

Fiza_3
.pdfвыход этого небольшого количества медиатора имеет важное значение для трофического влияния АЦХ на постсинаптичсскую мышечную клетку: стимуляции синтеза каналообразующих рецепторных белков, регуляции обменных процессов в клетке, поддержания ее тканевой специфичности.
Таким образом, в нервно-мышечном синапсе сигнал электрической природы (нервный импульс) преобразуется в химический сигнал — высвобождение нейромедиатора АЦХ, который через цепочку последовательных событий на постсинаптической мембране вновь обеспечивает возникновение электрического потенциала на мембране мышечного волокна в виде ПД. Этот потенциал является непосредственной причиной инициирования сокращения постсинаптической мышечной клетки.
Имеется много факторов, способных влиять на состояние синаптической передачи сигналов к скелетным мышцам. Это влияние можно наблюдать в условиях патологии, например, при отравлении ботулиническим токсином С — одним из продуктов метаболизма анаэробного микроорганизма Clostridium botuinum, другими ядами растительного и животного происхождения. При попадании в организм ботулинического токсина он накапливается в аксонных терминалях нервно-мышечных синапсов и, обладая свойствами фермента цинкзависимой эндопептидазы, разрушает белки, участвующие в экзоцитозе ацетилхолина. Нейромышечная передача команд моторных нейронов к мышцам становится неэффективной или прекращается. Это может вести к развитию парезов, параличей скелетных мышц, нарушению глотания, дыхания и в случаях тяжелого отравления — к остановке дыхания.
С другой стороны, ряд веществ, способных влиять на различные этапы синаптической передачи, используется в качестве лекарственных. Так, если ботулинический токсин (ботокс) ввести в низких концентрациях в мышцу с повышенным тонусом, развившемся вследствие ее избыточной активации нервными импульсами, часто поступающими от мотонейронов, то дозированное снижение эффективности синаптической передачи может помочь снизить повышенный тонус, восстановить координацию движений. В настоящее время ботокс применяют для снижения степени тонического напряжения наружных глазных и других поперечно-полосатых мышц, например при косоглазии, кривошее, других спастических состояниях мышц.
Следующий способ воздействия на синаптическую передачу также связан с влиянием на количество ацетилхолина в синаптической щели через управление скоростью его расщепления под действием фермента АХЭ. Он реализуется с помощью применения веществ, ингибирующих ферментативную активность АХЭ. Ими являются вещества, обратимо ингибирующие активность АХЭ (лекарственные вещества эзерин, прозерин, галангамин, физостигмин и др.). Эти вещества, блокируя активность АХЭ, способствуют снижению скорости расщепления АЦХ и его накоплению в синаптической щели, пролонгированию и усилению действия АЦХ во всех холинергических, в том числе и нервно-мышечных, синапсах. Применение (в небольших дозах) блокаторов активности АХЭ обратимого действия позволяет облегчить передачу сигналов к мышце, повысить ее тонус и усилить сокращение. Их введение в организм в небольших дозах улучшает синаптическую передачу и оказывает лечебный эффекг при многих неврологических заболеваниях, в частности при миастении (myastenia gravis).
Однако передозировка этих веществ или применение веществ, необратимо ингибирующих АХЭ — инсектицидов, боевых отравляющих веществ нервно-паралитического действия (соединения фосфорорганической природы — зарин, зоман), сопровождается накоплением в синапсе большого количества АЦХ. Это ведет к десенситизации н-холинорецепторов, развитию стойкой, продолжительной деполяризации постсинаптической мембраны,

невозможности дальнейшей генерации ПД на мембране мышечного волокна, блокаде передачи сигналов к скелетным мышцам, их расслаблению, парезам, нарушению или остановке дыхания.
