

3.2. Электрический синапс
С
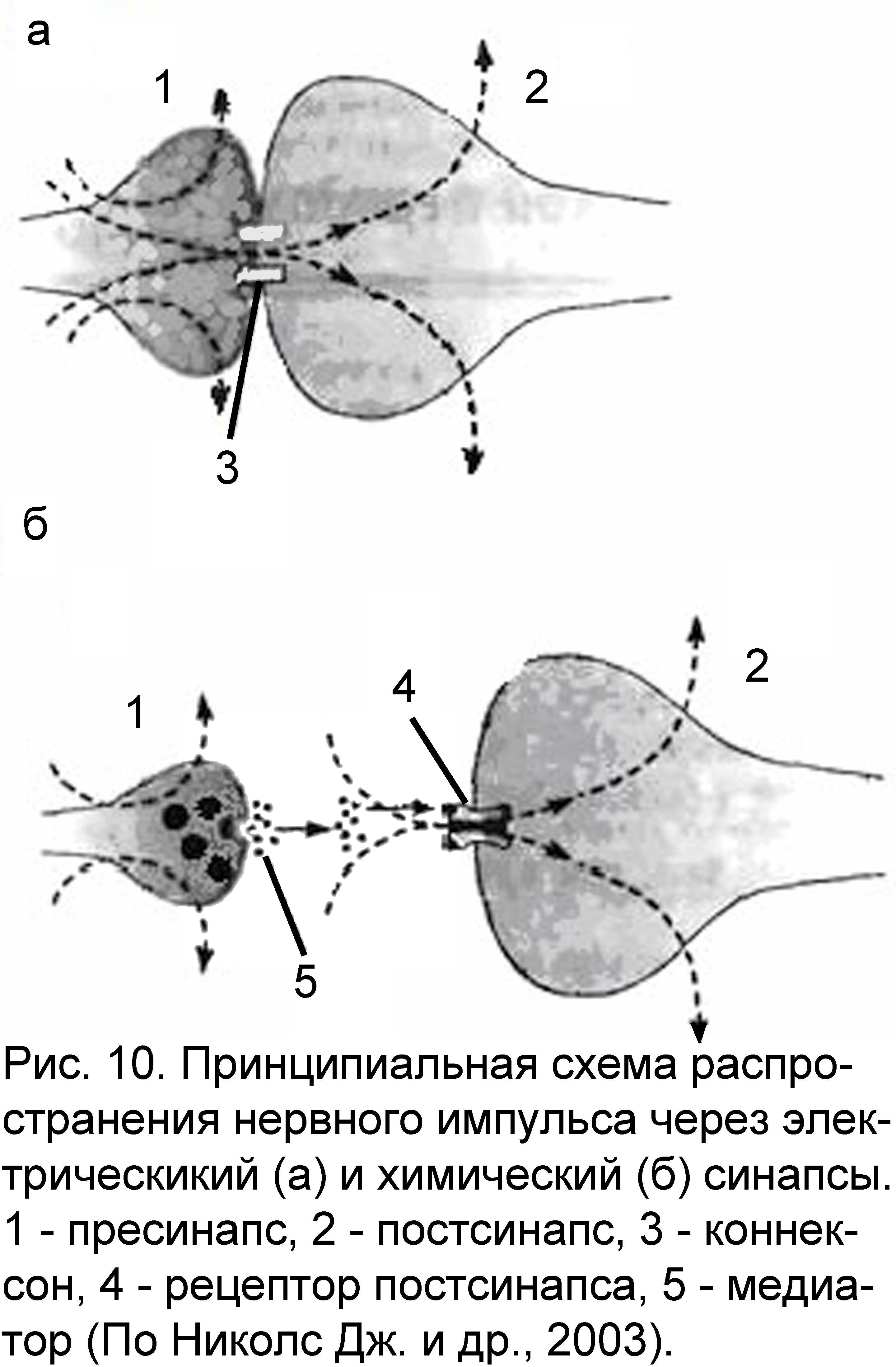
3.3. Химический синапс
В 1892 году Лэнгли предположил, что синаптическая передача в вегетативных ганглиях млекопитающих имеет химическую природу, а не электрическую. Примерно через 10 лет Эллиот обнаружил, что экстракт, полученный из надпочечников, — адреналин — производит на клетки такое же воздействие, как и стимуляция симпатических нервов. Это позволило предположить, что адреналин может секретироваться нейронами, и выделятся нервными окончаниями при проведении возбуждения. Однако лишь в 1921 году Леви поставил опыт, в котором была установлена химическая природа передачи в вегетативных синапсах между блуждающим нервом и сердцем. Он наполнял физиологическим раствором сосуды сердца лягушки и стимулировал блуждающий нерв, вызывая замедление сердцебиений. Когда жидкость из заторможенного стимуляцией сердца была перенесена на второе нестимулированное сердце, оно в результате такого воздействия также начинало биться медленнее. Было очевидно, что стимуляция блуждающего нерва вызывала освобождение в раствор тормозящего вещества. В последующих экспериментах Леви и его коллеги показали, что ацетилхолин полностью воспроизводил эффекты этого вещества.
