

2. 1952Г. Эксперимент Альфреда Херши и Марты Чейз.
Как известно, фаг Т2 является вирусом, инфицирующим бактерию E. coli. Фаговые частицы абсорбируются на наружной поверхности клетки, их материал проникает внутрь и примерно через 20 минут бактерия лизируется, освобождая большое количество фаговых частиц - потомков. В 1952 году Альфред Херши и Марта Чейз инфицировали бактерии фагами Т2, которые были мечены радиоактивными соединениями: ДНК - с помощью 32P (радиоактивный изотоп фосфора), т.к. ДНК содержит фосфор. Белковая часть фага - 35S (радиоактивный изотоп серы), т.к. некоторые АК содержат серу. После инфекции бактерии фагами, с помощью центрифугирования удалось выделить две фракции: пустые белковые оболочки фага и бактерии, инфицированных фаговой ДНК. Оказалось, что 80% метки 35S осталась в пустых фаговых оболочках, а 70% метки 32P - в инфицированных бактериях. Фаги-потомки получили только около 1% исходного белка, меченного 35S, однако они же обнаружили около 30% метки 32P.
Результаты этого эксперимента прямо показали, что радиоактивно меченая ДНК из родительских фагов попадает в клетки бактерий и обеспечивает размножение фагового потомства.
3. 1957г. Опыты Френкеля - Конрата
Френкель-Конрат работал с вирусом табачной мозаики (ВТМ). В этом вирусе содержится РНК, а не ДНК. Было известно, что разные штаммы вируса вызывают разную картину поражения листьев табака. После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком.
Следовательно, не только ДНК, но и РНК может служить носителем генетической информации.
32. Условия проведения гибридологического анализа.
Гибридологический анализ – это постановка системы скрещиваний, позволяющих выявить закономерности наследования признаков.
Условия проведения гибридологического анализа:
родительские особи должны быть одного вида;
родительские особи должны быть гомозиготными по изучаемым признакам;
изучаемый признак должен быть константным в ряду последующих поколений, полное доминирование;
родительские особи должны различаться по проявлению изучаемых признаков;
гены должны находиться в разных парах хромосом;
родительские особи скрещивают между собой один раз для получения гибридов первого поколения F1, которые затем скрещивают между собой для получения гибридов второго поколения F2 (индивидуальный анализ по потомству);
необходимо проведение строгого учета числа особей первого и второго поколения, имеющих изучаемый признак.
33. Взаимодействие генов.
Взаимодействие аллельных генов – различные типы доминирования. Изучение фенотипического расщепления – в соответствии с методикой гибридологического анализа, по законам Менделя, по схеме моногибридного скрещивания.
Полное - доминантная аллель полностью скрывает присутствие рецессивной (3:1)
Неполное - имеет место проявление промежуточного признака (1:2:1)
Кодоминирование - проявление у гетерозигот признаков, детерминируемых двумя аллелями (1:2:1)
Сверхдоминирование - доминантный ген в гетерозиготном состоянии имеет более сильное проявление, чем в гомозиготном. Например, у дрозофилы известна рецессивная летальная мутация, гетерозиготы обладают большей жизнеспособностью, чем гомозиготы (1:2:1)
(см. билет 2)
Взаимодействие неаллельных генов – проводится по схеме дигибридного скрещивания.
Взаимодействуют те гены, которые отвечают за проявление одного и того же признака. Изменяется соотношение классов в расщеплении (в F2).
Кооперация – появление новообразований при совместном действии двух доминантных неаллельных генов, когда в гомозиготном или гетерозиготном состоянии развивается новый признак, отсутствующий у родительских форм.
На примере попугайчиков:
P: AABB (зеленый) x aabb (белый)
F1: AaBb (зеленый)
F2: 9A_B_ (зеленый) 3A_bb (желтый) 3aaB_ (голубой) 1aabb (белый)
Комплементарность – неаллельные гены взаимно дополняют друг друга, при этом появляется новый признак.
