

История анатомии и физиологии человека
ПЕРИОДЫ И ЭТАПЫ РАЗВИТИЯ АНАТОМИИ
История анатомии является частью медицины и ее можно разделить на два периода:
Древний период (XX в. до н. э. — XV в. н. э.):
этап древней анатомии (анатомия в Древнем Китае, Индии, Египте, XX в. до н. э. — V в. н. э.);
этап примитивной описательной анатомии (Древняя Греция и Древний Рим, V–III в. до н. э.);
этап схоластической анатомии (Италия, Франция, Восток, II–XV вв. н. э.).
Период научной анатомии (начинается со времен Андреаса Везалия — XVI в. н. э. до н. в.):
этап макроскопической (описательной) анатомии (эпоха Возрождения, XVI–XVII вв.);
этап микроскопической (эволюционно-функциональной) анатомии (XVII–XX вв.);
этап ультрамикроскопической (молекулярной) анатомии (60-е гг. ХХ в. до н. в.).
ДРЕВНий период изучения АНАТОМИи
Первая известная школа древнегреческой медицины была открыта в Книде в 700 г. до н. э. В ней работал Алкмеон, автор первого труда по анатомии, и именно в этой школе была впервые применена практика наблюдения за пациентами. Позднее свою школу на острове Кос основал Гиппократ. Несмотря на общеизвестное уважение эллинов к египетской медицине, в те времена не было успешных попыток перенести опыт египтян на греческую землю из-за недостаточного количества письменных источников и трудностей в понимании древней медицинской терминологии. Известно, впрочем, что для своих медицинских справочников греки позаимствовали множество рецептов египетских лекарств. Влияние Египта стало более очевидным после создания греческой медицинской школы в Александрии.
Алкмеон Кротонский (V в. до н. э.) — древнегреческий философ, врач и ученый. Возможно, был учеником Пифагора. Автор первого древнегреческого медицинского трактата. Считал, что здоровье сохраняется при равновесии сил, то есть равновесии пар противоположностей: влажного и сухого, горячего и холодного, горького и сладкого (число пар противоположностей философ считал неопределенным). Болезнь наступает тогда, когда одна из сил берет господство. Основой терапии ученый считал лечение противоположного противоположным. Учение Алкмеона о здоровье оказало значительное влияние на античную медицинскую мысль.
Гиппократ (460–377 гг. до н. э.) — отец медицины. Сформулировал учение о четырех основных типах телосложения и темперамента, описал некоторые кости черепа и внутренние органы, суставы, мышцы и крупные сосуды.
Аристотель (384–322 гг. до н. э.) — ученик Платона. Описал сухожилия, нервы, кости и хрящи. Ввел термины «аорта» и «антропология». Отметил черты сходства человека с животными.
Герофил (ок. 335 до н. э. — ок. 280 до н. э.) — учитель медицины в Александрии. Внес исправления в труды Аристотеля, поместив разум в мозг и соединив нервную систему с движением и осязанием. Четко различил вены и артерии. Изучая пульсацию артерий, разработал диагностический метод, основанный на различении типов пульса. Он и его современник Эразистрат изучали роль и расположение вен и нервов на теле человека.
Эразистрат (ок. 335 до н. э. — ок. 280 до н. э.) — древнегреческий ученый, внук Аристотеля. Изучал строение человеческого мозга. Утверждал, что человеческая система кровеносных сосудов контролируется вакуумами, переносящими кровь по всему телу. Согласно физиологической теории Эразистрата, воздух поступает в тело, затем легкие гонят его в сердце, где он превращается в жизненную силу, которую артерии разносят по всему телу. Часть этой жизненной силы достигает мозга, где трансформируется в животный дух, который затем распространяется по нервам. Он предполагал в теле два противоположных элемента: жизненный дух и кровь.
Эразистрат изучал функции органов пищеварения на живых животных и наблюдал перистальтику желудка. Утверждал, что пищеварение происходит путем механического перетирания пищи желудком. Вскрывал человеческие трупы, описал печень и желчные протоки. Главной причиной болезней Эрасистрат считал излишества в пище, которая засоряет сосуды, вследствие этого возникают воспаления, язвы и другие заболевания.
Герофил и Эразистрат проводили свои эксперименты на заключенных, которых им выдавали птолемейские цари. Они проводили вскрытие на живых людях, и, «пока те еще дышали, изучали части тела, сокрытые природой от людского взора…».
Клавдий Гален (II в.) — римский медик, хирург и философ. Его теории доминировали в европейской медицине в течение 1300 лет. Изучал анатомию путем вскрытия свиней, собак, овец и других животных и был уверен в тождественности строения тела человека и животных. Труды Галена в течение 14 веков были основными источниками анатомических и медицинских знаний. Студенты медики изучали Галена до XIX в. включительно. Его теория о том, что мозг контролирует движения при помощи нервной системы, актуальна и сегодня.
В то время как в Европе созревало феодальное общество, которое тормозило развитие науки, на мусульманском Востоке наука развивалась более свободно. В VII–VIII вв. арабы вторглись в Европу, и труды ученых Древней Греции и Рима были переведены на персидский язык, что оказало большое влияние на развитие науки и культуры народов Востока.
Абу-али Ибн-Сина (Авиценна) (980–1037 гг.) — ученый и врач Востока. Написал труд «Канон врачебной науки», в котором содержались многочисленные сведения по анатомии и физиологии человека.
В период Средневековья изучение анатомии практически прекратилось, что сказалось на квалификации врачей. Невежество их настолько возросло, что в 1240 г. король Фридрих II и общественность города Салерно обратились к церкви с просьбой позволить врачам изучать анатомию. После этого было разрешено вскрывать один труп каждые 5 лет.
Дальнейшее развитие анатомии связано с открытием в Европе первых университетов. Мтудентам-медикам разрешалось вскрывать 1–2 человеческих трупа в год. В XIV–XV вв. преподавание анатомии началось в университетах Болоньи, Венеции, Вены, Монпелье и Парижа.
Мондино да Люцци (Мондинус) (1275–1327 гг.) — профессор Болонского университета, популярнейший лектор и ученый своего времени. Ему удавалось тайно вскрывать трупы людей и животных. В своем труде «Анатомия» он сделал попытку внести некоторые уточнения по анатомии, но в целом его труд был только дополнением к анатомии Галена. За два последующих века учебник переиздавался более 25 раз.
Леонардо да Винчи (1452–1519 гг.) — ученый-естествоиспытатель и выдающийся художник эпохи Возрождения. Вскрывал и изучал трупы, занимался пластической анатомией и сделал много точных рисунков мышц, костей и внутренних органов у человека и животных. Подошел к пониманию кровообращения. Он изучал пропорции тела человека и сделал попытку объяснить функции мышц с позиций механики. Его анатомические труды не были известны современникам и не оказали существенного влияния на последующие поколения медиков.
Яков Сильвий (1478–1555 гг.) — французский анатом и выдающийся лектор. Он описал латеральную борозду мозга, венозные клапаны, червеобразный отросток кишки, печень, но руководствовался идеологией учения Аристотеля и Галена. Сильвий был известным врачом, одним из сторонников учения Галена, учителем многих анатомов, в том числе и реформатора анатомии Везалия.
Период научной анатомии
Андреас Везалий (1514–1564 гг.) — бельгиец, сын придворного аптекаря Карла V. Слушал лекции Сильвия по учебнику Галена. Продолжил самостоятельное изучение анатомии, препарируя трупы. Уже в декабре 1537 г. ему присудили звание доктора медицины, разрешили публичное вскрытие трупов и преподавание анатомии и хирургии. В Падуанском университете Везалий заведовал кафедрой анатомии. С этого момента он стал излагать анатомию на основании того, что наблюдал при вскрытии, уже не доверяя авторитетам. Написал труд «О строении человеческого тела» в 7 книгах, прекрасно иллюстрированный художником Стефаном Калькаром. Эта работа переведена на русский язык в 1950–1954 гг. Работа Везалия должна была убедить весь мир в том, что схоластическая (основанная на отвлеченных рассуждениях, не проверяемых опытом) анатомия Аристотеля и Галена не может служить основой медицины. Анатомия Везалия, созданная по материалам препаровки трупов, содержала и некоторые физиологические объяснения.
Работы Везалия явились основой современной анатомии. Его данные способствовали развитию физиологии, сравнительной анатомии и эмбриологии.
Габриэль Фаллопий (1523–1562 гг.) — итальянский врач и анатом эпохи Возрождения, ученик Везалия. Дополнил работу своего учителя точным описанием мышц, костей, полукружных каналов, черепных нервов, а также маточных трубах, называемых в его честь фаллопиевыми. Полученные Фаллопием данные были изложены в книге «Анатомические наблюдения», которая получила одобрение Везалия.
Бартоломео Евстахий (1510–1574 гг.) — итальянский анатом и врач, профессор анатомии в римской школе. Один из основоположников научной анатомии, в основу которой положил сравнительно-анатомические исследования органов человека и человеческого зародыша, а также патологоанатомические вскрытия. В книге «Письма об органе слуха» (1563 г.) впервые подробно описал орган слуха человека; открыл соединительный канал между внутренним ухом и носоглоточным пространством (евстахиева, или слуховая, труба), полулунный клапан нижней полой вены.
Иероним Фабриций (1537–1619 гг.) — итальянский анатом и хирург; обогатил медицину многочисленными открытиями в области анатомии и эмбриологии и собрал богатый материал по оперативной хирургии. Был учителем Уильяма Гарвея.
Джованни Борелли (1608–1679 гг.) — итальянский ученый-энциклопедист, основоположник учения о физиологии движения. Автор трудов по физике, медицине, астрономии, геологии, математике, механике. Основоположник биомеханики. В работе «О движении животных» (1681 г.), опубликованной после его смерти, разобраны механизмы построения различных движений человека и животных. Эти данные вместе с данными об открытии кровообращения Гарвея заложили основы функционального направления в анатомии.
Уильям Гарвей (1578–1657 гг.) — английский медик, основоположник физиологии и эмбриологии. Издал небольшую книгу «Анатомическое исследование о движении сердца и крови у животных», где указал на движение крови по большому и малому кругу. Гарвею не удалось показать мелкие сосуды, соединяющие сосуды большого и малого круга кровообращения, тем не менее были созданы предпосылки для их открытия. С момента открытия Гарвея начинается подлинная научная физиология.
Фредерик Рюйш (Рёйс) (1638–1731 гг.) — знаменитый нидерландский анатом и ботаник. Усовершенствовал учение о лимфатических сосудах. Открыл методику сохранения анатомических препаратов и бальзамирования трупов, a также неизвестный в настоящее время способ наполнять тонкие кровеносные сосуды затвердевающей жидкостью.
Петр Великий, будучи в Амстердаме в 1698 г., весьма часто посещал анатомический театр Рюйша, присутствовал на его занятиях. «Петр I поддерживал отношения с Рюйшем и позже: так, в 1701 г. он послал Витзеку несколько экземпляров ящериц и червей с условием, чтобы половина из них была отдана Рюйшу; последний в благодарность послал царю несколько редких экземпляров животных Восточной и Западной Индии и в письме надавал царю наставления, как червяков кормить листьями, как прокалывать бабочек и т. д., и просил прислать из Москвы бабочек и гадин и из Азова разных зверьков и рыб».