Ряд веществ может легко связываться с никотинчувствительными холинорецепторами и блокировать при этом открытие лигандзависимых ионных каналов. Это, например, такие вещества, как кураре, D-тубокурарин, и вещества, входящие в состав ядов — кобратоксин, а-бунгаротоксин. Примененние кураре и курареподобных веществ позволяет, заблокировав взаимодействие АЦХ с холинорецепторами постсинаптической мембраны, вызывать блокаду передачи сигнала от мотонейронов к мышечным волокнам, расслабление мышц во время хирургических операций (миорелаксация) или при повышении их тонического напряжения при спастичесих состояних.
При некоторых аутоиммунных заболеваниях, например при myastenia gravis, в организме вырабатываются антитела к ацетилхолиновым рецепторам постсинаптической мембраны нервно-мышечных синапсов. Антитела могут блокировать рецепторы и разрушать их. В этих условиях даже при высвобождении достаточного количества АЦХ из нервного окончания амплитуда постсинаптичсского потенциала часто не достигает величины, необходимой для генерации ПД на мембране мышечного волокна. Вследствие длительного нарушения нервно-мышечной передачи развиваются слабость и повышенная утомляемость мышц. Введение больным миастенией ботокса или блокаторов АХЭ обратимого действия, улучшающих нервно-мышечную передачу, даже при сниженном количестве рецепторов приводит к повышению силы сокращения и работоспособности мышц.
Если кратко: Механизмы передачи возбуждения в синапсах на примере мионеврального синапса
Мионевральный (нервно-мышечный) синапс — образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптиче ской щели. По сле диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом:
ХР—ХЭ—ХР—ХЭ—ХР—ХЭ.
ХР + АХ = МПКП — миниатюрные потенциалы концевой пластины.

Затем происходит суммация МПКП. В результате суммации образуется ВПСП —
возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
ХЭ + АХ = разрушение АХ до холина и уксусной кислоты.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности . Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1—2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
5.
Нервный центр — это совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции определенной функции.
Основными клеточными элементами нервного центра являются многочисленные нейроны, скопление которых формирует нервные ядра. В состав центра могут входить нейроны, рассеянные за пределами ядер. Нервный центр может быть представлен структурами мозга, располагающимися на нескольких уровнях центральной нервной системы (например, центры регуляции дыхания, кровообращения, пищеварения).
Любой нервный центр состоит из ядра и периферии.
Ядерная часть нервного центра представляет собой функциональное объединение нейронов, в которое поступает основная информация от афферентных путей. Повреждение этого участка нервного центра приводит к повреждению или существенному нарушению осуществления данной функции.
Периферическая часть нервного центра получает небольшую порцию афферентной информации, и ее повреждение вызывает ограничение или уменьшение объема выполняемой функции (рис. 1).
Функционирование центральной нервной системы осуществляется благодаря деятельности значительного числа нервных центров, представляющих собой ансамбли нервных клеток, объединенных с помощью синаптических контактов и отличающихся огромным разнообразием и сложностью внутренних и внешних связей.
Рис. 1. Схема общего строения нервного центра В нервных центрах выделяют следующие иерархические отделы: рабочие, регуляторные и исполнительные (рис. 2).
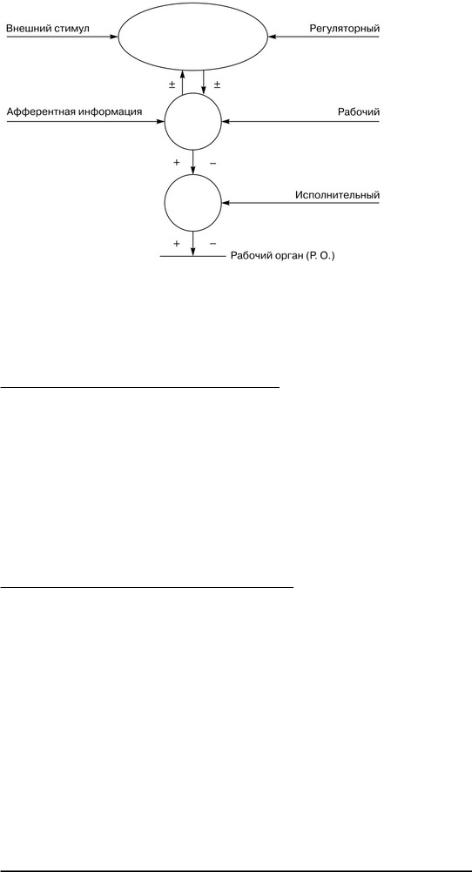
Рис. 2. Схема иерархического подчинения разных отделов нервных центров
Рабочий отдел нервного центра ответствен за осуществление данной функции. Например, рабочий отдел дыхательного центра представлен центрами вдоха, выдоха и пневмотаксиса, расположенными в продолговатом мозге и варолиевом мосту; нарушение этого отдела вызывает остановку дыхания.
