутаматом. При этом ионы Na+, K+ и Ca2+ проходят через эти каналы, вызывая ВПСП амплитудой, например, 20 мВ. Соединение глутамата с другим типом рецепторов (NMDA-рецепторы) не дает эффекта, т.к. их каналы блокированы ионами Mg2+, которые связаны с внутренними стенками поры при отрицательных значениях МП. Справа - серия ПД вызывает длительную потенциацию за счет большего количества высвобождаемого глутамата. Более сильная деполяризация в этом случае (за счет большего числа активированных А/К каналов) приводит к разблокированию NMDA-каналов путем удаления из них ионов Mg2+. Возникающий относительно сильный Ca2+ ток увеличивает внутриклеточную концентрацию этих ионов, что ведет к активации специфической ферментной системы, чье действие в шипиках длительно повышает реакцию на глутамат. Вероятно, катализируется также образование молекул NO, которые могут диффундировать к пресинаптическому окончанию как ретроградный медиатор и там стимулировать выброс новых квантов глутамата, тем самым, способствуя длительной потенциации.
4.
Нервно-мышечный синапс образован окончанием аксона моторного нейрона и мышечным волокном поперечно-полосатой мускулатуры.
Структурами, непосредственно образующими синапс, являются пресинаптическая мембрана аксонной терминали, синаптическая щель и постсинантическая мембрана, являющаяся частью плазматической мембраны (сарколеммы) постсинаптической мышечной клетки. Пресинаптической мембраной называют часть не покрытой миелином мембраны аксонной терминали, обращенной в синаптическую щель.
Пресинаптическая терминаль формируется тонкой веточкой аксона, которая, подходя к мышечному волокну, образует утолщение (пуговку, бляшку, бутон). В пресинаптической терминали находятся синаптические пузырьки (везикулы), содержащие запасы медиатора ацетилхолина. Везикул может быть до нескольких тысяч. Их диаметр около 40 им, и в каждой содержится несколько тысяч молекул медиатора. В отсутствие поступления
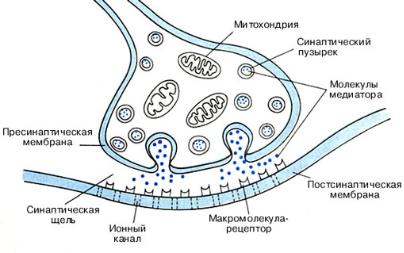
нервных импульсов везикулы с помощью белка синапсина связаны с цитоскелетом и малоподвижны. В пресинаптической терминали имеются также митохондрии, обеспечивающие выработку АТФ, ацетил СоА, белки цитоскелета, микротрубочки и микронити, по которым в терминаль из тела нейрона перемещается фермент ацетилхолинтрансфсраза. При участии этого фермента из ацетил СоА и холина образуется ацетилхолин.
Рис. 1. Строение нервно-мышечного синапса
Отличает нервно-мышечный синапс от центрального синапса большая протяженность пресинаптической мембраны, что способствует экзоцитозу большего количества медиатора. Этого количества медиатора достаточно для того, чтобы один потенциал действия, пришедший по нервному волокну, вызвал возбуждение мышечного волокна. В пресинаптической мембране, уже не покрытой миелиновой оболочкой, содержатся потенциалзависимые кальциевые каналы, преимущественно локализованные вблизи мест расположения везикул с медиатором. Такая локализация кальциевых каналов позволяет, изменяя разность потенциалов на пресинаптической мембране (и тем самым состояние каналов), идеально контролировать зависящий от концентрации кальция процесс высвобождения медиатора.
Между пре- и постсинаптическими мембранами располагается синаптическая щель шириной 50-100 им. Она заполнена межклеточным веществом и содержит тяжи плотного вещества из мукополисахаридов, с которым связана ацетилхолинэстераза (АХЭ) — фермент, разрушающий выходящий в синаптическую щель ацетилхолин на холин и уксусную кислоту.