«Урок анатомии Фредерика Рюйша» (худ. Адриан Баккер, 1670 г.)
Во время второго заграничного путешествия царь купил в 1717 г. анатомический кабинет Рюйша за 50 000 флоринов; Рюйш сообщил царю свой удивительный способ бальзамирования трупов, который раньше хотел продать Арескину за 50 000 флоринов; хотя способ был сообщен царю по секрету, но Петр передал его Блюментросту, последний — Шумахеру, а этот — лейб-медику Ригеру, который, покинув Россию, распубликовал его в Notitia rerum naturalium (статья Animal).
Находящиеся в музее Академии наук в Петербурге (Кунсткамера) препараты, приготовленные Рюйшем, превосходно сохранились до настоящего времени. Часть коллекций Рюйш продал польскому королю Станиславу, который их подарил Виттенбергскому университету.
Антони ван Левенгук (1632–1723 гг.) — нидерландский натуралист, конструктор микроскопов, основоположник научной микроскопии.
Марчелло Мальпиги (1628–1694 гг.) — итальянский биолог и врач. Используя микроскоп, описал кровеносные капилляры в легких и тем самым доказал, что артерии и вены большого и малого круга кровообращения соединяются капиллярами.
Рене Декарт (1596–1650 гг.) — французский философ, математик, физик и физиолог, создатель аналитической геометрии и современной алгебраической символики, предтеча рефлексологии. Под влиянием Гарвея Декарт считал, что от мозга радиально расходятся нервные «трубки», несущие автоматически отражения к мышцам.
Говерт Бидлоо (1649–1713 гг.) — ученик Рюйша, известный анатом. Показал, что нервные стволы состоят из массы тонких волокон и не содержат жидкости (против господствующего мнения о spiritus anima-lis).
Мари Франсуа Ксавье Биша́ (1771–1802 гг.) — выдающийся французский анатом, физиолог и врач, отец современной гистологии и патологии. Является одним из основоположников современной танатологии (раздел медицины, занимающийся динамикой и механизмом умирания). Биша принадлежит одно из определений понятия жизнь: «совокупность отправлений, противящихся смерти». Он первым ввел понятие тканей как независимых сущностей. Биша утверждал, что болезни атакуют ткани, а не органы целиком. Первым обратил внимание на наличие жировых тел между жевательной и щечной мышцами, впоследствии получивших его имя (комочки Биша́).
Карл Бэр (1792–1876 гг.) — один из основоположников эмбриологии и сравнительной анатомии. Открыл яйцеклетку человека.
Теодор Шванн (1810–1882 гг.) — немецкий цитолог, гистолог и физиолог, автор клеточной теории.
Иоганн Блюменбах (1752–1840 гг.) — известный немецкий анатом, антрополог и естествоиспытатель. Описал современные расы человека и высказал мысль об единстве их происхождения.
Вильгельм Рентген (1845–1923 гг.) — крупнейший немецкий физик-экспериментатор. Открыл (1895 г.) рентгеновские лучи, что привело к созданию рентгеноанатомии.
Джон Ленгли (1852–1925 гг.) — английский физиолог и гистолог. Основные труды по анатомии и физиологии вегетативной нервной системы. Открыл ее автономию и отделы.
развитие отечественной анатомии
Мартын Иванович Шеин (1712–1762 гг.) — русский анатом. Создал и иллюстрировал первый отечественный атлас анатомии «Словник, или иллюстрированный указатель всех частей человеческого тела» на латинском языке. Он же в 1757 г. впервые перевел на русский язык «Сокращенную анатомию, все дело анатомическое кратко в себе заключающую» Лаврентия Гейстера (Heister, Laurentii), которая стала первым в России практическим руководством по анатомии на русском языке. Он заложил основы русской анатомической терминологии.