Удивительно, что идея этого эксперимента пришла Леви во сне, он записал ее, но поутру не смог разобраться в том, что написал ночью. К счастью, сон повторился, и в этот раз Леви не стал рисковать: он помчался в лабораторию и поставил этот эксперимент. Вот как он вспоминает эту ночь: По серьезному размышлению, в холодных лучах утреннего света, я бы никогда не поставил этого эксперимента. То, что блуждающий нерв может освобождать тормозящее вещество, было очень маловероятно; еще менее вероятным было то, что химическое вещество, которое предположительно должно было действовать на коротком расстоянии между нервным окончанием и мышцей, освобождалось бы в таком большом количестве, что сохраняло бы эффективность после разведения в перфузирующей жидкости.
В начале 1930-х годов роль ацетилхолина в синаптической передаче в ганглиях вегетативной нервной системы была окончательно установлена Фельдбергом и его сотрудниками. Дейл, британский физиолог и фармаколог, установил роль ацетилхолина при передаче возбуждения в нервно--мышечном синапсе.
Принципиальным отличием химического синапса передача раздражения с пресинапса на постсинапс при помощи биологически активных веществ, называемых медиаторами (трансмиттерами). В связи с этим появляются и отличия в морфологии химического синапса (рис. 10б; приложение 2):
-
пресинаптическая область обязательно содержит везикулы (замкнутые в собственные мембраны полости), наполненные медиатором. Везикулы способны к перемещениям, когда они подходят к мембране нервного окончания, то их мембраны сливаются, а медиатор высыпается в синаптическую щель.
-
синаптическая щель имеется между пре- и постсинаптическими мембранами. Величина щели неодинакова в различных синапсах. Так, в приведенном ранее примере нервно-мышечного окончания, где медиатором является ацетилхолин, ширина синаптической щели достигает 30 нм. Пространство синаптической щели заполнено межклеточной жидкостью, а в нервно-мышечном окончании коллоидом (поэтому часто синаптическую щель в нервно-мышечном окончании рассматривают как базальную мембрану).
-
постсинаптическая мембрана содержит на своей поверхности рецепторы – белки, обладающие большим сродством к медиатору. Определенному типу медиатора соответствует определенный тип рецептора, например, в нервно-мышечном окончании – холинорецепторы, они распознают медиатор в окружающей среде и вступают с ним во взаимодействие. В результате этого взаимодействия происходят структурные изменения на постсинаптической мембране, в результате которых изменяется ионная проводимость через постсинапс. Это, в свою очередь, является причиной дальнейшего распространения нервного импульса (в случае, если медиатор оказывает тормозящий эффект, то развитию процессов торможения на постсинаптической мембране).
Кроме ацетилхолина в качестве медиатора могут выступать также:
-
аминокислоты: глутамат, глицин, аспартат
-
биогенные амины: дофамин, -аминомасляная кислота (ГАМК), норадреналин, адреналин, серотонин, гистамин
-
производные пурина: АТФ, АДФ, АМФ, аденозин
-
пептиды: энкефалины, эндорфины, вещество Р и многие другие.
Синапсы и нейроны, в соответствии с видом выделяемого медиатора называются по типу медиатора с добавлением окончания –эргический: холинэргический, адренэргический, ГАМКэргический, гистаминэргический и т.п.
Химические синапсы наиболее распространены в ЦНС позвоночных. В настоящее время известно, что нейрон может синтезировать и выделять несколько медиаторов (сосуществующие медиаторы). Нейроны также обладают нейромедиаторной пластичностью – способностью менять основной медиатор в процессе развития.