

8. Организация серого и белого вещества спинного мозга
8.1. Строение серого вещества спинного мозга
8.2. Основные проводящие пути спинного мозга
8.1. Строение серого вещества спинного мозга
Серое вещество спинного мозга условно может быть разделено на 10 слоев или пластин, в каждой из которых группируются нейроны определенного типа. Эта классификация предложена в 1952 году шведским гистологом Б. Рексердом и объединяет нейроны, близкие по своему функциональному назначению. Пластины обозначают римскими цифрами (рис. 30):
-
Пластина I - это тонкий слой нейронов, получающих ноцицептивный (болевая чувствительность) вход. Аксоны какой-то части этих нейронов переходят на противоположную сторону, аксоны другой части нейронов идут в составе спинно-таламического тракта ипсилатеральной (той же самой) стороны спинного мозга.
-
Пластины II и III составляют желатинозную субстанцию Роланда. Это небольшие нейроны, участвующие в контроле ноцицептивных сигналов, поступающих в спинной мозг. По своей функции нейроны желатинозной субстанции представляют собой тормозные интернейроны, тормозящие активность ноцицептивных и полимодальных (получающих другие виды рецепции) задних рогов серого вещества спинного мозга. Они образуют первый интеграционный центр болевой чувствительности.
-
Пластина IV состоит из нейронов, получающих входы периферических рецепторов, отвечающих на раздражения различных по своей природе стимулов. Аксоны этих нейронов поднимаются к латеральному шейному ядру в составе спинно-цервикального тракта. Вторая группа нейронов этой пластины формирует часть спинно-таламического тракта, которая проводит раздражения от любых раздражителей, кроме ноцицептивной.
-
Пластины V и VI – это шейка дорсального рога. Здесь заканчиваются волокна от сенсомоторной области больших полушарий и волокна, несущие проприоцептивную (регистрирующую движения мышц, суставов) чувствительность от туловища и конечностей. Одни аксоны нейронов этих пластин участвуют в образовании спинно-мозжечковых трактов, другие проецируются к ретикулярной формации ствола мозга и нижней оливе.
-
Пластина VII – это область окончания проприоспинальных и висцеральных волокон, а также афферентных и эфферентных связей с мозжечком и средним мозгом.
-
Пластина VIII – это бульбоспинальные (идущие из продолговатого мозга) и проприоспинальные связи (особенно контралатеральные и комиссуральные).
-
Пластина IX является первичной моторной областью. В ее медиальной части расположены мотонейроны, иннервирующие мускулатуру туловища, а в латеральной – мускулатуру конечностей