Иллюстрация М. И. Шеина
Петр Андреевич Загорский (1764–1846 гг.) — выдающийся анатом, эволюционист. Основоположник самостоятельной отечественной анатомической школы. Первый в России заложил основы экспериментальной и сравнительной физиологии. Утверждал русскую анатомическую терминологию взамен латинской.
Здесь важно отметить, что анатомия как наука и предмет преподавания не выделялась в самостоятельную дисциплину вплоть до начала XIX в. Она объединялась с физиологией, патологией, а позднее и с патологической физиологией и изучалась в тесной связи с хирургией. Таким образом, выдающиеся анатомы того времени были одновременно и блистательными хирургами, терапевтами, физиологами.
Семен Герасимович Зыбелин (1735–1802 гг.) — российский медик, один из первых русских профессоров медицины в Московском университете. Был известен как один из самых красноречивых лекторов. Ведя преподавание на русском языке (а не на латыни, как было принято), Зыбелин способствовал созданию правильного и точного языка для врачебной науки. Он ввел на лекциях демонстрацию опытов. В 1771 г. был одним из организаторов борьбы с эпидемией чумы в Москве.
Александр Михайлович Шумлянский (1748–1795 гг.) — врач, один из основоположников микроскопической анатомии в России, профессор Московской акушерской школы. Основной труд по вопросам строения и функций почек. Открыл капсулу Шумлянского (капсула Шумлянского–Боумена).
Илья Васильевич Буяльский (1798–1866 гг.) — ученик П. А. Загорского и его преемник по кафедре анатомии Петербургской медико-хирургической академии. Опубликовал «Анатомико-хирургические таблицы», сразу получившие мировую известность. Они состояли из 36 рисунков и 14 таблиц, изображавших органы в натуральную величину. Их ценность определялась также и тем, что существовавшие в то время учебник анатомии П. А. Загорского и учебник хирургии И. Ф. Буша иллюстраций не имели. Таблицы И. В. Буяльского объединили в себе данные топографической анатомии и оперативной хирургии и явились первым в истории России отечественным атласом по оперативной хирургии.
И. В. Буяльский был искусным анатомом и блестящим хирургом. Он много сделал для создания новых хирургических инструментов, разработал методы бальзамирования трупов, предложил новые способы изготовления тонких коррозионных анатомических препаратов. В числе первых русских хирургов применил наркоз, крахмальную повязку и средства антисептики.
Николай Иванович Пирогов (1810–1881 гг.) — русский хирург и анатом, естествоиспытатель и педагог, создатель первого атласа топографической анатомии, основоположник русской военно-полевой хирургии, основатель русской школы анестезии. Ввел метод «ледяной» анатомии и распилов замороженных трупов. Основатель топографической (хирургической) анатомии.
Петр Францевич Лесгафт (1838–1909 гг.) — основоположник теоретической анатомии и зачинатель отечественной науки о физическом воспитании. По Лесгафту, форма непрерывно изменяется под влиянием функций и условий питания, следовательно, совершенная форма здорового организма находится в прямой зависимости от активного воздействия на него упражнений, разработанных на научной основе. Таким образом, П. Ф. Лесгафт впервые проложил мост между анатомией и физическим воспитанием и образованием людей.
Николай Петрович Гундобин (1860–1908 гг.) — русский врач-педиатр. Основные труды посвящены изучению возрастных особенностей анатомии, физиологии и патологии детского организма.
Владимир Петрович Воробьев (1876–1937 гг.) — российский и советский анатом. Исследовал преимущественно анатомию нервной системы. Предложил метод прижизненного контроля с помощью вшитых электродов. Развил учение о целостности организма, о влиянии функций и труда на морфогенез (формирование органов). Впервые определил значение пограничной макромикроскопической области видения и разработал методы ее исследования.
Виктор Валерианович Бунак (1891–1979 гг.) — выдающийся русский антрополог, один из основоположников советской антропологической школы.
Иван Михайлович Сеченов (1829–1905 гг.) — российский ученый, создатель физиологической школы. В классическом труде «Рефлексы головного мозга» (1866 г.) обосновал рефлекторную природу сознательной и бессознательной деятельности, показал, что в основе психических явлений лежат физиологические процессы.
Иван Петрович Павлов (1849–1936 гг.) — российский физиолог, создатель учения о высшей нервной деятельности. Автор классических трудов по физиологии кровообращения и пищеварения (Нобелевская премия, 1904 г.). Ввел в практику хронический эксперимент, позволяющий изучать деятельность практически здорового организма.
Илья Ильич Мечников (1845–1916 гг.) — российский биолог и патолог, один из основоположников сравнительной патологии, эволюционной эмбриологии и отечественной микробиологии, иммунологии. Создатель учения о фагоцитозе и теории иммунитета.
Науки, изучающие организм человека
Морфология
Морфология человека — наука о внешнем строении организма человека, связи строения с функциями и о закономерностях изменчивости отдельных его частей.
Морфология человека тесно связана с происхождением человека и с местом человека в системе животного мира. Включает 2 раздела:
Соматология («сома» — тело) изучает закономерности индивидуальной изменчивости человеческого организма в целом, половые различия (половой диморфизм), возрастные изменения и влияние условий жизни на строение организма и отдельных органов.
Данный раздел имеет прикладное значение, т. к. тесно связан с медициной и геронтологией (наукой о старении).
Мерология изучает изменчивость отдельных частей организма человека.
Для изучения используются сравнительно-анатомические методы с целью выявления черт сходства и различия с организмами других приматов.
Возрастная морфология изучает закономерности роста и развития организма.
Анатомия
Анатомия человека — (греч. «рассечение») наука о внутреннем строении человека и отдельных органов.
Нормальная анатомия — анатомия здорового тела человека.
Сравнительная анатомия — раздел анатомии, изучающий общие закономерности строения органов при помощи их сравнения у разных таксонов животных.
Топографическая анатомия — прикладной раздел анатомии, изучающий расположение органов в пространстве. Используется в хирургической практике. Основана Н. И. Пироговым.
Функциональная анатомия — раздел анатомии, рассматривающий связь строения организма с его функциями.
Пластическая анатомия — раздел анатомии, изучающий внешнюю форму тела человека и его пропорции. Основателем данной науки считается Леонардо да Винчи.
Патологическая анатомия — научно-прикладная дисциплина, изучающая патологические болезненные процессы в организме.
Макроскопическая анатомия — (греч. «макро» — большой) изучает строение тела, отдельных органов и их частей на уровнях, доступных невооруженному глазу, или при помощи приборов, дающих небольшое увеличение (лупа).
Микроскопическая анатомия (греч. «микро» — малый) изучает строение органов при помощи микроскопа.
С появлением микроскопов из анатомии выделилась:
гистология — наука о животных тканях;
цитология — наука о клетках.
Методы изученияя анатомии:
наблюдение;
осмотр тела (соматоскопия);
препарирование (вскрытие) — метод основан на рассечении мертвых тканей;
метод Н. И. Пирогова — метод распила замороженного тела (ледяная анатомия);
метод инъекции — введение окрашивающих веществ;
метод коррозии — метод исследования полых органов путем введения застывающего вещества и последующим растворением тканей в кислотах или щелочах с целью получения слепка внутреннего строения органа;
макро-микроскопический метод — метод тонкого препарирования и исследования тканей с помощью лупы (разработал метод В. П. Воробьев);
микроскопические методы — исследования с помощью микроскопа;
эндоскопический метод — способ осмотра полых внутренних органов при помощи эндоскопа (рис. 1);
 Рис.
1
Рис.
1
рентгенография (рентген) — метод изучения внутренних органов с помощью рентгеновских лучей (рис. 2, 3);