Регуляторный отдел нервного центра - это центр, расположенный в коре больших полушарий мозга и регулирующий активность рабочего отдела нервного центра. В свою очередь, активность регуляторного отдела нервного центра зависит от состояния рабочего отдела, который получает афферентную информацию, и от внешних стимулов среды. Так, регуляторный отдел дыхательного центра расположен в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция небезгранична и зависит от функциональной активности рабочего отдела, афферентной им пульсации, отражающей состояние внутренней среды (в данном случае рН крови, концентрации углекислого газа и кислорода в крови).
Исполнительный отдел нервного центра - это двигательный центр, расположенный в спинном мозге и передающий информацию от рабочего отдела нервного центра к рабочим органам. Исполнительный отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам.
С другой стороны, одни и те же нейроны головного и спинного мозга могут участвовать в регуляции разных функций. Например, клетки центра глотания участвуют в регуляции не только акта глотания, но и акта рвоты. Этот центр обеспечивает все последовательные стадии акта глотания: движение мышц языка, сокращение мышц мягкого неба и его поднятие, последующее сокращение мышц глотки и пищевода при прохождении пищевого комка. Эти же нервные клетки обеспечивают сокращение мышц мягкого нёба и его поднятие во время акта рвоты. Следовательно, одни и те же нервные клетки входят и в центр глотания, и в центр рвоты.
6.
6.1. Одностороннее проведение возбуждение в нервном центре: возбуждение в ЦНС проводится в одном направлении с аксона на дендрит или тело клетки следующего нейрона. Основу этого свойства составляют особенности морфологической связи между нейронами.
Одностороннее проведение возбуждения зависит от строения синапса и гуморальной природы передачи в нем импульса: медиатор, осуществляющий передачу возбуждения, выделяется только в пресинаптическом окончании, а рецепторы, воспринимающие медиатор, расположены на постсинаптической мембране;
6.2. Задержка в проведение возбуждения (центральная задержка): в системе рефлекторной дуги медленнее всего проводится возбуждение в синапсах ЦНС. В связи с этим центральное время рефлекса зависит от количества вставочных нейронов.
Чем сложнее рефлекторная реакция, тем больше центральное время рефлекса. Его величина связана со сравнительно медленным проведением возбуждения через последовательно включенные синапсы. Замедление проведения возбуждения создается вследствие относительной длительности осуществляющихся в синапсах процессов: выделения медиатора через пресинаптическую мембрану, его диффузии через синаптическую щель, возбуждения постсинаптической мембраны, возникновения возбуждающего постсинаптического потенциала и его перехода в потенциал действия;
6.3. Пространственная и временная суммация:
Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда возбуждения постсинаптических потенциалов (ВПСП). Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторных нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются и в мембране нейрона генерируется распространяющийся ПД.
6.4. Трансформация ритма возбуждения: нервные центры способны изменять ритм поступающих к ним импульсов. Они могут отвечать на одиночные раздражители серией импульсов или на раздражители небольшой частоты — возникновением более частых потенциалов действия. В результате ЦНС посылает к рабочему органу количество импульсов, относительно независимое от частоты раздражений.
