

V стадия полового развития
Наступление этой стадии у девочек в большинстве случаев происходит в 15—16 лет, реже — в 13—14, у мальчиков в основном в 16—17, иногда в 18 лет, т.е. уже не в подростковом, а в юношеском возрасте. Темп полового созревания во многом определяется наследственностью, в частности типом телосложения. Так, у представителей астенического и астеноторокального типов половое созревание происходит сравнительно медленно, а половая зрелость наступает в возрасте 16—17 лет. У подростков мышечного и дигестивного типов половое созревание завершается в среднем на 1,5—2 года раньше. Люди с разным типом телосложения не только различаются внешне, их физиологические системы также функционируют по-разному. Поэтому определение типа телосложения имеет большое значение для индивидуальной характеристики подростка и юноши.
Физическое развитие. Обычно считается, что на завершающих этапах полового созревания, к началу юношеского возраста, основные физиологические системы уже созрели. Однако данные, полученные в последнее время, свидетельствуют, что это далеко не так. Результаты антропометрических измерений по 26 показателям свидетельствуют о том, что в этом возрасте продолжаются изменения в соотношении различных частей тела (его длины, ширины плеч, ширины тела). Эти преобразования тесно связаны с изменениями гормонального фона. Хотя внешние признаки указывают на завершение полового созревания, однако существенные гормональные перестройки, связанные с половым созреванием, еще продолжаются. В юношеском возрасте устанавливаются новые взаимоотношения между отдельными звеньями эндокринной системы, обеспечивающие совершенствование регуляторных процессов и их экономизацию.
Вегетативные функции. Важнейшая физиологическая особенность этого этапа развития — резкое расширение резервных возможностей органов и систем вегетативного обеспечения. Это расширение идет двумя путями: за счет развития функциональных возможностей периферических органов (к 17 годам системы вегетативного обеспечения выходят по большинству показателей на зрелый уровень функционирования) и за счет совершенствования центральных механизмов управления. В качестве примера рассмотрим процессы, связанные с мышечной деятельностью и ее вегетативным обеспечением. Периферическим звеном здесь являются скелетные мышцы. Хотя их строение в основном определено уже на предыдущем этапе развития, когда закладываются соотношения волокон различных типов (формируется метаболический и функциональный профиль), созревание волокон продолжается и в юношеском возрасте. Это проявляется в увеличении размеров (поперечника) волокон, их гликогеновых и жировых запасов. Все это существенно раздвигает рамки функциональных возможностей мышц. Одновременно повышается качество регулирования. На смену неэкономичным реакциям приходят очень специфичные, оптимальные по величине изменения именно тех показателей деятельности вегетативных систем, от которых зависит успешное решение двигательной задачи. Экономизация реакций проявляется, например, в резком снижении чувствительности дыхательного центра: одно и то же увеличение напряжения углекислого газа в крови приводит в юношеском возрасте к значительно меньшим сдвигам в легочной вентиляции, чем у детей и подростков. Более экономичными становятся терморегуляторные реакции, как при изменении температуры окружающей среды, так и во время мышечной деятельности. При мышечной работе, умеренной и большой мощности, между системами кровообращения, дыхания, терморегуляции и другими вегетативными системами отрабатывается тонкое взаимодействие, основанное на установлении минимального числа необходимых взаимосвязей, и лишь при утомлении оптимизация этих взаимодействий нарушается.
Работоспособность. В результате всех этих изменений, затрагивающих периферические и центральные физиологические механизмы, резко возрастает надежность функционирования всех систем организма. Это значительно повышает работоспособность. Юноша может выполнить объем работы (при сопоставимой ее интенсивности) примерно в 20—30 раз больший, чем ребенок 9—10 лет. Такое колоссальное увеличение работоспособности — результат не только увеличения размеров тела и структурных перестроек, но и оптимизации регуляторных процессов: совершенствуется как гормональная, так и нервная регуляция.
Мышцы
 Сократительная способность мышц
обеспечивается за счет структурных
элементов мышечной ткани, которые
называются миофибриллами. От строения
миофибрилл зависит поперечная
исчерченность мышц — чередование
светлых и темных полос, а толщина мышечных
волокон зависит от количества и
поперечного сечения миофибрилл.
Миофибриллы состоят из повторяющихся
блоков, называемых саркомерами, и
располагаются параллельно длинной оси
мышечного волокна. Скелетные мышцы
образованы поперечно-полосатой мышечной
тканью, основным структурным элемен-том
которой являются мышечные волокна —
миосимпласт. Волокнами они названы
из-за большой разницы между поперечным
сечением (от 0,05 до 0,11 мм) и длиной мышечной
клетки (до 15 см). Длина волокон зависит
от длины и строения самой мышцы.
Сократительная способность мышц
обеспечивается за счет структурных
элементов мышечной ткани, которые
называются миофибриллами. От строения
миофибрилл зависит поперечная
исчерченность мышц — чередование
светлых и темных полос, а толщина мышечных
волокон зависит от количества и
поперечного сечения миофибрилл.
