

Вопросы к экзамену.
-
Строение гена и биосинтез белка в прокариотической клетке.
Прокариоты – одноклеточные организмы, лишенные ядра, с хромосомами, находящимися в цитоплазме.
Ген – участок ДНК, кодирующий одну полипептидную цепь или одну молекулу tРНК, rРНК или sРНК
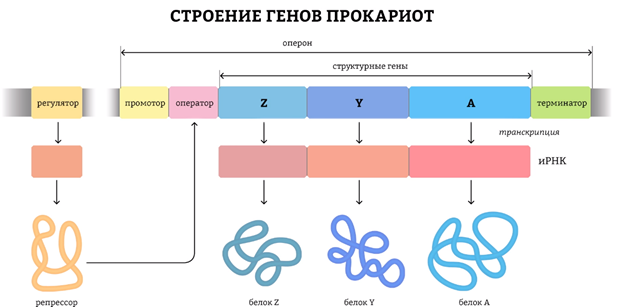
Гены прокариот состоят из двух основных элементов: регуляторной части и собственно кодирующей части. Регуляторная часть обеспечивает первые этапы реализации генетической информации, а кодирующая часть содержит информацию о структуре полипептида, тРНК, рРНК. У прокариот структурные гены, кодирующие белки одного метаболического пути, часто бывают объединены и называются опероном. Оперон – это участок ДНК, ограниченный с одной стороны промотором (местом присоединения РНК-полимеразы), с другой – терминатором (местом ее отсоединения). Оперон кодирует одну молекулу мРНК, на основе которой позже могут синтезироваться один или несколько белков. В опероне имеется оператор, “разрешающий” или “не разрешающий” работу РНК-полимеразы
У прокариот большинство генов “включено” (с них идет синтез белков), поэтому задача регуляции у них чаще всего сводится к их “выключению” с помощью веществ-репрессоров, причем один определенный репрессор влияет только на свой ген (или на небольшую группу генов) и не действует на другие. Упрощает регуляцию синтеза белка прокариотами наличие у них координированной регуляции: если несколько разных цистронов (то есть генов, кодирующих белки) регулируются одной регуляторной зоной (промотор + оператор), то воздействием на эту зону можно регулировать синтез сразу всех белков, закодированных этими цистронами.
Начало синтеза у прокариот и эукариот существенно различается. Это связано с тем, что у прокариот гены собраны в «пачки» для удобства регуляции. У них полицистронный принцип организации генов, при котором одна длинная информационная РНК считывается сразу с нескольких генов, участвующих в одном биохимическом процессе. Это удобно для регулирования процессов внутри клетки.
Прокариоты в большинстве случаев используют негативную регуляцию, через ген-специфичные репрессоры, действующие на промоторы структурных генов.
У прокариот, в отличие от эукариот, транскрипция и трансляция происходят в одном месте и в одно время. У них нет ядра, поэтому синтез белка может начинаться еще до окончания синтеза РНК. РНК-полимераза может считывать полицистронную мРНК, по матрице которой одновременно может начаться синтез белка.
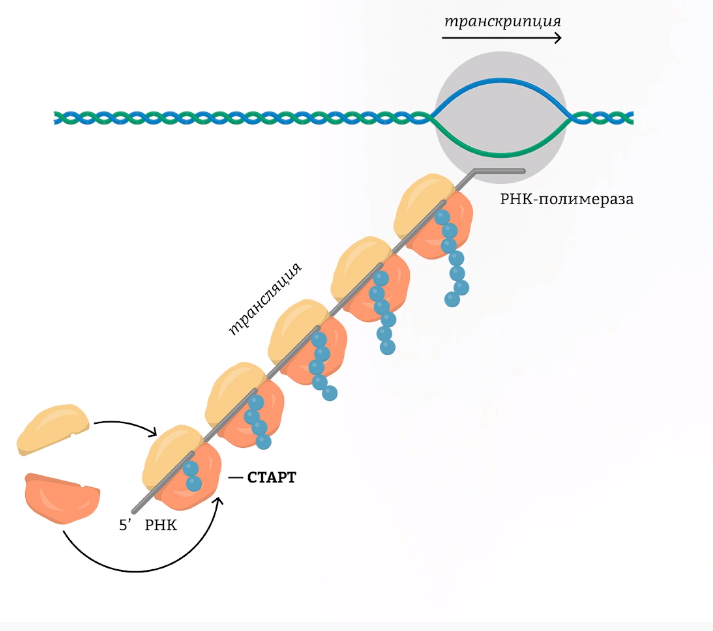
В связи с тем, что гены прокариот не имеют экзонов и интронов (исключение — гены архебактерий), располагаются группами, и на эту группу генов приходится один промотор, появляются следующие особенности транскрипции и трансляции у прокариот.