Это связано с тем, что нейрон является изолированной единицей нервной системы, к нему в каждый момент приходит множество раздражений. Под их влиянием происходит изменение мембранного потенциала клетки. Если создается небольшая, но продолжительная деполяризация (длительный возбуждающий постсинаптический потенциал), то один стимул вызывает серию импульсов (рис. 3);
Рис. 3. Схема трансформации ритма возбуждения
6.5. Рефлекторное последействие - способность сохранять возбуждение после окончания действия раздражителя, т.е. афферентных импульсов нет, а эфферентные продолжают действовать еще некоторое время.
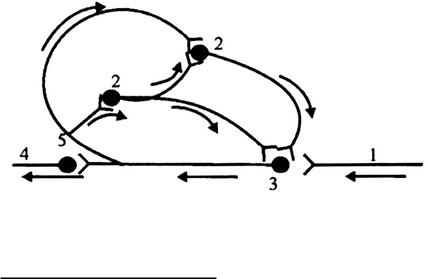
Последействие объясняется наличием следовой деполяризации. Если следовая деполяризация длительна, то на ее фоне в течение нескольких миллисекунд могут возникать потенциалы действия (ритмическая активность нейрона), вследствие чего сохраняется ответная реакция. Но это дает сравнительно короткий эффект последействия. Более длительное последействие связано с наличием кольцевых связей между нейронами. В них возбуждение как бы само себя поддерживает, возвращаясь по коллатералям к первоначально возбужденному нейрону (рис. 4);
Рис. 4. Схема кольцевых связей в нервном центре (по Лоренто де Но): 1 — афферентный путь; 2-промежуточные нейроны; 3 — эфферентный нейрон; 4 — эфферентный путь; 5 — возвратная ветвь аксона 6.6. Тонус нервных центров - состояние постоянной повышенной активности. Он
обусловлен постоянным поступлением к НЦ нервных импульсов от периферических рецепторов, возбуждающим влиянием на нейроны продуктов метаболизма и других гуморальных факторов. Например, проявлением тонуса соответствующих центров является тонус определенной группы мышц 6.7. Утомляемость нервного центра: нервный центр обладает малой лабильностью. Он
постоянно получает от множества высоколабильных нервных волокон большое количество стимулов, превышающих его лабильность. Поэтому нервный центр работает с максимальной загрузкой и легко утомляется.
Исходя из синаптических механизмов передачи возбуждения утомление в нервных центрах может объясняться тем, что по мере работы нейрона истощаются запасы медиатора и становится невозможной передача импульсов в синапсах. Кроме того, в процессе деятельности нейрона наступает постепенное снижение чувствительности его рецепторов к медиатору, что называется десенситизацией;
6.8 Чувствительность нервных центров к кислороду и некоторым фармакологическим веществам: в нервных клетках осуществляется интенсивный обмен веществ, для чего необходимы энергия и постоянный приток нужного количества кислорода.
Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга, после пяти-шести минут кислородного голодания они погибают. У человека даже кратковременное ограничение мозгового кровообращения приводит к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола, их функция восстанавливается через 15-20 мин после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливаются даже после 30 мин отсутствия кровообращения.
По сравнению с нервным центром нервное волокно малочувствительно к недостатку кислорода. Помешенное в атмосферу азота, оно только через 1,5 ч прекращает проведение возбуждения.
6.9.
Проявление и осуществление рефлекса возможно только при ограничении распространения возбуждения с одних нервных центров на другие. Это достигается взаимодействием возбуждения с другим нервным процессом, противоположным по эффекту процессом торможения.
Почти до середины XIX века физиологи изучали и знали только один нервный процесс - возбуждение.
Явления торможения в нервных центрах, т.е. в центральной нервной системе были впервые открыты в 1862 году И.М.Сеченовым ("сеченовское торможение”). Это открытие сыграло в физиологии не меньшую роль, чем сама формулировка понятия рефлекса, так как торможение обязательно участвует во всех без исключения нервных актах. И.М.Сеченов обнаружил явление центрального торможения при раздражении промежуточного мозга теплокровных. В 1880 году немецкий физиолог Ф.Гольц установил торможение спинальных рефлексов. Н.Е. Введенский в результате серий опытов по парабиозу вскрыл интимную связь процессов возбуждения и торможения и доказал, что природа этих процессов едина.