Миофибриллы состоят из повторяющихся
блоков, называемых саркомерами, и
располагаются параллельно длинной оси
мышечного волокна. Скелетные мышцы
образованы поперечно-полосатой мышечной
тканью, основным структурным элемен-том
которой являются мышечные волокна —
миосимпласт. Волокнами они названы
из-за большой разницы между поперечным
сечением (от 0,05 до 0,11 мм) и длиной мышечной
клетки (до 15 см). Длина волокон зависит
от длины и строения самой мышцы.
Мышечные волокна распо-лагаются параллельными рядами и соединяются в пучки, которые образуют саму скелетную мыш-цу. Небольшие мышечные пучки покрыты тонкой соединительной тканью — эндомизием, крупные — перимизием, а всю мышцу в целом покрывает плотная соединительная ткань — эпимизий. На концах мышцы переходят в сухожилия, которые обладают большей эластичностью, упругостью и прочностью, чем мышечные волокна, что позволяет избегать разрывов мышц и их отрывов от костей при интенсивной внутренней нагрузке или сильном внешнем воздействии.
Волокна составляют примерно 86–90% от общей массы мышцы. Остальные проценты делят между собой кровеносные сосуды и нервы, обеспечивающие трофику (жизнедеятельность), питание и работоспособность мышц.
Скелетные мышцы. Мышцы — одна из четырех наиболее важных тканей в организме среди таких, как нервная, соединительная и эпителиальная. Существует три вида мышц: сердечная мышца; гладкая мускулатура, локализованная в стенках кровеносных сосудов, дыхательных путей, кишечника и мочевого пузыря; скелетная (поперечно-полосатая) мускулатура. Только скелетные мышцы находятся под прямым волевым контролем, обеспечивающим как перемещение частей тела, так и поддержание позы. Независимо от вида мышечной деятельности функцией мышцы является проявление ею силы сокращения и использование для этого необходимой энергии.
Структура, иннервация и кровоснабжение. Скелетные мышцы отделены от соединительной ткани (перимизиума, или фасции) мембранами. Соединительная ткань проникает и в полость самой мышцы, уменьшаясь при этом по толщине (в данном случае соединительную ткань называют эндомизиумом), в связи с чем, происходит разделение мышцы на все более и более мелкие компартменты. Наименьшими из них являются пучки, включающие большое количество мышечных волокон, прикрепленных друг к другу и к эндомизиуму соединительной тканью. К обоим концам мышцы эта ткань переходит в сухожилия, прикрепляемые к костям скелета. Сухожилия представляют собой плотно упакованные коллагеновые волокна, образующие соединения между мышцами и костями. Внешняя коллагеновая мембрана живой кости (периостеум) переходит в мембрану волокон сухожилия. Отдельные мышцы состоят из многих мышечных волокон, параллельно соединенных между собой, которые могут (но не обязательно) протягиваться по всей ее длине. Внутри мышцы соединительная ткань также покрывает крупные кровеносные сосуды и нервы. Почти все мышечные волокна иннервируются только одним нервным окончанием, расположенным посредине волокна. Особый участок поверхности мышечного волокна, прилегающий к нервному окончанию в области синапса, называется концевой пластинкой. Нейропередатчиком, который, освобождаясь из нервного окончания, инициирует развитие мышцей напряжения, является ацетил-холин.
Кровеносные сосуды ориентированы в основном параллельно мышечным волокнам, а многочисленные капилляры направлены в пространство между ними. Гладкая мускулатура кровеносных сосудов сокращается или расслабляется под влиянием нервных, гормональных и локальных воздействий, что обеспечивает регуляцию тока крови. Во время выполнения
физических упражнений кровоснабжение работающих мышц может возрасти в 100 раз по сравнению с уровнем покоя.
Мышечные клетки представляют собой длинные многоядерные волокна. Их длина может варьировать от нескольких миллиметров до 30 см, а диаметр — в пределах 10-100 мкм. Каждое мышечное волокно окружено гомогенной мембраной — сарколеммой, содержащей на своей наружной поверхности коллагеновые волокна, с помощью которых прикрепляется к внутримышечным элементам соединительной ткани. Внутренняя часть сарколеммы обладает свойством, благодаря которому питательные вещества и шлаки могут проходить через мембрану, в результате в мышечном волокне может возникать и распространяться потенциал действия. Инвагинированные участки сарколеммы называются «Т-трубочки», по которым потенциал действия распространяется внутрь мышечного волокна.