-
П
ластина X обрамляет спинномозговой канал и содержит нейроны, клетки глии и комиссуральные волокна.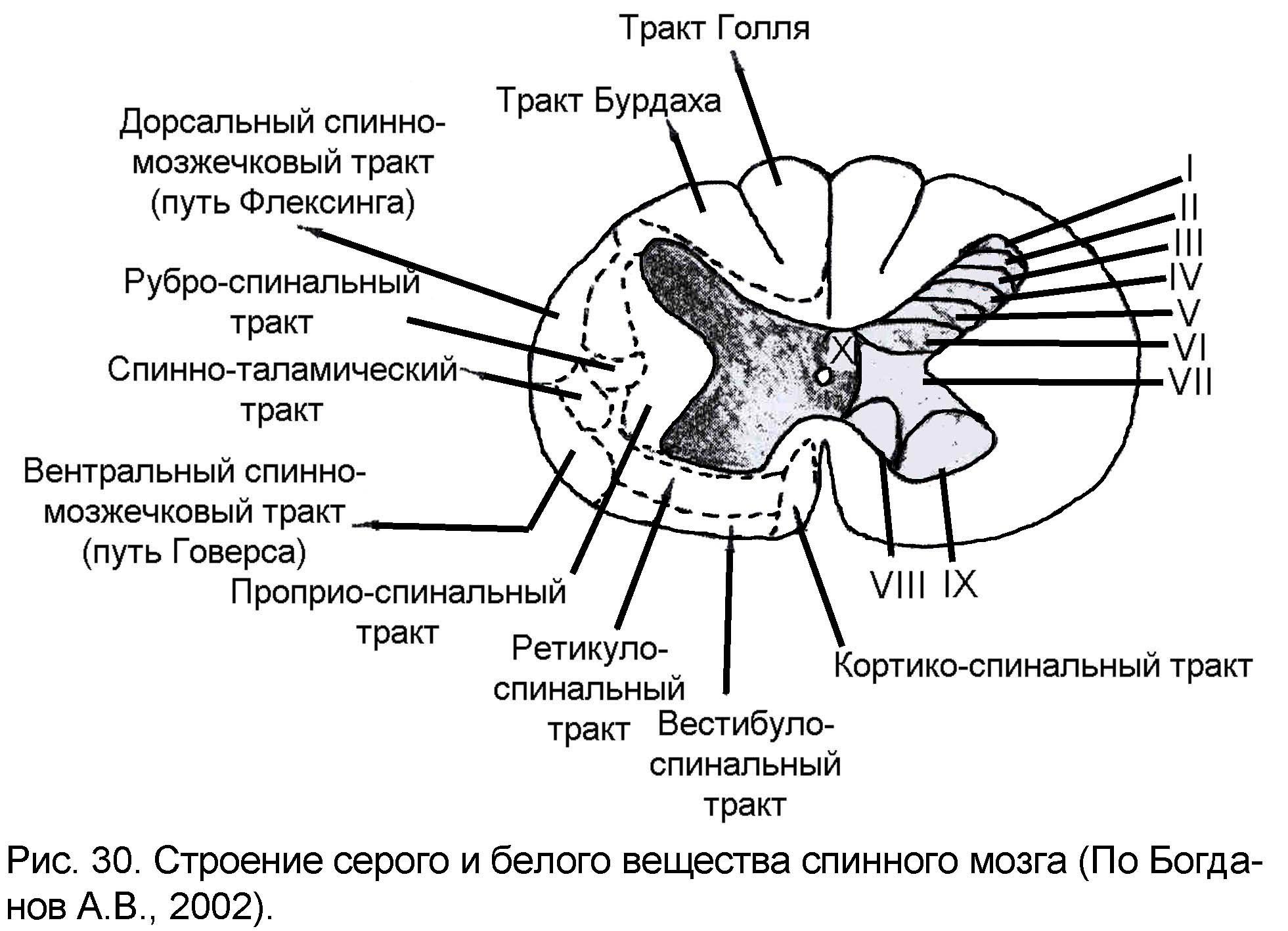
8.2. Основные проводящие пути спинного мозга
Не ставя перед собой задачи перечислить все проводящие пути ЦНС, рассмотрим основные принципы организации этих путей на примере наиболее важных из них (рис. 30). Проводящие пути в ЦНС делятся на:
-
восходящие - образуются аксонами клеток, тела которых расположены в сером веществе спинного мозга. Эти аксоны в составе белого вещества направляются к верхним отделам спинного мозга, стволу головного мозга и коре больших полушарий.
-
нисходящие – образуются аксонами клеток, тела которых расположены в различных ядрах головного мозга. Эти аксоны по белому веществу спускаются к различным спинальным сегментам, заходят в серое вещество и оставляют свои окончания на тех или иных его клетках.
-
Отдельную группу образуют проприоспинальные проводящие пути. Они могут быль как восходящими, так и нисходящими, но они не выходят за пределы спинного мозга. Пройдя несколько сегментов, они вновь возвращаются в серое вещество спинного мозга. Эти пути расположены в самой глубокой части латерального и вентрального канатиков, они связывают между собой различные нервные центры спинного мозга. Например, центры нижних и верхних конечностей.
Восходящие проводящие пути.
-
Тракты Голля (тонкий пучок) и Бурдаха (клиновидный пучок). Основные восходящие пути проходят через дорсальные канатики спинного мозга и представляют собой аксоны афферентных нейронов спинномозговых ганглиев. Они проходят по всему спинному мозгу и заканчиваются в области продолговатого мозга в ядрах дорсального канатика, которые называют ядрами Голля и Бурдаха. Поэтому они и именуются тракт Голля и тракт Бурдаха.
-
Первое звено нейронов:
-
Волокна, расположенные в канатике медиальнее несут к ядру Голля афферентные сигналы от нижней части тела, в основном от нижних конечностей.
-
Волокна, расположенные латеральнее, идут к ядру Бурдаха и передают афферентные сигналы от рецепторов верхней части туловища и передних конечностей.
-
-
Второе звено нейронов:
В свою очередь аксоны клеток ядер Голля и Бурдаха в стволе головного мозга перекрещиваются и в виде плотного пучка поднимаются до промежуточного мозга. Этот пучок волокон, образованный уже аксонами клеток ядер Голля и Бурдаха получил название медиальной петли.
-
Третье звено нейронов:
Клетки ядер промежуточного мозга дают аксоны, направляющиеся в кору больших полушарий.