Торможение - местный нервный процесс, приводящий к угнетению или предупреждению возбуждения. Торможение является активным нервным процессом, результатом которого служит ограничение или задержка возбуждения. Одна из характерных черт тормозного процессаотсутствие способности к активному распространению по нервным структурам. В настоящее время в центральной нервной системе выделяют два вида торможения: торможение центральное (первичное), являющееся результатом возбуждения (активации) специальных тормозных нейронов и торможение вторичное, которое осуществляется без участия специальных тормозных структур в тех самых нейронах в которых происходит возбуждение.
Центральное торможение (первичное) - нервный процесс, возникающий в ЦНС и приводящий к ослаблению или предотвращению возбуждения. Согласно современным представлениям центральное торможение связано с действием тормозных нейронов или синапсов, продуцирующих тормозные медиаторы (глицин, гаммааминомасляную кислоту), которые вызывают на постсинаптической мембране особый тип электрических изменений, названных тормозными постсинаптическими потенциалами (ТПСП) или деполяризацию пресинаптического нервного окончания, с которым контактирует другое нервно е окончание аксона. По этому выделяют цент рально е (первично е) постсинаптическое торможение и центральное (первичное) пресинаптическое торможение.
Постсинаптическое торможение (лат. post позади, после чего-либо + греч. sinapsis
соприкосновение, соединение) - нервный процесс, обусловленный действием на постсинаптическую мембрану специфических тормозных медиаторов (глицин, гаммааминомаслянная кислота), выделяемых специализированными пресинаптическими нервными окончаниями. Медиатор, выделяемый ими, изменяет свойства постсинаптической мембраны, что вызывает подавление способности клетки генерировать возбуждение. При этом происходит кратковременное повышение проницаемости постсинаптической мембраны к ионам К+ или CI −, вызывающее снижение ее входного электрического сопротивления и генерацию тормозного постсинаптического потенциала (ТПСП). Возникновение ТПСП в ответ на афферентное раздражение обязательно связано

с включением в тормозной процесс дополнительного звена - тормозного интернейрона, аксональные окончания которого выделяют тормозной медиатор. Специфика тормозных постсинаптических эффектов впервые была изучена на мотонейронах млекопитающих (Д. Экклс, 1951). В дальнейшем первичные ТПСП были зарегистрированы в промежуточных нейронах спинного и продолговатого мозга, в нейронах ретикулярной формации, коры больших полушарий, мозжечка и таламических ядер теплокровных животных.
Известно, что при возбуждении центра сгибателей одной из конечностей центр ее разгибателей тормозится и наоборот. Д. Экклс выяснил механизм этого явления в следующем опыте. Он раздражал афферентный нерв, вызывающий возбуждение мотонейрона, иннервирующего мышцу - разгибатель.
Нервные импульсы, дойдя до афферентного нейрона в спинномозговом ганглии, направляются по его аксону в спинном мозге по двум путям: к мотонейрону, иннервирующему мышцу - разгибатель, возбуждая ее и по коллатерам к промежуточному тормозному нейрону, аксон которого контактирует с мотонейроном иннервирующим мышцу - сгибатель, вызывая таким образом торможение антагонистической мышцы. Этот вид торможения был обнаружении в промежуточных нейронах всех уровней центральной нервной системы при взаимодействии антагонистических центров. Он был назван поступательным постсинаптическим торможением . Этот вид торможения координирует, распределяет процессы возбуждения и торможения между нервными центрами.