Возможные расщепления:
-
По фенотипу
По генотипу
9:6:1
9A_B_ (3A_bb + 3aaB_) 1aabb
9:7
9A_B_ (3A_bb + 3aaB_ + 1aabb)
9:3:4
9A_B_ 3A_bb (3aaB_ + 1aabb)
9:3:3:1
9A_B_ 3A_bb 3aaB_ 1aabb
На примере собак:
P: AABB (черный) x aabb (белый)
F1: AaBb (черный)
F2: 9A_B_ (черный) 3aaB_ (коричневый) 4 (A_bb aabb) (белый)
Эпитаз – такой тип наследования, при котором действие одного гена подавляется действием другого неаллельного гена. Противоположно комплементарному действию генов. Различают доминантный (ген I – ингибитор – подавляет действие гена С, но на нашем примере будет стандартно А и В) и рецессивный (гены ii подавляют действие гена С) эпистаз.
Доминантный
Возможные расщепления:
-
По фенотипу
По генотипу
13:3
(9A_B_ +1aabb + 3aaB_) 3A_bb
12:3:1
(9A_B_ + 3aaB_) 3A_bb 1aabb
9:3:4
9A_B_ 3A_bb (3aaB_ + 1aabb)
На примере кур:
P: AABB (белый) x aabb (белый)
F1: AaBb (белый)
F2: 13 (A_B_ aaB_ aabb) (белый) 3A_bb (окрашенный)
Рецессивный
Расщепление: 9A_B_ 3aaB_ 4 (A_bb + aabb)
Полимерия – тип наследования, при котором на выраженность признака влияет несколько неаллельных генов. Чем больше в генотипе доминантных генов, обусловливающих какой – либо признак, тем ярче этот признак выражается. Различают кумулятивную (аддитивную, суммирующую) и некумулятивную (неаддитивную) полимерию.
Кумулятивная
На примере пигментации человека:
P: A1А1А2А2 (черный) x a1а1а2а2 (белый)
F1: A1а1А2а2 (коричневый)
Расщепление: 1:4:6:4:1 (15 более или менее окрашены, 1 (а1а1а2а2) белый)
Некумулятивная
На примере оперения кур:
P: A1А1А2А2 (оперенная) x a1а1а2а2 (неоперенная)
F1: A1а1А2а2 (оперенная)
Расщепление: 9:3:3:1 (15 оперенные, 1 (а1а1а2а2) нет)
Плейотропия – множественное действие генов - когда один ген определяет развитие нескольких свойств и признаков организма.
Например: У большинства растений с красными цветками (наследственный признак) в стеблях тоже имеется красный пигмент. У растений с белыми цветками, стебли чисто зеленые.
Например: у человека известен ген, определяющий одновременно развитие дефекта ногтей и коленной чашечки.
34. Гетерогенность популяций и методы определения гетерозиготных носителей.
Популяция – единица эволюции – группа особей одного вида, которые свободно скрещиваются и занимают определенный ареал.
Гетерогенность популяций генотипическая (гр. heterogenes — неоднородный по составу) — неоднородность составляющих популяцию особей, обусловленная разнообразием генотипов в пределах популяционного генофонда. Определяет эффективность отбора.
Гетерогенность популяций фенотипическая — неоднородность составляющих популяцию особей, обусловленная разнообразием фенотипов. Фенотипическая гетерогенность генотипически детерминирована.
Выявить гетерозиготных носителей можно с помощью идентификации фрагментов ДНК. Методы выявления:
Блот-гибридизация по Саузерну
Водородные связи между парами оснований в молекуле ДНК являются непрочными, поэтому цепи ДНК могут легко разделяться (денатурировать) и восстанавливаться при изменении температуры или солевых концентраций. При каждом цикле денатурации-восстановления образуется двунитевая структура - дуплекс. Ее устойчивость определяется степенью комлементарности нитей ДНК. Наиболее устойчивые структуры представлены полностью комплементарными нитями ДНК.
Это свойство широко используется в молекулярной биологии для поиска и идентификации необходимых последовательностей в огромных молекулах ДНК с помощью специфических зондов – небольших фрагментов однонитевых ДНК, имеющих радиоактивную метку.
Метод блот-гибридизации по Саузерну является наиболее эффективным методом выявления определенных последовательностей среди электрофоретически разделенных фрагментов ДНК. Этапы Саузерн-блоттинга:
Геномную ДНК разрезают одной или несколькими рестриктазами
Фрагменты ДНК разделяют в геле методом электрофореза
Затем ДНК подвергается денатурации и переносится с геля на фильтр или мембрану
Фиксированную на фильтре ДНК гибридизуют с радиоактивно меченым ДНК-зондом
Методом авторадиографии определяют положение искомого фрагмента ДНК на электрофореграмме. Выявляться будут те фрагменты ДНК, которые комплементарны специфическому зонду.