1. В результате транскрипции образуется полицистронная иРНК, кодирующая несколько белков, совместно обеспечивающих определенную группу реакций.
2. иРНК имеет несколько центров инициации трансляции, терминации трансляции и НТО.
3. Не происходят КЭПирование, полиаденилирование и сплайсинг иРНК.
4. Трансляция начинается еще до завершения транскрипции; эти процессы не разделены во времени и пространстве, как это имеет место у эукариот.

1 — ДНК; 2 — РНК-полимераза; 3 — Нуклеозидтрифосфаты ГТФ, ЦТФ, АТФ, УТФ.
Можно добавить, что срок «жизни» прокариотических иРНК — несколько минут (у эукариот — часы и даже сутки).
-
Регуляторные элементы в ДНК: промоторы, терминаторы, участок Шайн-Далгарно, инициирующие и терминирующие кодоны, ori.
Промотор – последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как стартовая площадка для начала транскрипции. Промотор играет одну из ключевых ролей в процессе инициации транскрипции.
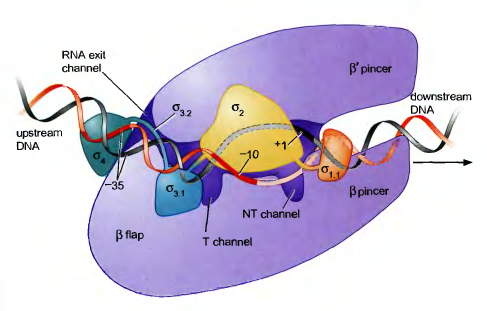
Коровая РНК-полимераза бактерий (состоящая из субъединиц α2ββ'ω) может инициировать транскрипцию в любом месте генома. Однако, в клетке инициация происходит только в промоторных участках. Такая специфичность обеспечивается σ-субъединицей (σ-фактор), которая в комплексе с коровым ферментом образует холофермент. Основным σ-фактором клеток Escherichia coli является σ70-субъединица.

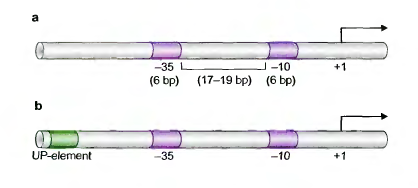
Классический (σ70) промотор представляет собой две консервативные последовательности длиной по 6 нуклеотидов, расположенные выше сайта начала транскрипции на 10 и 35 п.о., разделенные 17 нуклеотидами. Эти последовательности называются соответственно -10 и -35 элементами. Элементы не идентичны во всех промоторах, но для них можно получить консенсусные последовательности.
Некоторые сильные промоторы также имеют UP-элемент, расположенный выше -35-элемента, который повышает уровень связывания РНК-полимеразы. Некоторые σ70 промоторы не имеют -35-элемента, зато имеют -10-элемент, расширенный вверх на несколько нуклеотидов (extended -10). Таков промотор галактозного оперона E.coli. Иногда ниже -10-элемента располагается ещё один связывающий элемент – дискриминатор.

Терминатор (англ. Terminator) — нуклеотидная последовательность ДНК, на которой завершается транскрипция гена или оперона. Как правило, последовательность терминатора такова, что комплементарная ей последовательность в мРНК вызывает выход новосинтезированного транскрипта из транскрипционного комплекса. Эта последовательность в мРНК может сама по себе вызывать терминацию за счёт собственной вторичной структуры, а может привлекать особые белки — факторы терминации. После высвобождения РНК-полимераза и транскрипционные факторы приступают к транскрипции другого гена.
У прокариот существует два механизма терминации: ро-зависимый и ро-независимый.