Все остальные восходящие пути начинаются не от нейронов спинномозговых ганглиев, а от нейронов, расположенных в сером веществе спинного мозга. Следовательно, их волокна являются волокнами не первого, а второго порядка.
-
Первым звеном в этих путях также служат нейроны спинномозговых ганглиев, но в сером веществе они оставляют свои окончания на клетках как бы «второго звена».
Клетки этого «второго звена» посылают свои аксоны к ядрам ствола головного мозга и коре больших полушарий. Основная масса волокон этих путей проходит в латеральном канатике.
-
Спинно-таламические пути (вентральный и латеральный).
-
Второе звено нейронов:
Начинается в основании дорсального рога спинного мозга. Аксоны нейронов, образующих этот путь переходят на контралатеральную (противоположную) сторону, входят в белое вещество противоположного латерального или вентрального канатика и в нем поднимаются через весь спинной мозг и ствол головного мозга вплоть до ядер промежуточного мозга.
-
Третье звено нейронов:
Нейроны ядер промежуточного мозга переносят импульсацию в кору больших полушарий.
Все вышеописанные пути (Голля, Бурдаха и спинно-таламический) связывают рецептивные области каждой стороны тела с нейронами коры противоположного полушария.
-
Спинно-мозжечковые тракты. Еще два пути, проходящие в латеральных канатиках связывают спинной мозг с корой мозжечка.
Путь Флексинга – расположен дорсальнее и содержит волокна, непереходящие на противоположную сторону мозга. Это путь в спинном мозге начинается от нейронов ядра Кларка, аксоны которых достигают продолговатого мозга и поступают в мозжечок через нижнюю ножку мозжечка.
Путь Говерса – расположен вентральнее, содержит волокна, которые поднимаются вверх по латеральному канатику противоположной стороны тела, но в стволе мозга эти волокна снова перекрещиваются и входят в кору мозжечка с той стороны, на которой этот путь начинался. В спинном мозге начинается от ядер промежуточной зоны, аксоны вступают в мозжечок через верхнюю ножку мозжечка.
Если кора больших полушарий мозга всегда связана с афферентными волокнами противоположной стороны тела, то кора мозжечка получает волокна преимущественно от нейронных структур одноименной стороны.
Нисходящие проводящие пути. Волокна, идущие в нисходящем направлении, также подразделяются на несколько путей. В основе названия этих путей лежат названия тех отделов мозга, в которых они берут свое начало.
-
Кортико-спинальные (латеральный и вентральный) пути образованы аксонами пирамидных клеток нижних слоев моторной зоны коры больших полушарий. Часто эти пути называют пирамидными. Волокна проходят через белое вещество больших полушарий, основание ножек среднего мозга, по вентральным отделам Варолиева моста и продолговатого мозга в спинной мозг.
-
Латеральный путь перекрещивается в нижней части пирамид продолговатого мозга и заканчивается на нейронах основания заднего рога.
-
Вентральный путь пересекает пирамиды продолговатого мозга не перекрещиваясь. Перед вступлением в передний рог серого вещества соответствующего сегмента спинного мозга волокна этого пути переходят на противоположную сторону и заканчиваются на мотонейронах передних рогов контралатеральной стороны.
-
Таким образом, так или иначе, но двигательная область коры больших полушарий всегда оказывается связанной с нейронами противоположной стороны спинного мозга.
-
Рубро-спинальный путь – основной нисходящий путь среднего мозга, начинается в красном ядре. Аксоны нейронов красного ядра перекрещиваются сразу под ним и в составе белого вещества латерального канатика спускаются к сегментам спинного мозга, заканчиваясь на клетках промежуточной области серого вещества. Это связано с тем, что руброспинальная система наряду с пирамидной является основной системой контроля деятельности спинного мозга.
-
Текто-спинальный путь – Берет начало от нейронов четверохолмия среднего мозга и достигает мотонейронов передних рогов.
Проводящие пути, начинающиеся в продолговатом мозге:
-
Вестибуло-спинальный – начинается от вестибулярных ядер, главным образом от клеток ядра Дейтерса.
-
Ретикуло-спинальный – начинается от обширного скопления нервных клеток ретикулярной формации, занимающей центральную часть ствола мозга. Волокна каждого из этих путей заканчиваются на нейронах медиальной части переднего рога серого вещества спинного мозга. Основная часть окончаний располагаются на вставочных клетках.
-
Оливо-спинальный - образован аксонами клеток олив продолговатого мозга, заканчивается на мотонейронах передних рогов спинного мозга.