Ультраструктура мышечных волокон. Внутренность мышечного волокна заполнена саркоплазмой (цитоплазмой мышечной клетки), представляющей собой вязкую жидкость, содержащую ядра, митохондрии, миоглобин и около 500 нитевидных миофибрилл толщиной 1-3 мкм каждая и располагающихся от одного конца мышечного волокна к другому. Красный цвет саркоплазмы обусловлен присутствием в ней миоглобина — внутриклеточного дыхательного пигмента, благодаря которому создается запас кислорода. Миофибриллы находятся в окружении более совершенной структуры — эндоплазматического ретикулума, называемого саркоплазматическим ретикулумом (СР), который принимает участие в процессах роста, развития и восстановления мышцы. Взаимосвязанные мембранные трубочки находятся в узком пространстве между миофибриллами, окружая их и располагаясь параллельно с ними. Под микроскопом мышечные волокна обнаруживают поперечно-полосатую исчерченность, связанную с уникальной организацией миофибрилл. Темные диски А сменяются светлыми дисками I по всей длине каждой миофибриллы, которые являются сократительными элементами. Как показано, посредине A-диска имеется более светлая полоска, называемая зоной Н, которая видима только в расслабленном состоянии мышечного волокна. Сама зона H разделяется темной линией М.. Диски I также разделяются посредине темной зоной, названной Z-пластинками. Саркомер определяется как участок, расположенный между двумя последовательными Z-пластинками и является наименьшим сократительным элементом мышечного волокна. Каждая миофибрилла представляет собой цепь саркомеров. На молекулярном уровне можно обнаружить, что в паттерне миофибриллярных участков внутри каждого саркомера имеется два типа белковых филаментов (миофиламентов). Тонкие миофиламенты включают белки — актин, тропомиозин и тропонин; они простираются через диск I и частично проникают в диск А; толстые миофиламенты содержат белок миозин и располагаются в A-диске. Линия Z представляет собой белковый слой в форме диска и служит участком, к которому прикреплены тонкие миофиламенты.
Зона Н является областью, в которой толстые филаменты не перекрываются тонкими, поэтому под микроскопом она выглядит более светлой по сравнению с диском А, когда миофибрилла находится в расслабленном состоянии. М-линия посредине зоны Н выглядит несколько темнее из-за тонких прядей, соединяющих воедино примыкающие толстые миофиламенты.








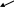













П оследовательные
поперечные разрезы миофибриллы
обнаруживают,
что в областях, где толстые и тонкие
миофиламенты перекрываются,
каждый толстый миофиламент окружен
гексагональным расположением
шести тонких миофиламентов, а каждый
тонкий миофиламент
расположен внутри треугольника,
образованного тремя толстыми
миофиламентами.
оследовательные
поперечные разрезы миофибриллы
обнаруживают,
что в областях, где толстые и тонкие
миофиламенты перекрываются,
каждый толстый миофиламент окружен
гексагональным расположением
шести тонких миофиламентов, а каждый
тонкий миофиламент
расположен внутри треугольника,
образованного тремя толстыми
миофиламентами.
В присутствии достаточного количества кальция и аденозинтрифосфата (АТФ) филаменты взаимодействуют между собой, образуя актомиозин, и сокращаются, скользя, относительно друг друга. Электрическое возбуждение, распространяясь в виде потенциала действия вдоль саркомера и к Т трубочкам, вызывает освобождение кальция из саркоплазматического ретикулума и выход его в саркоплазму с последующей активацией сокращением филаментов. Возбуждение инициируется достижением нервного импульса мышечной мембраны через двигательную концевую пластинку.
М олекулярный
состав миофиламентов.
Каждый толстый
миофиламент содержит около 200 молекул
миозина, из
которых каждая, в свою очередь, имеет
стержнеподобный
«хвост» с двумя
глобулярными структурами
на конце, которые обладают
АТФ-азной активностью. Миозиновые
головки взаимодействуют
со специфическими участками, расположенными
на тонких миофиламентах
с образованием, так называемых поперечных
мостиков, и
генерируют развитие напряжения,
приводящего к сокращению мышцы. В
толстом филаменте молекулы миозина
связаны вместе таким образом,
что их «хвосты» образуют центральную
часть филамента, а их глобулярные
структуры обращены наружной поверхностью
в направлениях,
противоположных друг другу.
Благодаря этому каждый толстый
филамент имеет относительно гладкую
центральную секцию с двумя
концами, усеянными множеством подвижных
головок миозина. головок миозина.
олекулярный
состав миофиламентов.
Каждый толстый
миофиламент содержит около 200 молекул
миозина, из
которых каждая, в свою очередь, имеет
стержнеподобный
«хвост» с двумя
глобулярными структурами
на конце, которые обладают
АТФ-азной активностью. Миозиновые
головки взаимодействуют
со специфическими участками, расположенными
на тонких миофиламентах
с образованием, так называемых поперечных
мостиков, и
генерируют развитие напряжения,
приводящего к сокращению мышцы. В
толстом филаменте молекулы миозина
связаны вместе таким образом,
что их «хвосты» образуют центральную
часть филамента, а их глобулярные
структуры обращены наружной поверхностью
в направлениях,
противоположных друг другу.
Благодаря этому каждый толстый
филамент имеет относительно гладкую
центральную секцию с двумя
концами, усеянными множеством подвижных
головок миозина. головок миозина.
Тонкие миофиламенты включают в себя актин и несколько регуляторных белков. Глобулярные (G) мономеры актина полимеризованы внутри длинных участков, называемых фибриллярным (F) актином. Две актиновые нити, сплетаясь вместе, образуют основу каждого тонкого филамента. Далее хвостоподобные молекулы тропомиозина обвивают цепочки F-актина, чем помогают миофиламенту стать прочнее.