Ро-зависимые терминаторы функционируют посредством особого белка — ро-фактора, который обладает РНК-хеликазной активностью и разрушает комплекс ДНК, мРНК и РНК-полимеразы. Ро-зависимые терминаторы встречаются у бактерий и фагов. Ро-зависимые терминаторы находятся ниже стоп-кодона, на котором завершается трансляция, и представляют собой неструктурированные, обогащённые цитозином последовательности в мРНК, известные как rut-сайты (от англ. Rho utilization site), после которых находятся точки остановки транскрипции (англ. tsp от transcription stop point). Консенсусная последовательность для rut-сайтов не установлена. Rut-сайт выступает в роли места посадки ро-фактора на мРНК и его активатора. Активированный ро-фактор начинает гидролизовать АТФ и за счёт энергии гидролиза движется вдоль мРНК, пока не столкнётся с РНК-полимеразой. Контакт между ро-фактором и РНК-полимеразой стимулирует распад транскрипционного комплекса за счёт аллостерических эффектов ро-фактора на РНК-полимеразу.
Ро-независимые терминаторы формируют шпильки в структуре синтезирующегося транскрипта, которые при столкновении с РНК-полимеразой вызывают диссоциацию комплекса ДНК, мРНК и РНК-полимеразы. Типичный ро-независимый терминатор состоит из 20 нуклеотидов, обогащён парами GC и обладает диадной симметрией, а вслед за ним следует обогащённый тимином участок (поли(Т)-тракт), которому в мРНК соответствует участок, обогащённый урацилом. Предположительный механизм работы ро-независимых терминаторов заключается в том, что шпилька вызывает остановку РНК-полимеразы, из-за которой повышается вероятность диссоциации фермента от матрицы.


Старт-кодон или инициаторный кодон — первый кодон матричной РНК, c которого начинается трансляция белка в рибосоме. У эукариот и архей старт-кодон всегда кодирует метионин, а у бактерий — модифицированный метионин (N-формилметионин). В большинстве случаев роль инициаторного кодона играет триплет AUG. Старт-кодону предшествует 5′-нетранслируемая область (5'-UTR). В 5'-UTR бактерий локализована последовательность Шайна — Дальгарно (AGGAGG), которая служит для связывания рибосомы и отделёна спейсером от старт-кодона.

Альтернативные старт-кодоны отличаются от стандартного кодона AUG. Такие кодоны встречаются как у прокариот, так и у эукариот. Альтернативные старт-кодоны обычно кодируют метионин, когда они находятся в начале белка (даже если кодон кодирует другую аминокислоту). Так например, кодон GUG кодирует валин в случае, если он находится внутри кодирующей последовательности, и стартовый метионин, если расположен в начале последовательности. Это происходит потому, что для инициации трансляции используется специальная транспортная РНК. Антикодоном инициирующей аминоацил-тРНК всегда является CAU; он полностью комплементарен основному старт-кодону AUG и частично комплементарен более редким кодонам. Кроме частично комплементарных GUG и UUG в исключительных случаях, особенно в клетках бактерий, инициация может начинаться с триплетов AUU, AUA, ACG и CUG. Эти так называемые «слабые» кодоны могут выполнять свою функцию в комбинации с сильными последовательностями Шайна-Дальгарно или другими структурными элементами, способствующими инициации.
Последовательность Шайна — Дальгарно (англ. Shine-Dalgarno sequence, Shine-Dalgarno box) — сайт связывания рибосом на молекуле мРНК прокариот, обычно на расстоянии около 10 нуклеотидов до стартового кодона AUG. Описана австралийскими учёными Джоном Шайном и Линн Дальгарно.
Консенсусом является последовательность из шести нуклеотидов AGGAGG; в случае E. coli последовательность Шайна — Дальгарно — AGGAGGU. Комплементарная последовательность CCUCCU, называемая последовательностью анти-Шайна — Дальгарно, располагается на 3'-конце молекулы 16S рибосомной РНК. Комплементарное взаимодействие между последовательностями Шайна — Дальгарно и анти-Шайна — Дальгарно служит для помещения старт-кодона мРНК в P-сайт рибосомы для начала биосинтеза белка.