Возвратное (антидромное) постсинаптическое торможение (греч. antidromeo бежать в противоположном направлении) - процесс регуляции нервными клетками интенсивности поступающих к ним сигналов по принципу отрицательной обратной связи. Он заключается в том, что коллатерали аксонов нервной клетки устанавливают синаптические контакты со специальными вставочными нейронами (клетки Реншоу), роль которых заключается в воздействии на нейроны, конвергирующие на клетке, посылающей
эти аксонные коллатерали (рис. 87). По такому принципу осуществляется |
торможение |
мотонейронов. |
|
Возникновение импульса в мотонейроне млекопитающих не только активирует мышечные волокна, но через коллатерали аксона активирует тормозные клетки Реншоу. Последние устанавливают синаптические связи с мотонейронами. Поэтому усиление импульсации мотонейрона ведет к большей активации клеток Реншоу, вызывающей усиление торможения мотонейронов и уменьшение частоты их импульсации. Термин "антидромное” употребляется потому, что тормозной эффект легко вызывается антидромными импульсами, рефлекторно возникающими в мотонейронах.
Чем сильнее возбужден мотонейрон, чем больше сильные импульсы идут к скелетным мышцам по его аксону, тем интенсивнее возбуждается клетка Реншоу, которая подавляет активность мотонейрона. Следовательно, в нервной системе существует механизм, оберегающий нейроны от чрезмерного возбуждения. Характерная особенность постсинаптического торможения заключается в том, что оно подавляется стрихнином и столбнячным токсином (на процессы возбуждения эти фармакологические вещества не действуют).
В результате подавления постсинаптического торможения нарушается регуляция возбуждения в цнс, возбуждение разливается ("диффундирует”) по всей цнс, вызывая перевозбуждение мотонейронов и судорожные сокращения групп мышц (судороги).
Торможение ретикулярное (лат. reticularis - сетчатый) - нервный процесс,
развивающийся в спинальных нейронах под влиянием нисходящей импульсации из ретикулярной формации (гигантское ретикулярное ядро продолговатого мозга). Эффекты,

создаваемые ретикулярными влияниями, по функциональному действию сходны с возвратным торможением, развивающимся на мотонейронах. Влияние ретикулярной формации вызывают стойкие ТПСП, охватывающие все мотонейроны независимо от их функциональной принадлежности. В этом случае, так же, как и при возвратном торможении мотонейронов происходит ограничение их активности. Между таким нисходящим контролем со стороны ретикулярной формации и системочй возвратного торможения через клетки Реншоу существует определенное взаимодействие, и клетки Реншоу находятся под постоянным тормозящем контролем со стороны двух структур. Тормозящее влияние со стороны ретикулярной формации являются дополнительным фактором в регуляции уровня активности мотонейронов.
Первичное торможение может вызываться механизмами иной природы, не связанными с изменениями свойств постсинаптической мембраны. Торможение в этом случае возникает на пресинаптической мембране (синаптическое и пресинаптическое торможение).
Синаптическое торможение (греч. sunapsis соприкосновение, соединение) - нервный процесс, основанный на взаимодействии медиатора, секретируемого и выделяемого пресинаптическими нервными окончаниями, со специфическими молекулами постсинаптической мембраны. Возбуждающий или тормозной характер действия медиатора зависит от природы каналов, которые открываются в постсинаптической мембране. Прямое доказательство наличия в цнс специфических тормозящих синапсов было впервые получено Д. Ллойдом (1941).
Данные относительно электрофизиологических проявлений синаптического торможения: наличие синаптической задержки, отсутствие электрического поля в области синаптических окончаний дали основание считать его следствием химического действия особого тормозящего медиатора, выделяемого синаптическими окончаниями. Д. Ллойд показал, что если клетка находится в состоянии деполяризации, то тормозной медиатор в ы з ы в а е т г и п е р п о л я р и з а ц и ю, в т о в р е м я к а к н а ф о н е гиперполяризации постсинаптической мембраны он вызывает ее деполяризацию.
Пресинаптическое торможение (лат. praе -впереди чего-либо + греч. sunapsis
соприкосновение, соединение) - частный случай синаптических тормозных процессов, проявляющихся в подавлении активности нейрона в результате уменьшения эффективности действия возбуждающих синапсов еще на пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мембраны не подвергаются каким-либо изменениям. Пресинаптическое торможение осуществляется посредством специальных тормозных интернейронов. Его структурной основой являются аксо-аксональные синапсы, образованные терминалиями аксонов тормозных интернейронов и аксональными окончаниями возбуждающих нейронов.