 Рис.
2
Рис.
3
Рис.
2
Рис.
3
ультразвуковое исследование (УЗИ) — исследование организма человека с помощью ультразвуковых волн (рис. 4, 5);

 Рис.
4
Рис. 5
Рис.
4
Рис. 5
компьютерная томография — метод получения послойного изображения объекта с помощью рентгеновских лучей с последующей компьютерной обработкой снимков (рис. 6);
магнитно-резонансная томография (МРТ) — метод получения послойного изображения объекта без использования рентгеновских лучей (рис. 7).


Рис. 6 Рис. 7
Физиология
Физиология человека — наука о функциях человеческого организма и его отдельных органов.
Нейрофизиология — раздел физиологии человека и животных, изучающий функции нервной системы.
Возрастная физиология — раздел физиологии человека и животных, изучающий закономерности развития функций организма на протяжении онтогенеза (индивидуального развития организма) — от оплодотворения яйцеклетки до конца жизни.
Сравнительная физиология — раздел физиологии, изучающий методом сравнения различия физиологических функций у разных животных.
Эволюционная физиология изучает изменение физиологических функций организмов в процессе эволюционного развития.
Экологическая физиология изучает изменчивость физиологических реакций организма под влиянием экологических факторов.
Методы физиологических исследований:
исследования выделенных органов в питательной среде;
хронические эксперименты на живых животных (выведение фистулы для наблюдения желудочной секреции (метод И. П. Павлова), вживление электродов для изучения мозговой активности) (рис. 8, 9);

 Рис.
8
Рис. 9
Рис.
8
Рис. 9
эндоскопический метод;
метод электрокардиографии (ЭКГ) — метод исследования электрической активности сердца (рис. 10, 11);