ПЦР (полимеразная цепная реакция)
ПЦР - метод амплификации ДНК in vitro, с помощью которого в течение нескольких часов можно выделить и размножить определённую последовательность ДНК в миллиарды раз. Метод ПЦР широко применяется в молекулярно-биологических исследованиях ДНК и РНК, а также в клинической практике для диагностики наследственных болезней, вирусных и протозойных инфекций.
Если в анализируемом образце присутствует искомая ДНК (ДНК-мишень), то в процессе реакции амплификации с ней происходит ряд событий, обеспечиваемых определенными температурными циклами.
Все этапы ПЦР проводят в специальном приборе – автоматическом термоциклере (амплификаторе), в котором программируется время проведения ПЦР и все условия ее проведения.
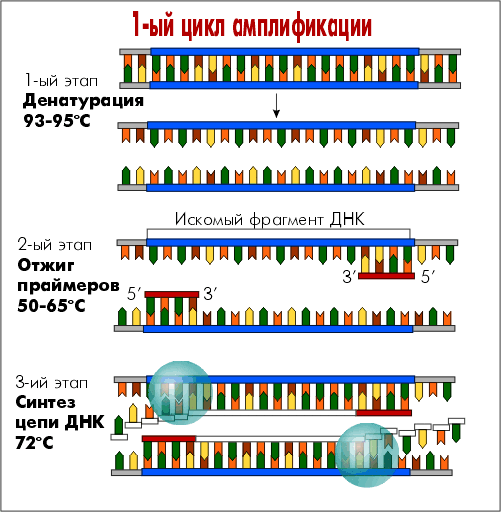
Каждый цикл амплификации состоит из трех этапов:
Денатурация. На первом этапе необходимо денатурировать ДНК, находящуюся в образце. Для этого реакционную смесь нагревают до 92-95°С, в результате чего двухцепочечные молекулы ДНК расплетаются с образованием двух одноцепочечных молекул.
Отжиг с праймерами. На втором этапе праймеры комплементарно присоединяются к одноцепочечной ДНК-мишени.
Элонгация (синтез новых цепей ДНК). После отжига праймеров Taq-полимераза начинает достраивание второй цепи ДНК, начиная с 3’-конца ДНК-праймера. На третьем этапе температуру в реакционной смеси доводят до оптимума работы Taq-полимеразы, и синтез второй цепи продолжается с максимальной эффективностью.
В дальнейшем этапы денатурации, отжига и элонгации многократно повторяются. На каждом цикле количество синтезированных копий фрагмента ДНК удваивается.
35. Секвенирование.
Секвенирование – это общее название методов, которые позволяют установить последовательность нуклеотидов в молекуле ДНК.
На практике наиболее широкое распространение получило секвенирование по классическому методу Сэнгера.
Изучаемый фрагмент ДНК (например, продукт ПЦР) играет роль матрицы, на которой при участии праймера, дезоксирибонуклеотид-трифосфатов и ДНК-полимеразы инициируется направленный комплементарный синтез новых молекул ДНК, осуществляемый одновременно в 4 параллельных реакциях. Смысл метода в том, что в каждую из реакций добавляется один из 4 химически измененных дезоксирибонуклеотид-трифосфатов (терминаторов). В качестве таких терминаторов выступают дидезоксирибонуклеотид-трифосфаты (ddGTP, ddATP, ddTTP и ddCTP), которые при встраивании в состав молекулы ДНК обрывают дальнейший синтез полинуклеотидной цепи. В результате этого в каждой из пробирок накапливается набор дочерних молекул ДНК различной длины, оканчивающихся строго на один и тот же нуклеотид.
После электрофореза продуктов этих реакций в 4 соседних дорожках проводится считывание информации от более коротких фрагментов к более длинным, при этом выявляемый пошаговый порядок удлинения цепей и отражает последовательность нуклеотидов в составе анализируемой ДНК-матрицы. Одномоментно может быть секвенирован участок ДНК, не превышающий 500 п.о., поэтому для исследования более длинных участков гена они должны быть амплифицировапы в виде совокупности и коротких перекрывающихся фрагментов, каждый из которых секвенируется отдельно. Обычно для обнаружения мутаций секвенируются отдельные экзоны изучаемого гена вместе с примыкающими к ним интронными последовательностями (областями сплайсинга).