Мутации в последовательности Шайна — Дальгарно снижают эффективность трансляции. Это обусловлено снижением эффективности образования комплекса мРНК-30S рибосомная субъединица. Показано, что комплементарные мутации в последовательности анти-Шайна — Дальгарно обеспечивают восстановление эффективности трансляции.
В момент образования комплекса последовательности Шайна — Дальгарно и анти-Шайна — Дальгарно, с 30S-рибосомной субъединицей связываются и факторы инициации трансляции IF2-GTP, IF1, IF3, а также инициаторная формилметионил-тРНК (fMet-tRNA). К образовавшемуся преинициаторному комплексу затем присоединяется 50S-рибосомная субъединица.
Стоп-кодон или кодон терминации — единица генетического кода, тройка нуклеотидных остатков (триплет) в ДНК — кодирующая прекращение (терминацию) синтеза полипептидной цепи (трансляцию).
Стоп-кодоны выполняют важную функцию завершения (терминацию) сборки полипептидной цепи и также называются терминаторными кодонами. Некоторые из них вызывают обязательное прекращение синтеза, другие являются условными.
Кроме того, стоп-кодон, как кодон, при котором не происходит включения аминокислоты в белок, ещё называют бессмысленным кодоном или нонсенс-кодоном.
Так, кодон UAG (Янтарь) — условный терминаторный кодон и супрессируемые Amber-мутации вызывают преждевременную терминацию трансляции (условно летальные мутации).
Сквозная трансляция может проходить через кодоны UAG (Янтарь) и UGA (Опал), но не через кодон UAA (Охра).
Кодоны UAA и UAG в митохондриальной ДНК вызывают безусловное прекращение трансляции.
Некоторые мРНК в действительности содержат два тандемных терминаторных кодона — часто это кодоны различного типа на конце кодирующей последовательности.
Точка начала репликации (англ. origin of replication = ori) — это фрагмент молекулы нуклеиновой кислоты, с которого начинается её репликация. Структура точки начала репликации (нуклеотидная последовательность) отличается у разных видов, но у всех организмов это АТ-богатая и потому легкоплавкая последовательность. Точка начала репликации и прилегающие к ней фрагменты нуклеиновой кислоты, не отделённые сайтами терминации, составляют единицу репликации — репликон. Репликация ДНК может начинаться от точки начала репликации в одном или двух направлениях.
Хромосомы и плазмиды прокариот содержат одну, реже несколько точек начала репликации ДНК; хромосомы эукариот имеют множество таких точек. Также точки начала репликации РНК обнаружены у РНК-содержащих вирусов, например, у вирусов, содержащих двуцепочечные РНК.


-
Передача и изменение генетической информации: трансформация, конъюгация, трансдукция, рекомбинация, мутации, репарация.
В основе генотипической изменчивости лежат мутации и рекомбинации.
1. Мутации - внезапные, скачкообразные изменения наследственных свойств. Основу этого явления составляют качественные или количественные изменения последовательности нуклеотидов в ДНК, которые могут возникать при жизнедеятельности организмов под влиянием внешних и внутренних факторов. Организмы с измененными признаками называются мутантами.
Различают спонтанные и индуцированные мутации.
• Спонтанные (самопроизвольные) мутации возникают под влиянием неизвестных причин. Частота спонтанных мутаций мала. Спонтанные мутации служат основным источником естественной изменчивости микроорганизмов и лежат в основе эволюции.
• Индуцированные (направленные) мутации проявляются в результате обработки микроорганизмов специальными мутагенами (физическими факторами и химическими веществами). В основе механизма лежит прямое или косвенной влияние на ДНК.
2. Рекомбинация
Перестройка заключенной в молекулах ДНК генетической информации осуществляется благодаря протеканию многих разнообразных процессов, обобщенно называемых генетической рекомбинацией.
Выделяют три основных типа генетической рекомбинации.
Гомологичная генетическая рекомбинация (общая рекомбинация) представляет собой обмен генами между двумя любыми молекулами ДНК (или участками одной и той же молекулы), которые содержат протяженные отрезки почти идентичной последовательности. Последовательности могут быть любыми, главное, чтобы они были похожими. При сайт-специфической рекомбинации происходят обмен только определенных последовательностей ДНК. Транспозиция ДНК отличается от двух других типов генетической рекомбинации тем, что в ней обычно участвует короткий участок ДНК, обладающий способностью перемещаться по хромосоме с одного сайта на другой.