Другим важным белком, присутствующим в тонких филаментах, является тропонин, который содержит три субъединицы. Одна из них, тропонин I, связана с актином; другая, тропонин Т, связана с тропомиозином, а третья, тропонин С, может быть связана с ионами кальция.
Механизм проявления силы мышечного сокращения. При сокращении мышечного волокна его саркомеры укорачиваются, Н-зоны исчезают, и расстояние между ближайшими линиями Z уменьшается. Сами же филаменты не изменяют своей длины. Скольжение миофиламентов начинается, когда головки миозина образуют поперечные мостики, прикрепляясь к активным участкам актиновых субъединиц тонких филаментов. Каждое образование и разъединение поперечных мостиков во время сокращения происходит несколько раз храповикоподобным образом, в результате чего тонкие филаменты перемещаются в направлении центра саркомера. В результате такого процесса длина всего мышечного волокна становится короче.
Для образования миозиновых поперечных мостиков требуется присутствие ионов кальция. В расслабленном мышечном волокне кальций находится в саркоплазматическом ретикулуме и, в отсутствие этих ионов, связывающие участки миозина физически блокированы тропомиозином. Ионы кальция при освобождении из саркоплазматического ретикулума (вследствие возбуждения нервным импульсом) связываются с тропонином С и тем самым изменяют его конформацию, что приводит к физическому перемещению тропомиозина от связывающих участков миозина на расположенную внутри цепь актина.
 Активированные,
или «склеванные» головки теперь
присоединяются к актину и, в данном
случае головная часть молекулы изменяет
свою активную
конфигурацию до такого состояния своей
связывающей поверхности, которая
обеспечивает ее перемещение на тонкий
филамент путем
скольжения по направлению к средине
саркомера. Представленная
схема отражает развитие цикла поперечных
мостиков и стимуляцию его аденозиндифосфатом
(АДФ) и неорганическим фосфатом
(Фн).
Как только новая молекула АТФ присоединяется
к участку миозиновой
головки, обладающему АТФ-азной активностью,
миозиновые мостики
отсоединяются от актина.
Активированные,
или «склеванные» головки теперь
присоединяются к актину и, в данном
случае головная часть молекулы изменяет
свою активную
конфигурацию до такого состояния своей
связывающей поверхности, которая
обеспечивает ее перемещение на тонкий
филамент путем
скольжения по направлению к средине
саркомера. Представленная
схема отражает развитие цикла поперечных
мостиков и стимуляцию его аденозиндифосфатом
(АДФ) и неорганическим фосфатом
(Фн).
Как только новая молекула АТФ присоединяется
к участку миозиновой
головки, обладающему АТФ-азной активностью,
миозиновые мостики
отсоединяются от актина.
Гидролиз АТФ до АДФ и Фн, происходящий при участии АТФ-азы, обеспечивает энергией, необходимой для возвращения миозина в его активированное, «склевывающее» состояние, позволяющее при наличии энергетического потенциала осуществлять последовательную работу поперечных мостиков.
До тех пор, пока миозин находится в активном состоянии, АДФ и Фн остаются связанными с миозиновой головкой. Когда же последняя обретает способность связываться с другим участком актина, расположенного последовательно в тонком миофи-ламенте, цикл связывания, развития напряжения и разъединения попе-речных мостиков, как и активация миозина, повторяются.
Взаимное скольжение филаментов описанным способом может продолжаться до тех пор, пока в саркоплазме присутствуют ионы кальция (концентрация должна быть не менее 10 ммоль). Перемещение и обратный захват АТФ-зависимой кальциевой помпой саркоплазматического ретикулума кальция восстанавливает тропомиозиновое угнетение образования поперечных мостиков и расслабление мышечных волокон.
Р егуляция
силы мышечных сокращений. Проявление
мышечным
волокном силы сокращения является
результатом нервной импульсации
от двигательного нерва, распространяющейся
в виде потенциала действия по сарколемме.
Потенциал действия, достигнув двигательной
концевой
пластинки, вызывает освобождение
нейромедиатора ацетилхолина,
который перемещается специальным
синапсом между нервным окончанием
и мышечным волокном (нейромышечным
соединением) и взаимодействует
с ацетилхолиновыми рецепторами,
расположенными на
сарколемме. Это приводит к открытию
натриевых каналов, в результате чего
поток ионов натрия внутрь мышечного
волокна снижает градиент их
концентрации. Происходит деполяризация
мембраны и, как следствие,
генерация потенциала действия, который,
распространяясь по
сарколемме мышечного волокна в обе
стороны и внутрь Т-трубочек, вызывает
полную активацию мышечного волокна.
Передача потенциала
действия к участкам, где Т-трубочки
примыкают к саркоплазматическому
ретикулуму, обусловливает последующее
освобождение
из него кальция (кальциевые каналы
временно открываются) и его свободная
концентрация в саркоплазме становится
более 10 ммоль, что сопровождается
образованием поперечных мостиков, как
описано выше.