 Рис.
10
Рис.
11
Рис.
10
Рис.
11
ультразвуковое исследование;
лабораторные анализы — биохимический состав крови и других жидкостей;
компьютерное моделирование работы органов.
Анатомия и физиология дополняют друг друга и служат основой для медицины и гигиены.
Гигиена
Гигиена — раздел медицины, изучающий влияние условий жизни и труда на здоровье человека, разрабатывающий меры по профилактике заболеваний и созданию условий для сохранения и укрепления здоровья.
В зависимости от предмета изучения гигиеническая наука подразделяется:
Гигиена окружающей среды — раздел гигиены, изучающий влияние окружающей среды на здоровье человека.
Гигиена питания и здоровья — раздел гигиены, изучающий влияние образа жизни на здоровье человека.
Гигиена труда — раздел гигиены, изучающий влияние условий труда на здоровье человека.
Социальная гигиена — раздел гигиены, изучающий влияние общества на здоровье человека.
На организм человека воздействуют факторы внешней среды:
химические — химические элементы и соединения, находящиеся в воздухе, воде, почве, пище;
физические — температура, влажность и давление воздуха, радиация, вибрация, шум и т. п.;
биологические — микроорганизмы, гельминты, грибы;
социальные факторы — характер и условия трудовой деятельности, питание, жилищно-бытовые условия и др.
Задача гигиены — снизить вредное влияние факторов среды на здоровье человека.
МЕТОДЫ ГИГИЕНЫ:
метод санитарного описания и обследования (определение санитарного состояния объекта);
метод клинических исследований (выяснение причин массовых заболеваний);
физиологические (установление приспособительных возможностей организма к неблагоприятным условиям природной и социальной среды);
метод лабораторных исследований (физические, химические, биологические методы исследования);
санитарно-статистический метод (привлечение статистических данных);
демографический метод (исследования динамики численности, рождаемости, смертности, заболеваемости населения);
экспериментальные методы (для изучения влияние среды на организм).
Санитария — практическое применение гигиенических нормативов, правил и мероприятий.
Достижения гигиены внедряются путем общественно-гигиенических мероприятий. С этой целью разрабатываются различные санитарные правила и нормы. Для эффективного использования достижений гигиены необходимо, чтобы каждый человек понимал их значение. В связи с этим огромная роль принадлежит санитарному просвещению.
Санитарно-эпидемиологическая служба (СЭС) осуществляет государственный надзор и контроль за исполнением обязательных требований российского законодательства в сфере обеспечения санитарно-эпидемиологического благополучия населения, защиты прав потребителей и в области потребительского рынка. В настоящее время СЭС носит другое название — Федеральная служба по надзору в сфере защиты прав потребителей и благополучия человека РФ, или просто Роспотребнадзор.
Карантин — запрещение или ограничение на въезд или выезд на территорию предполагаемого заражения.
Бактериальные и вирусные заболевания человека. Их профилактика
Многие организмы, относящиеся к разным систематическим группам, являются паразитами человека и вызывают заболевания. Заболевания, возбудители которых являются микроскопическими (вирусы, бактерии и простейшие), называются инфекционными.
В зависимости от природы возбудителей различают разные типы инфекционных заболеваний:
Вирусные, например грипп, ОРВИ, корь, оспа, ВИЧ-инфекция, вирусные гепатиты, клещевой энцефалит, желтая лихорадка.
Бактериальные, например чума, холера, столбняк, сибирская язва, стрептококковая и стафилококковая инфекции, коклюш.
Протозойные (вызываемые простейшими, то есть одноклеточными эукариотами), например малярия, сонная болезнь, амебная дизентерия, токсоплазмоз.
Грибковые, например кандидоз, эпидермофития (грибок стопы).
Прионные (вызываемые особыми инфекционными белками) — «коровье бешенство» (BSE), куру.
Возбудители разных групп чувствительны к разным лекарственным веществам. Поэтому медикаментозное лечение заболевания может проводиться только после диагностики природы заболевания и понимания природы его возбудителя. Так, большинство антибиотиков действуют на клетки бактерий, тогда как вирусы являются своеобразной неклеточной формой жизни. Поэтому бессмысленно лечить вирусные заболевания антибиотиками. Для борьбы с вирусами применяют противовирусные средства (следует отметить, что высокоспецифичные и эффективные противовирусные средства появились относительно недавно и существуют далеко не для всех вирусных заболеваний).
история изучения инфекционных заболеваний
природа возбудителей
Представление о заразности болезней, в частности таких как чума, оспа, холера, зародилось еще у древних народов. Простейшие меры предосторожности в отношении заразных больных применялись еще до нашей эры. Однако эти догадки были еще далеки от научных знаний о природе заболевания.
Уже в Древней Греции некоторые философы высказывали мысль о живых возбудителях инфекционных болезней, но эти ученые не могли подтвердить свои предположения какими-либо достоверными фактами. Выдающийся древнегреческий врач Гиппократ (около 460–377 гг. до н. э.) объяснял происхождение эпидемий действием «миазм» — заразных испарений, которые будто бы могут вызвать ряд болезней.

Антони ван Левенгук (голландская почтовая марка)
Переворот в понимании природы заболеваний связан с возникновением и распространением микроскопии. Голландец Антони ван Левенгук в конце XVII в. изготовил микроскоп и обнаружил различные микроорганизмы в зубном налете, в воде из лужи и настое растений. Левенгук не первым изготовил микроскоп, считается, что первыми были отец и сын Янсены в 1590 г. Однако микроскоп Левенгука был им существенно усовершенствован и давал гораздо больше возможностей, чем более примитивные микроскопы предшественников. Так был открыт невидимый глазу мир микробов, многие из которых могли, очевидно, являться возбудителями болезней. Левенгук описал свои наблюдения в книге «Тайны природы, открытые Антонием Левенгуком при помощи микроскопов». Но даже после его открытия идея о микробах как возбудителях инфекционных болезней долгое время не получала еще необходимого научного обоснования, хотя эпидемии уносили тысячи человеческих жизней по всей Европе. Наблюдения за эпидемиями убеждали людей в заразности заболеваний. Но только в 1840–1850-х гг. были впервые открыты патогенные для человека микробы.