Функции систем генетической рекомбинации так же разнообразны, как и их механизмы. К ним относится участие в специализированных системах репарации ДНК, определенная активность при репликации ДНК, регуляция экспрессии некоторых генов, облегчение расхождения хромосом при делении эукариотических клеток, поддержание генетического разнообразия и реализация запрограммированных генетических перестроек в ходе эмбрионального развития.
Трансформация - направленный перенос генетической информации от донорских клеток в реципиентные с помощью изолированной ДНК. При этом в хромосому реципиента включается только одна нить ДНК донора с образованием молекулярной гетерозиготы.
Конъюгация - форма полового процесса, при котором происходят соединение мужской и женской микробных клеток и обмен между ними ядерным веществом через цитоплазматический мостик, образующийся между клетками. При этом генетический материал клетки-донора переходит в клетку-реципиент. После рекомбинации и деления клетки образуются формы с признаками конъюгирующих клеток.
Трансдукция – перенос генетической информации из одной бактериальной клетки в другую с помощью бактериофагов.
Таким образом, все три формы комбинативной изменчивости одинаковы по существу. При трансформации участок ДНК клетки-донора входит в клетку-реципиент; при трансдукции эту роль выполняет фаг, а при конъюгации перенос генетической информации осуществляется через цитоплазмитический мостик (пили).
Вследствие генетических рекомбинаций образуются новые бактериальные клетки – рекомбинанты, у которых имеются наследственные признаки обоих «родителей».
3. Репарация - особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённых при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических реагентов. Осуществляется специальными ферментными системами клетки.
Каждая из систем репарации включает следующие компоненты:
 • ДНК-хеликаза
— фермент, «узнающий» химически
изменённые участки в цепи и осуществляющий
разрыв цепи вблизи от повреждения;
• ДНК-хеликаза
— фермент, «узнающий» химически
изменённые участки в цепи и осуществляющий
разрыв цепи вблизи от повреждения;
• ДНКаза (дезоксирибонуклеаза) — фермент, "разрезающий" 1 цепочку ДНК (последовательность нуклеотидов) по фосфодиэфирной связи и удаляющий повреждённый участок: экзонуклеаза работает на концевые нуклеотиды 3` или 5`, эндонуклеаза - на нуклеотиды, отличные от концевых;
• ДНК-полимераза — фермент, синтезирующий соответствующий участок цепи ДНК взамен удалённого;
• ДНК-лигаза — фермент, замыкающий последнюю связь в полимерной цепи и тем самым восстанавливающий её непрерывность.
У бактерий имеются по крайней мере 3 ферментные системы, ведущие репарацию — прямая, эксцизионная и пострепликативная. У эукариот к ним добавляется ещё Mismatch и SOS-репарация.
• Прямая репарация
Прямая репарация — наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов.
• Эксцизионная репарация
Эксцизионная репарация (англ. excision — вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы по комплементарной цепи. Ферментативная система удаляет короткую однонитевую последовательность двунитевой ДНК, содержащей ошибочно спаренные или поврежденные основания, и замещает их путём синтеза последовательности, комплементарной оставшейся нити.
• Пострепликативная репарация
Tип репарации, имеющей место в тех случаях, когда процесс эксцизионной репарации недостаточен для полного исправления повреждения: после репликации с образованием ДНК, содержащей повреждённые участки, образуются одноцепочечные бреши, заполняемые в процессе гомологичной рекомбинации при помощи белка RecA.
-
Этапы получения сверхпродуцентов методами ги
Сверхпродуцент – мутантный штамм микроорганизма, продуцирующий большое количество какого-либо вещества (напр., белка). Сверхпродуценты обладают более высокими показателями скорости наработки целевого вещества, либо имеют и другие полезные свойства, которые, в основном, делают производство более прибыльным.
Для получения сверхпродуцентов используют селекцию, мутагенез, генетическую инженерию. При селекции генотип подвергается изменениям косвенно, в рез-те улучшаются их естественные способности синтезировать целевой продукт (аминокислоты, витамины, ферменты и др.). В случае использования методов генной инженерии происходит непосредственное вмешательство в генетический аппарат путем рекомбинации хромосом или отдельных генов in vitro.