Последующая активация кальциевой помпы
приводит к возвращению
кальция в саркоплазматический ретикулум
(обычно это происходит в
пределах 30 мс), а ингибирование тропомиозина
восстанавливается, если
концентрация кальция в саркоплазме
становится очень низкой. Такая
последовательность процессов повторяется,
когда другие импульсы, поступающие
от двигательного нерва, достигают
двигательной концевой
пластинки. Если же частота импульсации
высокая, то ионы кальция продолжают
высвобождаться из саркоплазматического
ретикулума и концентрация
кальция в саркоплазме, окружающей
миофиламенты, значительно
возрастает. В таком случае, мышечные
волокна, между последующими
стимулами полностью не расслабляются.
егуляция
силы мышечных сокращений. Проявление
мышечным
волокном силы сокращения является
результатом нервной импульсации
от двигательного нерва, распространяющейся
в виде потенциала действия по сарколемме.
Потенциал действия, достигнув двигательной
концевой
пластинки, вызывает освобождение
нейромедиатора ацетилхолина,
который перемещается специальным
синапсом между нервным окончанием
и мышечным волокном (нейромышечным
соединением) и взаимодействует
с ацетилхолиновыми рецепторами,
расположенными на
сарколемме. Это приводит к открытию
натриевых каналов, в результате чего
поток ионов натрия внутрь мышечного
волокна снижает градиент их
концентрации. Происходит деполяризация
мембраны и, как следствие,
генерация потенциала действия, который,
распространяясь по
сарколемме мышечного волокна в обе
стороны и внутрь Т-трубочек, вызывает
полную активацию мышечного волокна.
Передача потенциала
действия к участкам, где Т-трубочки
примыкают к саркоплазматическому
ретикулуму, обусловливает последующее
освобождение
из него кальция (кальциевые каналы
временно открываются) и его свободная
концентрация в саркоплазме становится
более 10 ммоль, что сопровождается
образованием поперечных мостиков, как
описано выше.
Последующая активация кальциевой помпы
приводит к возвращению
кальция в саркоплазматический ретикулум
(обычно это происходит в
пределах 30 мс), а ингибирование тропомиозина
восстанавливается, если
концентрация кальция в саркоплазме
становится очень низкой. Такая
последовательность процессов повторяется,
когда другие импульсы, поступающие
от двигательного нерва, достигают
двигательной концевой
пластинки. Если же частота импульсации
высокая, то ионы кальция продолжают
высвобождаться из саркоплазматического
ретикулума и концентрация
кальция в саркоплазме, окружающей
миофиламенты, значительно
возрастает. В таком случае, мышечные
волокна, между последующими
стимулами полностью не расслабляются.
И развиваемое мышцей напряжение будет более сильным и непрерывным (вплоть до максимального), до тех пор, пока не прекратится нервная импульсация.
Нервная импульсация. Группы мышечных волокон (одного и того же типа) объединены в мышце их взаимосвязью с одним и тем же двигательным нейроном. Каждая такая группа называется двигательной единицей. Двигательные единицы различаются по количеству содержащихся в них волокон: одни из них содержат около 50 волокон, другие — до 1700. Мышцы, выполняющие тонкие градуальные движения (например, мышцы глаз и рук), содержат незначительное количество двигательных единиц; мышцы же, имеющие большую массу и выполняющие грубые движения (например, мышцы ног), имеют большее количество двигательных единиц.
Импульс, распространяющийся по аксону двигательного нерва, вызывает деполяризацию его концевых пластинок. Все мышечные волокна, принадлежащие одной и той же двигательной единице, будут либо не отвечать (а), либо отвечать (б) проведением потенциала действия по мышечным волокнам, что сопровождается одновременной активацией всех волокон.
Ответы мышечных волокон на одиночные импульсы (достаточной силы) с одиночным сокращением и расслаблением продолжаются около 30 мс. Повторяющиеся сверхпороговые стимулы при сближении их вплотную вызывают суммацию, и когда частота стимуляции составляет более 60 в секунду (т. е. 60 Гц), происходит слияние одиночных сокращений (тетанус), при котором мышечное волокно развивает более высокое напряжение.
Обычно частота стимуляции находится в пределах от 5 Гц, при которой развивается низкое напряжение, до 70 Гц. В последнем случае сила сокращения мышечных волокон высокая. Так, сила сокращений целостной мышцы может повышаться в результате повышения частоты стимуляции активных двигательных единиц (а), а также вовлечением (рекрутированием) и увеличением их количества (б).
При выполнении физических упражнений с фиксированной мощностью по мере развития утомления некоторые двигательные единицы перестают участвовать в развиваемом мышцей напряжении, однако их вклад в генерацию силы будет немедленно возмещаться другими двигательными единицами до тех пор, пока все из них не будут рекрутированы. При выполнении физических упражнений максимальной интенсивности первоначально полное (или почти полное) рекрутирование всех двигательных единиц сопровождается в дальнейшем постепенным снижением эффективности их участия в развитии мышцей напряжения, т. е. проявлением утомления.
Энергетика мышц. Мышцы используют универсальный источник энергии – аденозинтрифосфорную кислоту.
Под действием фермента АТФ-аза АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты (НЗР04), и превращается в АДФ, при этом высвобождается энергия.
АТФ + Н20 = АДФ + НЗР04 + энергия
Головка миозинового мостика при контакте с актином обладает АТФазной активностью и соответственно возможностью расщеплять АТФ и получать энергию, необходимую для движения.