Луи Пастер
Великий французский ученый Луи Пастер (1822–1895 гг.) в своих работах доказал участие микроорганизмов в процессах брожения и гниения, а также в развитии инфекционных болезней. Работы Пастера объяснили природу инфекционных болезней человека, они легли в основу асептики и антисептики (см. ниже), разработанных в хирургии Н. И. Пироговым и Дж. Листером, а также их последователями и учениками, что привело к революционному прорыву в развитии хирургии.
Начиная со второй половины XIX в. медицинская микробиология получила быстрое развитие. В начале XX в. уже сложились четкие научные представления о природе, механизмах заражения и путях передачи ряда инфекционных заболеваний и оказалось возможным осуществлять гигиенические мероприятия, направленные на предупреждение этих болезней.

Илья Ильич Мечников
И. И. Мечников (1845–1916 гг.) открыл фагоциты, участвующие в обеспечении иммунной защиты организма, и положил начало исследованиям иммунитета (невосприимчивости) при инфекционных болезнях, была показана исключительно важная роль клеточной (фагоцитоз и др.) и гуморальной (антитела) защиты организма.
Помимо чисто клинического исследования инфекционных больных для диагностики отдельных заболеваний с конца XIX в.стали широко применять лабораторные методы, такие как культивирование и микроскопия возбудителей. Также неоценимый вклад в эпидемиологию внесли эксперименты по заражению лабораторных животных. Но некоторые исследователи ставили эксперименты на самих себе. Задолго до появления выдающихся исследований Пастера, осветивших роль микробов в человеческой патологии, наш соотечественник С. С. Андреевский (1786 г.) самоотверженным опытом прививки самому себе содержимого сибиреязвенного карбункула от больного животного доказал, что сибирскую язву у людей и домашних животных вызывает одна и та же причина. Врачи Одесской городской больницы О. О. Мочуткозский и Г. Н. Минх в 1870-х гг. для того, чтобы доказать, что заразное начало при сыпном и возвратном тифах находится в крови больного, впрыснули себе под кожу кровь больных и перенесли затем тяжелое заболевание.
антисептика и асептика
Интересно, что Пастер, который не был врачом, совершенно правильно оценил значение своего открытия для медицины. В обращении к членам Парижской академии хирургии в 1878 г. он сказал: «Если бы я имел честь быть хирургом, то сознавая опасность, которой грозят зародыши микробов, имеющиеся на поверхности всех предметов, особенно в госпиталях, я бы не ограничивался заботой об абсолютно чистых инструментах; перед каждой операцией я сперва бы тщательно промывал руки, а затем держал бы их в течение секунды над пламенем горелки; корпию, бинты и губки я предварительно прогревал бы в сухом воздухе при температуре 130–150 ºC; я никогда бы не применял воду, не прокипятив ее».
Антисептика (лат. «анти» — против, «септикус» — гниение) — система мероприятий, направленных на уничтожение микроорганизмов в ране, патологическом очаге, органах и тканях, а также в организме больного в целом, использующая механические и физические методы воздействия, активные химические вещества и биологические факторы. Основоположником антисептики считают английского хирурга Джозефа Листера (1829–1912 гг.), который, ознакомившись с работами Пастера, пришел к выводу, что микроорганизмы попадают в рану из воздуха и с рук хирурга. Убедившись в антисептических свойствах карболовой кислоты, он применил повязку с ее раствором в лечении открытого перелома и в дальнейшем использовал для обеззараживания оборудования и раны. До появления антисептики хирурги практически никогда не шли на риск операций, связанных со вскрытием полостей человеческого тела, так как вмешательства в них сопровождались почти стопроцентной летальностью от хирургических инфекций. Профессор Эрикоен, учитель Листера, в 1874 г. заявлял, что брюшная и грудная полости, а также полость черепа навсегда останутся недоступными для хирургов.
Асептика — комплекс мероприятий, направленных на предупреждение попадания микроорганизмов в рану. Асептику следует отличать от антисептики, которая имеет целью уничтожить возбудителей воспаления, уже имеющихся в ране. Одним из основателей асептики считается немецкий хирург Эрнст фон Бергманн. Он предложил физические методики обеззараживания — кипячение, обжигание, автоклавирование. Основой асептики является стерилизация — уничтожение микроорганизмов и их спор. Не следует путать стерилизацию с дезинфекцией, при которой споры микробов не уничтожаются. Существуют различные методы стерилизации: автоклавирование (обработка паром под давлением), прокаливание или обжигание огнем, погружение в антисептические растворы, обработка ионизирующими излучениями (например, на заводах по производству шприцов и другого одноразового оборудования).
история лекарств
Важный раздел учения об инфекционных болезнях — разработка методов их активного лечения. Наука о лекарственных веществах и их воздействии на организм называется фармакологией. В основе фармакологии лежит применение лекарственных растений и других природных веществ для лечения заболеваний, которое практиковали люди всей земли с древнейших времен. Традиционные способы лечения при этом вырабатывались на основе опыта, методом проб и ошибок, и далеко не все из них имели под собой какую-либо научную основу. Стандартизированные, отвечающие всем научным критериям процедуры клинических испытаний, необходимые для того, чтобы какое-либо вещество в наши дни применялось как лекарство, возникли относительно недавно. Многие из народных способов лечения основаны на религиозных и мистических представлениях и не имеют под собой никаких оснований. Однако некоторые получили в дальнейшем научное подтверждение. Так, для лечения больных малярией применяли хинную кору еще индейцы кечуа, от которых о ней узнали европейцы, а с 1821 г. стали использовать алкалоид хинин — первое из химиотерапевтических средств, полученных синтетическим путем. Хинин применялся для профилактики и лечения малярии. Однако развитие фармакологии задержалось надолго. Лишь с 1909 г. в лечебной практике появились препараты мышьяка (сначала арсацетин, затем сальварсан, неосальварсан и др.), полученные в Германии, которые затем с успехом применялись для лечения возвратного тифа, сифилиса, сибирской язвы и некоторых других инфекционных болезней.
Еще в 1871 г. известный русский ученый В. А. Манассеин, а вслед за ним А. Г. Полотебнов изучили ряд свойств зеленого плесневого грибка (из рода Реnicillium), подметив его антагонистическое влияние на некоторые бактерии. А. Г. Полотебнов, кроме того, применил зеленый плесневый грибок для лечения гуммозных язв и гнойных ран.
|
|
|
Александр Флеминг |
Ховард Флори |
Эрнст Чейн |