При этом окончание аксона тормозного нейрона является пресимпатическим по от н о ш е н и ю к т е рм и н а л и во з буж д а ю щ е го н е й р о н а, кото р а я о ка з ы ва е т с я постсинаптической по отношению к тормозному окончанию и пресинаптической по отношению к активируемой им нервной клетки. В окончаниях пресинаптического тормозного аксона освобождается медиатор, который вызывает деполяризацию
возбуждающих окончаний за счет увеличения проницаемости их мембраны для CI −. Деполяризация вызывает уменьшение амплитуды потенциала действия, приходящего в возбуждающее окончание аксона. В результате происходит угнетение процесса высвобождения медиатора возбуждающими нервными окончаниями и снижение амплитуды возбуждающего постсинаптического потенциала.

Характерной особенностью пресинаптической деполяризации является замедленное развитие и большая длительность (несколько сотен миллисекунд), даже после одиночного афферентного импульса.
Пресинаптическое торможение существенно отличается от постсинаптического и в фармакологическом отношении. Стрихнин и столбнячный токсин не влияют на его течение. Однако наркотизирующие вещества (хлоралоза, нембутал) значительно усиливают и удлиняют пресинаптическое торможение. Этот вид торможения обнаружен в различных отделах цнс. Наиболее часто оно выявляется в структурах мозгового ствола и спинного мозга. В первых исследованиях механизмов пресинаптического торможения считалось, что тормозное действие осуществляется в точке, отдаленной от сомы нейрона, поэтому его называли "отдаленным” торможением.
Функциональное значение пресинаптического торможения, охватывающего пресинаптические терминали, по которым поступают афферентные импульсы, заключается в ограничении поступления к нервным центрам афферентной импульсации. Пресинаптическое торможение в первую очередь блокирует слабые асинхронные афферентные сигналы и пропускает более сильные, следовательно, оно служит механизмом выделения, вычленения более интенсивных афферентных импульсов из общего потока. Это имеет огромное приспособительное значение для организма, так как из всех афферентных сигналов, идущих к нервным центрам, выделяются самые главные, самые необходимые для данного конкретного времени. Благодаря этому нервные центры, нервная система в целом освобождается от переработки менее существенной информации. Вторичное торможение- торможение осуществляющееся теми же нервными структурами, в которых происходит возбуждение. Этот нервный процесс подробно изложен в работах Н.Е. Введенского (1886, 1901г.г.).
Торможение реципрокное (лат. reciprocus - взаимный) - нервный процесс, основанный на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через посредство вставочных нейронов торможение других групп клеток. Реципрокные отношения возбуждения и торможения в цнс были открыты и продемонстрированы Н.Е. Введенским: раздражение кожи на задней лапке у лягушки вызывает ее сгибание и торможение сгибания или разгибания на противоположной стороне. Взаимодействие возбуждения и торможения является общим свойством всей нервной системы и обнаруживается как в головном, так и в спинном мозге. Экспериментально доказано, что нормальное выполнение каждого естественного двигательного акта основано на взаимодействии возбуждения и торможения на одних и тех же нейронах цнс.
Общее центральное торможение - нервный процесс, развивающийся при любой рефлекторной деятельности и захватывавающий почти всю цнс, включая центры головного мозга. Общее центральное торможение обычно проявляется раньше возникновения какой-либо двигательной реакции. Оно может проявляться при такой малой силе раздражения при которой двигательный эффект отсутствует. Такого вида торможение было впервые описано И.С. Беритовым (1937). Оно обеспечивает концентрацию возбуждения других рефлекторных или поведенческих актов, которые могли бы возникнуть под влиянием раздражений. Важная роль в создании общего центрального торможения принадлежит желатинозной субстанции спинного мозга. При электрическом раздражении желатинозной субстанции у спинального препарата
кошки происходит общее торможение рефлекторных реакций, вызываемых раздражением сенсорных нервов. Общее торможение является важным фактором в создании целостной