Запас молекул АТФ в мышце ограничен, поэтому расход энергии при работе мышцы требует постоянного его восполнения.
Мышца имеет три источника воспроизводства энергии: расщепление креатинфосфата; гликолиз; окисление органических веществ в митохондриях.
Креатинфосфат обладает способностью отсоединять фосфатную группу и превращаться в креатин, присоединяя фосфатную группу к АДФ, которая превращается в АТФ.
АДФ + креатинфосфат = АТФ + креатин
Эта реакция получила название - реакции Ломана. Запасы креатинфосфата в волокне не велики, поэтому он используется в качестве источника энергии только на начальном этапе работы мышцы, до момента активизации других более мощных источников - гликолиза и кислородного окисления. По окончании работы мышцы, реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Анаэробный (лактатный) гликолиз) - процесс распада одной молекулы глюкозы (С6Н1206) на две молекулы молочной кислоты (СЗН603) с выделением энергии, достаточной для "зарядки" двух молекул АТФ, протекает в саркоплазме под воздействием 10 специальных ферментов.
С6Н1206 + 2НЗР04 + 2АДФ = 2СЗН603 + 2АТФ + 2Н20.
Гликолиз протекает без потребления кислорода (такие процессы называются анаэробными) и способен быстро восстанавливать запасы АТФ в мышце.
Аэробный (алактатный) гликолиз) протекает в митохондриях под воздействием специальных ферментов и требует затрат кислорода, а соответственно и времени на его доставку. Такие процессы называются аэробными. Окисление происходит в несколько этапов. Сначала идет гликолиз, но образовавшиеся в ходе промежуточного этапа этой реакции две молекулы пирувата не преобразуются в молекулы молочной кислоты, а проникают в митохондрии, где окисляются в цикле Кребса до углекислого газа С02 и воды Н20 и дают энергию для производства еще 36 молекул АТФ. Суммарное уравнение реакции окисления глюкозы выглядит так:
С6Н1206 + 602 + 38АДФ + 38НЗР04 = 6С02 + 44Н20 + 38АТФ
Итого распад глюкозы по аэробному пути дает энергию для восстановления 38 молекул АТФ. То есть окисление в 19 раз эффективнее гликолиза.
Далее следует окисление – распад свободных жирных кислот, кетоновых тел и т.п.
Типы мышечных волокон. Существование различных типов волокон в скелетных мышцах достаточно очевидно и их изучение продолжается на протяжении длительного периода. Однако детальные физиологические и биохимические основы этой дифференциации, а также их функциональное значение установлены лишь в последнее время. Во многом стимулами для этих исследований явилось понимание того, что успех в спортивных соревнованиях, требующих способности спортсмена к проявлению либо максимальной силы, либо выносливости, существенно зависит от пропорционального соотношения в мышце типов волокон. Мышечные волокна, однако, чрезвычайно пластичны, и хотя их соотношение генетически детерминировано, реализация определенной тренировочной программы способна во многом повлиять на метаболический потенциал мышцы независимо от соотношения в ней типов волокон.
Первоначальной основой для разделения мышечных волокон на красные, белые и промежуточные применительно к целостной мышце послужил простой визуальный осмотр. Однако основной функциональной характеристикой дифференциации типов мышечных волокон является скорость их укорочения и расслабления. Медленносокращающиеся волокна характеризуются относительно продолжительным временем, необходимым для достижения пика напряжения (около 80-100 мс для волокон мышц человека), а также длительным полупериодом расслабления. В противоположность им для достижения пика напряжения быстросокращающимися волокнами у человека требуется около 40 мс, следовательно, время их расслабления соответственно короче. Эти два типа волокон образуют отдельные группы с частичным совпадением их свойств.
Поскольку для изучения механических свойств волокон доступ к ткани затруднителен, а небольшие образцы мышцы относительно легко могут быть получены с помощью пункционной биопсии, классификация типов мышечных волокон обычно базируется на биохимическом окрашивании их поперечных срезов. На этой основе волокна мышц человека обычно подразделяют на три главные группы — типы I, IIа и IIб, хотя возможно также дальнейшее подразделение. Аналогичное деление свойственно и мышцам животных, волокна которых классифицируются на основе прямого определения их функциональных свойств: соответственно медленносокращающиеся, быстросокращающиеся медленноутомляемые и быстросокращающиеся быстроутомляемые.
Миозин различных типов мышечных волокон существует в различных молекулярных формах (изоформах), а АТФ-азная активность миофибрилл проявляет неодинаковую чувствительность к рН. Миозиновая АТФ-аза волокон типа II инактивируется при низких значениях рН (менее 4,5), тогда как АТФ-азная активность миозина в волокнах типа I остается неизменной. При значениях, превышающих рН 9, ситуация изменяется: АТФ-азная активность миозина в волокнах типа II остается стабильной, в то время как аналогичная активность в волокнах типа I снижается. Эти два отдельных подтипа типа II можно распознать путем преинкубации волокон при значениях рН, которые находятся в конце указанного диапазона: АТФ-азная активность миозина в волокнах типа IIа инактивируется при рН 4,6-4,8, в то время как в волокнах типа IIб она не изменяется. Иногда путем преинкубации можно выявить и волокна типа IIв, но этот тип вряд ли стоит принимать во внимание, поскольку в мышцах человека он составляет менее 1 % всех волокон.