Постсинаптическую мембрану нервно-мышечного синапса называют также концевой пластинкой. На ней имеются многочисленные впячивания, которые увеличивают площадь этой мембраны и в ней может расположиться до 20 миллионов рецепторных белковых молекул к ацетилхолину. Их плотность достигает 10 000 на 1 нм 2. Эти белки наряду с выполнением рецепторной функции формируют неселективные лигандзависимые каналы, через которые могут проходить ионы К+ и Na+. Рецепторы чувствительны также к никотину; их полное название — никотинчувствительные ацетилхолиновые рецепторы мышечного типа, или сокращенно н-ХРмт.
Когда нервный импульс, посланный моторным нейроном, распространяется по нервному волокну и достигает пресинаптической терминали, он вызывает деполяризацию ее мембраны.
Деполяризация приводит к открытию встроенных в мембрану потенциалзависимых кальциевых каналов, и ионы Са 2+' из межклеточного пространства поступают внутрь
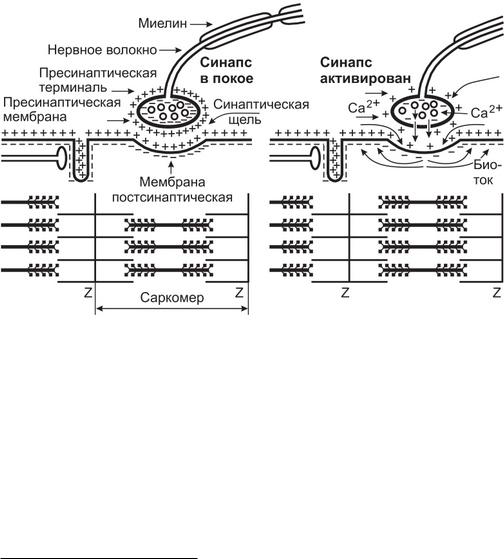
пресинаптичсской терминали. Ионы Са 2 перемещаются в терминаль по градиенту концентрации, так как снаружи аксонной терминали содержание кальция в 10 000 раз больше, чем внутри. Содержание кальция в цитоплазме терминали повышается, и это приводит к запуску ряда событий, необходимых для высвобождения ацетилхолина в синаптическую щель. Среди них отшнуровывание везикул от эндосом, приближение их к пресинаптической мембране, слияние с мембраной и экзоцитоз квантов ацетилхолина в синаптическую щель (рис. 2).
При поступлении к аксонной терминали одного ПД в синаптическую щель выбрасывается ацетилхолин из десятков синаптических пузырьков. Количество выделяемого АЦХ пропорционально величине и продолжительности деполяризации пресинаптической мембраны, которая в свою очередь определяется частотой и количеством поступивших по аксону нервных импульсов.
Молекулы ацетилхолина за время около 0,2 мс диффундируют к постсинаптической мембране и связываются с н-холинорецепторами, которые выполняют роль ворот в несслективных лигандзависимых ионных каналах, проницаемых для ионов Na+ и К+. Ворота открываются, и через ионные каналы внутрь мышечного волокна начинают входить ионы Na+, а из волокна выходить ионы КЛ Поток входящих ионов Na+ больше, чем поток выходящих ионов К+, так как ионы Na+ идут не только но градиенту концентрации, но и но градиенту электрического поля (на внутренней стороне мембраны отрицательный заряд до — 90 мВ).
Рис. 2. Структура нервно-мышечного синапса в покое и при активации
Положительно заряженные ионы Na+, входящие в мышечное волокно, деполяризуют постсинаптическую мембрану, нейтрализуя часть отрицательных зарядов на ее внутренней стороне. Амплитуда деполяризации зависит от количества выделившегося АЦХ и, следовательно, от числа нервных импульсов, поступивших к мышечному волокну от иннервирующего его мотонейрона. Она может достигать 40-50 мВ, длиться около 1 мс и сменяется реполяризацией за счет выхода из постсинаптической клетки катионов К . Возникающая деполяризация не сопровождается перезарядкой постсинаптической мембраны и развитием на ней ПД.
