

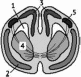
132
Подтип
Позвоночные. Надкласс Наземные
позвоночные
-
плечевая кость,
-
лопатка,
4-
вилочка (сросшиеся ключицы),
-
коракоиды,
-
ребро,
-
большая грудная мышца,
-
грудина с вентральным продольным
гребнем - килем,
-
вертикальная составляющая усилия,
-
горизонтальная составляющая усилия
Рис.
156. Поперечное сечение тела птицы через
пояс передних конечностей
Рис.
157. Череп птицы
1
- внешняя ноздря, 2 - височная дуга, 3 -
наружный слуховой проход, 4-глазница,
5 - мозговая капсула черепа, 6 - выступ
чешуйчатой кости, под которым расположено
среднее
ухо
Особенно
удлинены кости конечностей.
В
головном мозге (рис. 158) особенно крупный
мозжечок (7), площадь поверхности которого
увеличена за счёт развития рельефа,
и полушария конечного мозга (2). Смыкаясь
с мозжечком, они смещают в стороны
крупные зрительные доли среднего мозга
(6). Обонятельная доля (1) и обонятельная
(древняя) кора развиты слабо.
- Позвонок,


Класс
Птицы
Рис.
158. Головной мозг птицы (по
Ромер, Парсонс, 1992)
1
- обонятельная доля, 2 - полушарие
конечного мозга, 3 - промежуточный мозг,
4- гипофиз, 5 - зрительный нерв, 6 - зрительная
доля среднего мозга, 7 - мозжечок (задний
мозг), 8 - продолговатый мозг
Рис.
159. Поперечное сечение больших полушарий
рептилии (слева) и птицы (справа) (по
Ромер, Парсонс, 1992).
1
- латеральный желудочек, 2 - древняя
(обонятельная) кора, 3 - старая кора, 4 -
подкорковые ядра, 5 - новая кора
Для
птиц ещё больше, чем для рептилий,
характерно мощное развитие подкорковых
ядер больших полушарий (стриатума)
и развитие собственной коры на зрительных
долях среднего мозга (рис. 158, 159). Именно
эти два отдела ответственны за сложные
формы стереотипного поведения,
достигающие у птиц большого совершенства.
Способность к научению, особенно
развитая у вра- новых и крупных попугаев,
связана с участками, гомологичными


134 Подтип Позвоночные. Подкласс Наземные позвоночные
-
склера,
-
сосудистая оболочка,
-
сетчатка,
-
роговица,
-
костное склеральное кольцо,
-
связка,
-
ресничное тело,
-
радужина,
-
связка,
-
хрусталик,
-
оболочка хрусталика,
-
стекловидное тело,
-
гребень,
-
зрительный нерв
Рис.
160. Схема строения глаза птицы (по
Наумову, Карташову, 1979)
новой
коре млекопитающих. Связи стриатума
со зрительными долями проходят через
ядра промежуточного мозга. У птиц 12 пар
черепно-мозговых нервов.
Ведущий
анализатор птиц — зрительный (рис. 160)
. Глазные яблоки очень крупные, могут
в 2—3 раза превышать массу мозга и
составлять 0,5—5% массы тела. У разных
видов на один квадратный миллиметр
сетчатки приходится от 50 тыс. до 1 млн.
фоторецепторов — палочек
и колбочек;
цвета и их оттенки могут различать все
виды птиц.
В
области слепого
пятна
(зона выхода зрительного нерва)
расположен сосудистый вырост —
гребень,
участвующий в обмене веществ
стекловидного тела и внутренних слоёв
сетчатки. Аккомодация происходит
за счёт изменения формы хрусталика при
его одновременном перемещении и
некотором изменении кривизны
роговицы. Склера утолщённая, в ней
появляются костные
пластинки.
Хорошо развиты подвижные веки и
мигательная перепонка (третье веко),
двигающаяся непосредственно по
поверхности роговицы, очищая её.
Острота зрения птиц, по-видимому, в
несколько раз превосходит остроту
зрения у человека.
Слышат
все птицы очень хорошо, и звуковые
сигналы по своему значению для
организации поведения уступают, пожалуй,
только зрительным. Несмотря на отсутствие
ушной раковины и наличие только одной
слуховой косточки (стремени) больший-