Волокна типа IIа являются красными клетками, в которых метаболические и физиологические характеристики находятся между крайними значениями других двух типов волокон. Они содержат высокоактивную миозиновую АТФ-азу, аналогичную волокнам типа IIб, однако характеризуются окислительной способностью, более близкой к волокнам типа I.
Результаты последних исследований показывают, что для большинства мышечных волокон их фактическая идентификация определяется одной и более изоформой миозина. Такая совместная экспрессия различных изоформ миозина создает предпосылки для расширения диапазона сократительных характеристик внутри определенного типа волокон (классифицируемых в соответствии с окраской гистологических образцов) и степени их частичного совпадения между типами волокон при определении их сократительных характеристик. В действительности мы продолжаем оперировать сократительными и биохимическими характеристиками, соответствующими двум крайним типам классификации, определяемым как волокна типов I и IIб.
С различиями в скорости сокращения и метаболического профиля основных типов мышечных волокон отмечается дифференциация и двигательных нейронов, иннервирующих эти волокна. Так, волокна типа I иннервируются нейронами, имеющими малый диаметр и характеризующимися низкой скоростью проведения потенциала действия, а также низким порогом активации. Волокна типа II иннервируются нейронами большего диаметра, которые отличаются относительно более высокой прочностью и более высоким порогом возбудимости. Различие в порогах возбудимости двигательных нейронов связано с типом мышечных волокон, которые они иннервируют, с их рекрутированием при мышечных нагрузках, что и определяет метаболический ответ на последние.
Для упрощения понимания можно провести аналогию:
1 тип сравнивается с паровозом.
Используется любое топливо – мазут, уголь, дрова. Энергия используется опосредованно – вначале нагревается вода, затем сжатый пар давит на поршни и приводит механизм в движение.
В нашем случае это свободные жирные кислоты, кетоны, молочная кислота. Они тоже преобразуются в митохондриях в АТФ, который уже используется как источник энергии. Двигаться можно долго, так как топлива много, но большую мощность и скорость не разовьёшь.
2б тип – это ракета.
Используется топливо определённого типа. Вся энергия используется напрямую. При этом развивается огромная мощность, но ресурсы ограничены и быстро исчерпываются.
В мышцах это АТФ, креатинфосфат, для быстрого ресинтеза АТФ.
Скорость протекания данных процессов высокая, что и позволяет развивать большую мощность, но количество АТФ и креатинфосфата невелико, а это, в свою очередь, ограничивает время работы.
2а тип аналогичен автомобилю.
Топливо тоже определённого типа. Используется достаточно эффективно. Промежуточный вариант между паровозом и ракетой. При относительно высокой мощности можно достаточно долго двигаться.
В мышцах это гликоген, который может использоваться быстро, без участия кислорода (лактатный гликолиз), но малоэффективно, с выделением молочной кислоты. По аналогии с автомобилем – турбонаддув, топливо не успевает сгорать полностью. При расщеплении глюкозы под воздействием кислорода (аэробный гликолиз) скорость реакции ниже, но продолжительность работы дольше, сохраняя достаточную мощность.
При различных видах двигательной активности проявляется определенная иерархия включения в нее двигательных единиц, в зависимости от их размера, которая, в общих чертах, соответствует волокнам типов I и II. Это свидетельствует о том, что, при легких физических нагрузках, в наибольшей степени задействованы волокна типа I, при нагрузках средней мощности — волокна типов I и II, а при более напряженной мышечной деятельности — все типы волокон включаются в работу.
В се
мышцы организма представляют собой
смесь указанных выше трех
типов волокон, хотя пропорциональное
соотношение в каждой из них
и в каждом из индивидуумов обнаруживает
существенное различие.
се
мышцы организма представляют собой
смесь указанных выше трех
типов волокон, хотя пропорциональное
соотношение в каждой из них
и в каждом из индивидуумов обнаруживает
существенное различие.
100
80
60
40
20


 IIб
IIб


 IIa
IIa


 I
I




время
Схема рекрутирования мышечных волокон, в зависимости от интенсивности нагрузки.
Нагрузка в % от максимума |
Тип волокон |
Продолжительность |
Источник энергии |
Примерное время восстановления |
85 -100 |
|
|
|
|
75 - 80 |
|
|
|
|
65 - 70 |
|
|
|
|
55 - 60 |
|
|
|
|
50 - 55 |
|
|
|
|
40 - 45 |
|
|
|
|
Так, мышцы, отвечающие за поддержание позы тела, содержат высокий процент (обычно более 70 %) волокон типа I, функция которых состоит в поддержании пролонгированного, но относительно невысокого напряжения. Однако быстросокращающиеся волокна типа II преобладают в мышцах, выполняющих быстрые движения (например, в мышцах руки и глаза). Другие же мышцы, как, например, четырехглавая мышца бедра, имеют разное соотношение типов волокон. Их композиционный состав генетически детерминирован и не поддается значительным изменениям под влиянием тренировки. Следовательно, способность к достижению высоких спортивных результатов является в значительной степени врожденной (с учетом того, что генетический потенциал индивидуума реализуется при соответствующей методике тренировки и питания).