Возможности генной инженерии:
1) можно скрещивать гены видов, стоящих на разных ступенях эволюции,
2) можно управлять процессом рекомбинации, т.к. он происходит в пробирке и не защищен запрещающими механизмами клетки,
3) можно предсказать результат, т.к. отбирается потомство одной молекулы ДНК (молекулярное клонирование).
Т.о. продуцент возможно перепрограммировать на выработку соединений, синтез которых в естественных природных условиях им никогда не был присущ (гормоны человека и животных).
Этапы ГИ:
-
Выделение гена, который планируем встроить в генетический аппарат клетки-реципиента. Для получения ядерной ДНК сначала бактериальные клетки обрабатывают ферментом, разрушающим клеточную стенку, затем клеточные белки экстрагируют смесью фенола и хлороформа,а ДНК из растворимой фракции осаждают охлажденным этанолом. Чтобы выделить клеточную ДНК из животных клеток, их лизируют с помощью додецилсульфата натрия (ДСН, или англ. SDS), а затем обрабатывают РНКазой и протеиназой К,которые разрушают РНК и белки. Белки и SDS удаляют высаливанием, а ДНК из раствора осаждают спиртом.

После выделения гена ее обрабатывают рестриктазами (вид гидролаз) дабы расщепить ДНК цепь на более короткие фрагменты. Возможно обрабатывание выделенной ДНК другими ферментами (лиазы, синтетазы, модифицирующие ферменты).
Затем необходимо провести ПЦР. В ходе ПЦР происходит многократное увеличение числа копий специфических фрагментов ДНК. Другими словами, ПЦР позволяет селективно амплифицировать выбранный фрагмент ДНК. Метод ПЦР позволяет применить амплификацию ДНК при решении многих молекулярно-генетических задач, например, введение в ДНК функциональных элементов; соединение двух фрагментов ДНК; встраивание новых участков в последовательность ДНК; удаление фрагментов ДНК из гена; осуществление сайт-направленного мутагенеза.
Иногда возможно не выделение необходимого гена, а его непосредственный синтез. Это возможно, только если нужный нам ген будет небольшого размера (не более 100 пар нуклеотидов).
-
Встраивание полученного гена в генетический элемент, способный к репликации (вектор). Конструирование рекомбинантных ДНК осуществляется in vitro. Кольцевая молекула вектора размыкается рестриктазой. Полученная линейная молекула ДНК должна содержать липкие концы, комплементарные концам вводимой ДНК. Комплементарные липкие концы вектора и вводимого гена сшивают ДНК-лигазой и полученную рекомбинантную ДНК с помощью той же ДНК-лигазы вновь замыкают с образованием единой кольцевой молекулы.
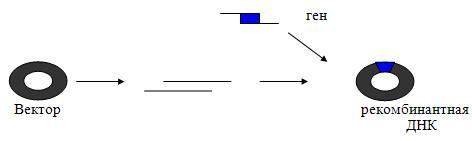
Различают два основных класса векторов: вирусы и плазмиды. Однако наибольшее применение в генетической инженерии нашли бактериальные плазмиды, особенно плазмиды E. Coli, т.к. вирусы хоть и эффективны, но из них сложно делать векторы.
-
Введение гена в составе вектора в организм-реципиент. Для бактерий достигается путем трансформации, однако трансформации могут подвергаться только компетентные клетки. Поэтому важным этапом является отделение компетентных клеток, подвергшихся трансформации, от некомпетентных клеток. Осуществить это можно следующим образом: в плазмиду дополнительно заранее встраиваем ген устойчивости к какому-нибудь антибиотику. Клетки, которые поглотили эту плазмиду, смогут жить в присутствии этого антибиотика, а другие клетки не смогут. В результате, чтобы выделить из всех бактерий, к которым мы добавили плазмиду, те, которые смогли эту плазмиду использовать по назначению, нам достаточно будет добавить к бактериальной культуре соответствующий антибиотик.
Затем выращиваем полученную культуру бактерий.
-
Экспрессия введенной последовательности ДНК. => получаем нужное нам вещество.