Исключительно важную роль сыграли наблюдения английского микробиолога Александра Флемминга (1881–1954 гг.), который в 1929 г. обнаружил, что фильтраты среды, в которой росла зеленая плесень Penicillium notatum, обладают способностью подавлять рост стафилококка и ряда других бактерий. Основываясь на этих фактах и разработав сложную технологию извлечения активных веществ из культуры указанной плесени, Говард Флори и Эрнст Чейн в 1941 г. в Оксфорде получили антибиотик пенициллин. В 1945 г. Флеминг, Флори и Чейн были удостоены Нобелевской премии в области физиологии и медицины. Еще в 1930-х гг. были получены синтетические антибактериальные препараты сульфаниламидного ряда, правда, первые лекарства этой группы имели много побочных эффектов. Пенициллин переносился лучше. После открытия пенициллина появились и другие антибиотики: в 1944 г. — стрептомицин, в 1948 г. — хлоромицетин, хлорамфеникол и их аналоги.
В настоящее время антибиотики являются основным средством лечения ряда инфекционных болезней. Изобретение антибиотиков кардинально изменило жизни людей всего мира, привело к существенному увеличению средней продолжительности жизни, позволило победить страшные эпидемии бактериальных заболеваний, предотвратить гибель женщин при родах, солдат при ранениях. К сожалению, бактерии быстро мутируют и приобретают устойчивость к антибиотикам. Проблема устойчивости к антибиотикам является одной из важнейших в современной медицине. Одним из направлений развития фармакологии является разработка новых антибиотиков. Для предотвращения развития устойчивости ни в коем случае нельзя применять антибиотики без диагностики и врачебного назначения, следует четко следовать предписаниям по дозировке и режиму приема лекарственного средства, нельзя злоупотреблять антибиотиками. При подозрении на устойчивость к антибиотикам в некоторых случаях может быть осуществлен диагностический посев возбудителя на среды с различными антибиотиками, чтобы выявить те, к которым он чувствителен.
вакцины и вакцинация
Одним из важнейших средств предотвращения заболеваний сегодня является вакцинация, или прививки. Прививки — это не лекарства в обычном смысле. Ими не лечат заболевание, а предотвращают. Действие прививок основано на природной способности организма приобретать иммунитет, то есть устойчивость к возбудителю после перенесенного заболевания. К каким-то из заболеваний иммунитет сохраняется всю жизнь, к каким-то — на определенный период времени. Вакцина — биопрепарат, содержащий убитых возбудителей болезни или их компоненты, либо живой, но ослабленный штамм возбудителя, не способный вызвать болезнь. При попадании в организм вакцины начинается иммунный ответ на чужеродные белки и другие биомолекулы, в результате чего вырабатываются антитела — естественные защитные белки организма, специфичные именно к данному конкретному возбудителю. В-лимфоциты, продуцирующие антитела, могут сохраняться в организме долгие годы. Если в организме вырабатывается достаточное количество антител к возбудителю, человек устойчив к заболеванию или переносит его в легкой форме.
Страшным бичом человечества в древности и Средние века был вирус оспы. Тысячи людей по всему миру умирали при эпидемиях этой болезни. Пытаясь защититься от нее, еще в Средневековье в Китае и Персии практиковали вариоляцию — прививку оспенного гноя здоровым людям, что приводило к заболеванию оспой в легкой форме и приобретению иммунитета к этой болезни. Конечно, при этом у многих людей развивалась полная форма заболевания, и нередко они умирали. Но другая часть приобретала спасительный иммунитет. Так как других способов борьбы со смертоносным вирусом все равно не было, вариоляцию продолжали практиковать и в Европе (в 1718 г. эта практика была привезена британским послом из Турции в Англию). В XVIII в. наблюдатели обратили внимание на коровью оспу, неопасную для человека. Доярки, а также кавалеристы, которые часто переносили коровью оспу, впоследствии не заражались натуральной оспой. В 1796 г. английский врач и ученый Эдуард Дженнер (1749–1823 гг.) привил 8-летнему мальчику коровью оспу, после чего у мальчика развилось лишь легкое недомогание, которое быстро прошло, а затем в том же году привил ему натуральную оспу. Натуральной оспой мальчик не заболел. Через несколько месяцев и 5 лет натуральная оспа была привита снова, с тем же результатом. Таким образом Дженнер доказал эффективность вакцинации. В дальнейшем были разработаны вакцины от множества заболеваний. Большой вклад в развитие вакцинации внес Луи Пастер, который разработал методы ослабления вирулентных свойств возбудителей путем особого подбора соответствующих условий для их культивирования. Пастером были получены вакцины для прививок против сибирской язвы и бешенства. Вакцинация открыла новую эру в истории человечества, позволив победить или обуздать многие страшные болезни.
Еще в первой половине XX в. от оспы ежегодно умирали миллионы людей. Всемирная кампания по борьбе с оспой была начата по инициативе СССР в 1958 г. В 1967 г. Всемирная организация здравоохранения (ВОЗ) принимает решение об интенсификации искоренения натуральной оспы с помощью массовой вакцинации человечества. Благодаря этой кампании оспа была побеждена во всем мире. Сейчас образцы вируса хранятся только в двух лабораториях, в Америке и в России.
Здравоохранение в нашей и других странах предполагает систему обязательных прививок. Проводится обязательная вакцинация от таких болезней, как дифтерия, коклюш, столбняк, туберкулез, полиомиелит, корь. Проведение вакцинации в полном объеме является важным фактором защиты не только конкретного человека, но и всего населения в целом и предотвращает эпидемии. Кроме того, необходимо проводить вакцинацию перед поездкой в определенные эпидемиологически опасные районы. В связи с участившимися отказами от прививок по религиозным и другим соображениям в последние годы в развитых странах участились вспышки таких заболеваний, как корь и коклюш, с эпицентрами в религиозных общинах, где проживают невакцинированные люди. Важно понимать уровень своей ответственности за жизнь и здоровье не только своих детей, но и общества в целом.
Строение клеток эукариот. Одномембранные органеллы. Ядро
Эукариотическая клетка отличается от прокариотической большими размерами и более сложным строением. Линейные размеры эукариотической клетки обычно составляют десятки мкм (у животных около 10–40 мкм, у растений 100–200 мкм).
Основные структуры эукариотической клетки: цитоплазматическая мембрана, цитоплазма и ядро.
Клетки эукариот содержат множество внутренних структур, выполняющих определенные функции (рис. 1). Эти структуры называются органеллами.