Исследование m. vastus lateralis у марафонцев высокой квалификации показало, что эта мышца. содержит больший процент (около 80 %) волокон типа I, тогда как у элитных спринтеров обнаружился более высокий процент (около 60 %) быстросокращающихся волокон типа II
Безусловно, такие исследования требуют применения методики пункционной биопсии, которая позволяет изъять около 1000 волокон в одной пробе. Относительно небольшое количество исследований по изучению распределения волокон в мышцах человека было проведено при вскрытии трупов. Региональная вариативность композиционного состава мышц проявляется незначительно, хотя и отмечается тенденция к тому, что в глубоколежащих слоях преобладают волокна типа I. Это подтверждается и данными, полученными с помощью достаточно надежного метода — пункционной биопсии. Исследования, проведенные на m. vastus lateralis у человека, свидетельствуют о том, что разница в показателях между пробами составляет не более 6 % (коэффициент вариации) по отношению к доминирующему типу волокон.




Пластичность скелетных мышц. Скелетная мышца является чрезвычайно пластичной тканью: она обладает значительной способностью адаптироваться к различным видам активности либо к бездействию. Адаптация может проявляться в изменении размеров мышцы, композиционного состава волокон, метаболической способности, плотности капилляризации.
Размер мышц и их функции с возрастом изменяются. Максимальной силы мышцы у мужчин и женщин достигают в возрасте между 20 и 30 годами. К 70 годам сила мышц снижается в среднем на 30 %. Уменьшение мышечной массы является ведущим фактором в этом процессе. Возможно также, что происходящее с возрастом снижение мышечной силы связано с уменьшением размеров волокон, в частности волокон типа II. Не исключено уменьшение при этом общего количества мышечных волокон, обусловленное утратой к старости двигательных нейронов. Иннервация мышечных волокон необходима для поддержания их существования (возможно, это связано с выработкой в нерве факторов роста), а денервация приводит к атрофии мышечных волокон и замещению их соединительной тканью.
Связанное со старением снижение мышечной массы может происходить как за счет самих возрастных изменений, происходящих в организме, так и в результате снижения двигательной активности либо одновременно за счет обоих факторов. Однако ясно, что и в старом организме мышцы еще сохраняют способность адаптироваться в ответ на силовую тренировку и что значительного улучшения физиологических, структурных и физических характеристик можно достичь без реализации напряженных тренировочных программ. В молодом возрасте частота, интенсивность и продолжительность упражнений являются решающими факторами, определяющими диапазон происходящих под влиянием тренировки адаптационных изменений.
Возрастные этапы становления энергетики мышечной деятельности. Первый год жизни ребенка представляет собой период бурного становления мышечной функции и, разумеется, ее энергетического и вегетативного обеспечения. Этот этап продолжается до возраста 3 лет, после чего преобразования в мышцах тормозятся, и следующий этап начинается вместе с полуростовым скачком примерно в 5 лет. Важнейшим событием здесь является появление уже близких к взрослому варианту типов мышечных волокон, хотя их соотношение еще является «детским», да и функциональные возможности вегетативных систем еще недостаточно велики. В школьном возрасте ребенок проходит еще целый ряд этапов, только на последнем из них достигая «взрослого» уровня регуляции, функциональных возможностей и энергетики скелетных мышц:
1-й этап — возраст от 7 до 9 лет ______________________________________________
____________________________________________________________________________________________________________________________________________________________________________________________________________________
2-й этап — возраст 9—10 лет ________________________________________________
____________________________________________________________________________________________________________________________________________________________________________________________________________________
3-й этап — период от 10 до 12—13 лет _________________________________________
____________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________
4-й этап — возраст от 13 до 14 лет _____________________________________________
____________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________
5-й этап — возраст 14—15 лет _______________________________________________
________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________
6-й этап — период от 15 до 17 лет _____________________________________________
________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________
На процессы созревания энергетических и вегетативных систем огромное влияние оказывает половое созревание, так как половые гормоны непосредственно влияют на метаболические возможности скелетных мышц. Аэробное энергообеспечение, достигающее расцвета еще до начала пубертата, на первых его стадиях даже несколько ухудшается, однако к возрасту 14 лет отмечается новый рост возможностей аэробных систем энергообеспечения. Это связано, в частности, с внутренними потребностями мышц, которым для последнего этапа дифференцировок требуются мощные окислительные системы. Анаэробное энергообеспечение резко активируется уже на начальных стадиях полового созревания, затем (III стадия) темп его совершенствования замедляется, а после достижения IV стадии полового созревания (15—16 лет у мальчиков, 13—14 лет у девочек) наблюдается бурный рост анаэробных возможностей, особенно у юношей. Девушки в этот период уже сильно отличаются от юношей по характеру и уровню развития мышечной энергетики.