-
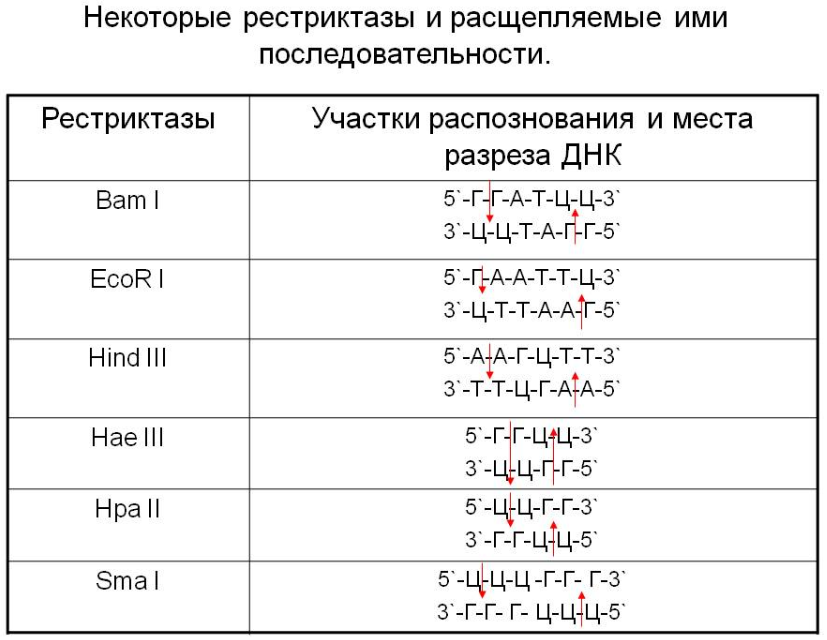
Система модификации-рестрикции. Рестриктазы, метилазы: номенклатура, классы рестриктаз.
Эндонуклеазы - группа ферментов, относящихся к классу гидролаз, катализирующих реакцию гидролиза нуклеиновых кислот.
В отличие от экзонуклеаз, рестриктазы расщепляют нуклеиновые кислоты не с конца молекулы, а в середине. При этом каждая рестриктаза узнаёт определённый участок ДНК длиной от четырёх пар нуклеотидов и расщепляет нуклеотидную цепь внутри участка узнавания или вне его.
Модификация заключается в метилировании определенных оснований в последовательности, узнаваемой сопряженной рестриктазой; тем самым обеспечивается защита данного участка ДНК от воздействия рестриктазы. Одновременное наличие в клетке этих двух ферментативных активностей (так называемая R-M система) препятствует гидролизу собственной нуклеиновой кислоты. Чужеродная же ДНК при проникновении в бактериальную клетку служит субстратом для обоих ферментов.
Первоначально многие считали, что единственной функцией R-M систем является защита клеток от инфицирования фагами. Однако дальнейшие исследования позволили сделать предположение о том, что R-M системы осуществляют функцию ограничения скрещивания между различными бактериальными видами и штаммами, которая, однако, не абсолютна и по-зволяет части чужеродной ДНК проникать в клетку, рекомбинационно встраиваться и поддерживаться в качестве генетического фонда для получения эволюционного преимущества. Уместно заметить, что у бактерий весьма проблематично определение вида. Существуют даже предположения об общем генофонде всех микроорганизмов, что должно было бы привести к бесконечному появлению новых видов бактерий во времени. Реально же мы видим, что бактерии проявляют определенное постоянство морфологических, генетических и биохимических характеристик: Достойными кандидатами для обеспечения в эволюции относительной стабильности генетического материала, т. е. для осуществления генетической изоляции, не отрицающей обмена определенными блоками, являются системы рестрикции-модификации.
Системы рестрикции и модификации найдены практически у всех исследованных бактерий. Недавно рестриктазы обнаружены и у некоторых видов дрожжей.
В 1968 г. М. Мезельсон и Р. Юань сообщили о выделении первой рестриктазы из штамма Е. coli К12. Подобный фермент был получен и из штамма Е. coli В. Данные эндонуклеазы ЕсоК и ЕсоВ отличались высокой специфичностью по отношению к узнаваемой последовательности нуклеотидов, yо расщепляли молекулы ДНК неспецифическив другом месте, отстоящем от участка (сайта) узнавания. В 1970 г. X. Смит иК. Вилькокс выделили из Haemophius influenzae рестриктазу HindII, не только специ-фически узнающую, но испецифически расщепляющую молекулы ДНК. При гидролизе вирусной или плазмидной ДНК рестриктазами такого типа образуется строго определенный набор фрагментов. Это наглядно выявляется при электрофоретическом разделении смеси получающихся фрагментов.