Кратковременное (около 4 мс) уменьшение разности потенциалов (деполяризация) постсинаптической мембраны нервно-мышечного синапса называют потенциалом концевой пластанки (ПКП) . По характеру влияния на постсинаптическую клетку он

аналогичен ВПСП. Деполяризация постсинаптической мембраны приводит к возникновению локального кругового электрического тока между нею и сарколеммой, граничащей с синапсом. В сарколемме, прилежащей к синапсу, имеются потенциалзависимые селекгивные быстрые натриевые и медленные калиевые каналы. Под действием локальных токов сарколемма деполяризуется и, если уровень деполяризации достигает Ек, каналы открываются и на граничащем с синапсом участке сарколеммы возникает потенциал действия.
Вестественных условиях при поступлении нервных импульсов и их успешном проведении через нервно-мышечный синапс, амплитуда возникающего ПКП всегда превышает пороговый уровень, необходимый для генерации ПД на сарколемме. Возникший ПД распространяется вдоль мышечного волокна по сарколемме и вглубь его по мембранам поперечных трубочек, пронизывающих волокно.
Почему же потенциал действия возникает на прилежащей к синапсу мембране, а не па постсинаптической мембране? Постсинаптическая мембрана не может генерировать ПД, так как на ней нет потенциалзависимых натриевых каналов, которые обеспечивают быстрый вход ионов Na+ и перезарядку мембраны. Перезарядке противодействуют также выходящие из постсинаптической клетки ионы К+. В то же время вход натрия, движимый силами его концентрационного и электрического градиентов, опережает выход калия, который обусловлен действием только сил концентрационного градиента калия и осуществляется против сил электрического поля. Вход натрия в мышечное волокно, опережающий выход калия, создает условия для кратковременной деполяризации постсинаптической мембраны и последующей ее реполяризации, т.е. для возникновения ПКП.
Ионные каналы постсинаптической мембраны остаются открытыми, пока концентрация ацетилхолина в синаптической щели не понизится примерно до 10 нмолей. Снижение концентрации АЦХ в синаптической щели в нормальных условиях происходит под действием фермента ацетилхолинэстеразы (АХЭ). Значение АХЭ для нормальной работы нервно-мышечного синапса чрезвычайно велико. Для того чтобы идущие от мотонейронов друг за другом нервные импульсы могли оказывать на рецепторы постсинаптической мембраны активирующее действие, необходимо к моменту поступления очередного импульса удалить из синаптической щели предшествующую порцию медиатора главным образом путем разрушения.
Когда содержание медиатора понизится до уровня 10 нмолей, ацетилхолин диссоциирует из связи с рецептором, восстанавливается способность рецепторов связываться с новой порцией АЦХ и открывать лигандзависимые ионные каналы. Синапс становится готовым к передаче нового сигнала. В устранении молекул ацетилхолина из синаптической щели также имеют значение обратный захват продукта его расщепления (холина) пресинаптической мембраной в пресинаптическую терминаль, диффузия АЦХ в интерстициальнос пространство и далее в кровь. Время от момента прихода ПД к пресинаптической мембране до момента возникновения ПД на мембране мышечного волокна называют синаптической задержкой . В нервно-мышечном синапсе она составляет около 1 мс.
Всостоянии покоя наблюдается спонтанное высвобождение (экзоцитоз) ацетилхолина в синаптическую щель. Объем экзоцитируемого медиатора составляет около 1 кванта в секунду, что эквивалентно количеству АЦХ, высвобождаемого из одной везикулы. Выход медиатора в этом объеме способен вызвать лишь малую (0,1-0,2 мВ) деполяризацию постсинаптической мембраны (миниатюрный потенциал концевой пластинки), и его недостаточно для инициации сокращения мышцы. Однако считается, что спонтанный