Класс
Птицы
135
ство
видов воспринимает звуки в диапазоне
от 30 до 20 тыс. Гц (примерно как человек
с обострённым слухом). При этом некоторые
виды, вероятно, воспринимают и ультразвук
до 35—50 кГц, присутствующий в их голосе.
Многие виды способны с высокой точностью
(два-три угловых градуса, а у сов — один
градус) определять положение источника
даже крайне слабых звуковых сигналов.
Некоторые виды (гуахаро, саланганы)
используют звуковую эхолокацию для
ориентации в пещерах. Звуковые сигналы
используются птицами в самых разных
поведенческих сферах — общение родителей
с птенцами и партнёров друг с другом,
регуляция территориальных отношений,
сигналы тревоги и т. д.
Органы
осязания — механорецепторы — и
терморецепторы располагаются как
на оперённых, так и на лишённых перьев
участках кожи, на клюве, в ротовой
полости и на задних конечностях.
Обычно
считается, что вкусовая рецепция и
обоняние для большинства видов
играют подчинённую роль, но в последнее
время показано, что у хищников-падалыциков,
рыбоядных морских птиц, дятлов и многих
других групп обоняние достаточно
острое.
Важнейшее
преобразование в кровеносной
системе птиц
по сравнению с современными рептилиями
— возникновение четырёхкамерного
сердца, исключающего смешивание венозной
и артериальной крови (рис. 161).
Рис.
161. А - сердце рептилии; Б - сердце птицы
1
- сонные артерии, 2 - работающие дуги
аорты, 3 - артериальный проток, 4 - правая
лёгочная артерия, 5 - межжелудочковая
перегородка , 6 - редуцированная у птиц
левая дуга аорты, 7 - левая подключичная
артерия (для рептилий не показана)
136 Подтип Позвоночные. Подкласс Наземные позвоночные
-
бронх,
-
вторичные бронхи,
-
место соединения вторичного бронха с
воздушным мешком,
-
парабронхи,
-
бронхиоли
Рис.
162. Схема строения лёгкого птицы
(по
Наумову, Карташову, 1979)
После
полного разделения желудочка левая
дуга аорты, не связанная у предков птиц
с сонными и подключичными артериями,
редуцируется. В большой круг поступает
чисто артериальная кровь. Телесные
сосудистые сети у птиц, по сравнению с
рептилиями, существенно не меняются.
Сердце
у птиц крупное (1—2% от массы тела), его
работа очень интенсивна. У птиц среднего
размера (0,5 кг) в покое пульс 200—300, в
полёте —400—500 ударов в минуту; у мелких
птиц, соответственно —400—600 и 1000 ударов
в минуту. Среднее артериальное
давление у птиц — 120—200 мм рт. ст. (для
сравнения: у млекопитающих — 70—160 мм,
а у рептилий — только 30—50 мм). Количество
эритроцитов и содержание в них гемоглобина
у птиц и млекопитающих примерно равны
и заметно выше, чем у рептилий. Это
позволяет теплокровным повысить
кислородную ёмкость крови в 2—4 раза
по сравнению с пресмыкающимися.
Система
органов дыхания птиц
уникальна и по возможностям газообмена
не имеет себе равных среди позвоночных.
Верхние дыхательные пути и трахея
построены без принципиальных отличий
от рептилий, но в районе бифуркации
(разделения на два бронха) трахеи
находится характерная только для птиц
нижняя
гортань,
которая (в отличие от обычной — верхней)
у птиц является звукопорождающим
органом.
Лёгкие,
приросшие у птиц к рёбрам, невелики и
малорастяжимы. Войдя в лёгкое, бронх
делится на 15—20 вторичных бронхов, часть
из которых связана с тонкостенными
выростами — воздушными
мешками.
Между собой вторичные бронхи связаны
тонки-

Класс
Птицы
137
-
трахея,
-
лёгкое,
-
шейный мешок,
4-
межключичный мешок,
5,
6, 7 и 8 - выросты межключич- ного мешка,
переднегрудной
мешок,
-
заднегрудной мешок,
-
брюшной воздушный мешок
Рис.
163. Воздушные мешки птицы, вид с брюшной
стороны
(по
Наумову, Карташову, 1979)
ми
парабронхами,
от которых радиально отходит множество
тончайших бронхиол,
густо оплетённых капиллярами (рис.
162).
Воздух
при вдохе, пройдя трахею (рис. 163) и бронх,
попадает не в лёгкое, а всасывается в
задние воздушные мешки, а затем, сквозь
лёгкое, проходит в передние мешки,
откуда при выдохе выталкивается в
трахею. Таким образом, воздух сквозь
лёгкое идёт всегда в одном направлении
(рис. 164).
Перепады
давления между задними и передними
воздушными мешками возникают при
движении грудины, покачивающей-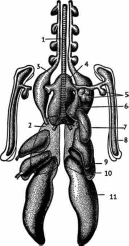
138 Подтип Позвоночные. Подкласс Наземные позвоночные
вдох
выдох
Рис.
164. Схема движения воздуха в дыхательной
системе птиц
(из
Наумова, Карташова, 1979 по Шмидту -
Нильсону)
Сплошные
стрелки показывают направление тока
воздуха, пунктирные стрелки - расширение
или сужение воздушных мешков; * - место
сужения стенок, прекращающее здесь ток
воздуха в данной фазе.
ся
поперёк оси, образованной головками
коракоидов, при работе полётной
мускулатуры (рис. 165).
На
земле вентиляция осуществляется, как
и у рептилий, за счёт работы межрёберной
мускулатуры. Площадь газообменного
эпителия в лёгком птиц соизмерима с
его площадью у млекопитающих и
намного превосходит внутреннюю площадь
лёгкого у рептилий. Однако эффективный
газообмен связан не только с этим, а в
первую очередь с системой противотока,
характерной для лёгкого птиц (рис.
166). Воздух и кровь проходят сквозь
лёгкое во встречных направлениях, что
позволяет поддерживать высокую разницу
концентрации кислорода по всей длине
капил-