Принципиальное значение для разработки методологии генетической инженерии имело открытие в 1971 г. Р. Ёшимори рестриктаз EcoR I и EcoR II. Спомощью первой из них удалось выполнить пионерскую работу по направленной реконструкции генетического материала in vitro. В настоящее время рестриктазы используют практическиво всех генно-инженерных экспериментах. Такое широкое применение ферментов данного типа обусловлено их высокой специфичностью, а также особенностями структуры концов фрагментов ДНК, образуемых рестриктазами. Общепринято термины рестриктаза, эндонуклеаза рестрикции, сайтспецифическая эндодезоксирибонуклеаза считать синонимами.
X. Смит и Д. Натане в 1973 г. предложили номенклатуру рестриктаз, которая включает следующие пункты:
1.Название каждого фермента является производным от бинарного родо-видового обозначения микроорганизма-хозяина, содержащего данную R-M систему, и составляется по следующему правилу: к первой прописной букве названия рода добавляют две первые строчные буквы вида. Например: Streploinyces albiis - Sal, Escherichia coli - Eco.
2. За родо-видовым названием следует, в случае необходимости, обозначение серотипа или штамма: Haemophihis influenzae d - Hind. Escherichia coli В - ЕсоВ.
3.Различные системы рестрикции-модификации, кодируемые одной и той же бактериальной клеткой, обозначаются римскими цифрами, например: HindI, HindII, HindIII.
4. Ферменты рестрикции-модификации в общем виде обозначаются как эндонуклеаза R или метилаза М с последующим определением названия системы, например: эндонуклеаза R * HindII или метилаза М * HindII.
5. Если система генетически локализована в геноме фага или на плазмиде, то после родо-видового названия указывается символ внехромосомного элемента: EcoРI, EcoKII. Штаммовая принадлежность в этих случаях указывается в скобках: Есо(К)РI.
Рестриктазы I класса, к которым относятся, к примеру, ферменты ЕсоК, EcoB, EcoPl и другие, атакуют ДНК в произвольных местах и образуют сплошной спектр рестриктов. Эти рестриктазы в генетической инженерии не исполь-зуются, поскольку с их помощью нельзя получить фрагменты ДНК строго детерминированных размеров.
Рестриктазы класса II. Системы рестрикции-модификации класса II состоят из отдельных белков рестрикционной эндонуклеазы и модификационной метилазы. Поэтому рестриктазы данного класса можно выделить в индиви-дуальном состоянии, свободном от метилазной активности, что в значительной мере упрощает их изучение и последующее использование для расщепления молекул ДНК.
Рестриктазы класса IIIимеют некоторое сходство с рестриктазами класса I. Нативный фермент состоит из двух различных субъединиц и бифункционален, т. е. обладает как рестриктазной, так и метилазной активностью. Рестриктазы класса III узнают несимметричные последовательности длиной 5-6 пн и расщепляют ДНК в стороне от участков узнавания на расстоянии 24-27 пн, образуя одноцепочечные 5'-концы длиной 2-3 нуклеотида. Для проявления эндонуклеазной активности требуются только АТР и ионы Mg2+, a SAM лишь стимулирует реакцию,причем расщепление ДНК не сопровождается гидролизом АТР. При действии ферментов данного класса in vitro не удается исчерпывающе гидролизовать ДНК. Причины этого пока не ясны.
-
Ферменты, используемые в ГИ: рестриктазы, ДНК-лигаза, нуклеаза Bal31.
Рестриктазы (рестрицирующие эндонуклеазы, эндонуклеазы рестрикции) — это ферменты, узнающие и атакующие определенные последовательности нуклеотидов в молекуле ДНК (сайты рестрикции). (Щелкунов Генетическая инженерия)
Рестриктазы узнают строго определенные последовательности нуклеотид:


Лигаз — фермент, катализирующий соединение двух молекул с образованием новой химической связи –лигирование.


Bal31