Рис. 1. Строение эукариотической клетки
цитоплазма
Цитоплазма — обязательная часть клетки, заключенная между плазматической мембраной и ядром. Цитоплазма объединяет все клеточные структуры и способствует их взаимодействию друг с другом.
Цитоплазма заполнена взвесью частиц и органелл.
Цитозоль — свободная часть цитоплазмы, в которой взвешены органеллы.
Цитозоль — густой раствор белков, углеводов (глюкозы) и ионов, заполняющий все внутреннее пространство клетки.
Концентрации ионов натрия и калия в цитозоле (т. е. внутри клетки) отличаются от таковых во внеклеточном пространстве. Эти различия в концентрациях ионов играют важную роль в осморегуляции и передаче сигнала.
У прокариот большинство реакций метаболизма протекает в цитозоле, т. к. нет мембранных органоидов.
У эукариот часть химических реакций протекает в цитозоле, а часть — внутри органоидов.
В животных клетках различают два слоя цитоплазмы:
эктоплазма — наружный слой цитоплазмы (мало органоидов, высокая вязкость);
эндоплазма — внутренний слой цитоплазмы (содержит основные органоиды).
В цитоплазме осуществляются все процессы клеточного метаболизма, кроме синтеза нуклеиновых кислот, происходящего в ядре. Под контролем ядра цитоплазма способна к росту и воспроизведению, при частичном удалении она полностью регенерирует. Цитоплазма, как правило, не способна к длительному автономному существованию.
Циклоз — одно из основных свойств цитоплазмы живой клетки — способность к движению, которое обеспечивает транспорт веществ и связь органоидов.
ядро
Это крупная органелла около 6–7 мкм диаметром (рис. 2). Оно окружено ядерной оболочкой, которая образована двумя параллельно расположенными мембранами. Ядерная оболочка пронизана ядерными порами, где мембраны смыкаются, и полость ядра сообщается с цитоплазмой. В порах находятся сложные белковые комплексы. Они переносят через оболочку из ядра в цитоплазму крупные молекулы и молекулярные комплексы, такие как мРНК и рибосомы, а из цитоплазмы в ядро — ядерные белки, которые синтезируются в цитоплазме. Внутри ядра находится одно или несколько ядрышек — плотных образований, где происходит синтез рибосомных РНК и сборка субъединиц рибосом. Остальное пространство ядра заполнено полужидкой кариоплазмой, в которой находятся молекулы ДНК, соединенные со специфическими белками, — хроматин.
 Рис.
2. Ядро. Трансмиссионный электронный
микроскоп. Внутри ядра видны ядрышко и
хроматин. Оболочка ядра двухмембранная.
Вокруг расположены цистерны ЭПС и
митохондрии
Рис.
2. Ядро. Трансмиссионный электронный
микроскоп. Внутри ядра видны ядрышко и
хроматин. Оболочка ядра двухмембранная.
Вокруг расположены цистерны ЭПС и
митохондрии

Рис. 3. Строение хромосомы
В процессе клеточного деления нити хроматина укорачиваются и утолщаются, превращаясь в хромосомы (рис. 3) Перед делением хромосомы имеют Х-образную форму. Центральная часть, в которой соединяются две половины хромосомы, носит название центромеры, или первичной перетяжки. Кроме того, в хромосоме выделяются более плотные концевые участки, называемые теломерами. Различные хромосомы отличаются размерами и положением центромер. Для каждого вида живых организмов характерен определенный набор хромосом, который отличается от наборов других видов. Видоспецифичный набор хромосом со всеми их характеристиками называется кариотипом (рис. 4). Клетки могут содержать один набор хромосом или их кратное число. Число хромосомных наборов называется плоидностью. Клетки, содержащие один набор хромосом, называются гаплоидными, содержащие два набора — диплоидными, три набора — триплоидными, четыре набора — тетраплоидными и т. д. Чаще всего мы имеем дело с организмами, состоящими из диплоидных клеток: это животные и большинство растений. Встречаются организмы, построенные из гаплоидных клеток, например мхи. Организмы, в клетках которых более двух наборов хромосом, принято называть полиплоидами.

Рис. 4. Кариотип
плазматическая мембрана

Рис. 5. Цитоплазматическая мембрана
Снаружи эукариотическая клетка, как и прокариотическая, окружена цитоплазматической мембраной (рис. 5). Она выполняет те же функции, что и у прокариот: изолирующую, транспортную и рецепторную. Рецепторная функция у эукариотических клеток развита гораздо сильнее, чем у прокариот, поэтому в цитоплазматической мембране у них гораздо больше белков-рецепторов. У многоклеточных организмов цитоплазматическая мембрана выполняет также функцию межклеточного узнавания и взаимодействия. У растений и грибов снаружи от цитоплазматической мембраны лежит клеточная стенка. У растений она построена на основе целлюлозы, а у грибов — на основе хитина. У животных клеточной стенки нет, но к мембране снаружи прикрепляется довольно толстый слой специфических полисахаридов и белков, называемый гликокаликс. В отличие от клеточной стенки, он эластичен, что позволяет клеткам менять свою форму. В отличие от клеточной стенки, гликокаликс прочно связан с мембраной и не отделяется от нее.
Гликокаликс и его функции
Все органеллы эукариотической клетки можно условно разделить на три группы:
Одномембранные, стенка которых образована одной мембраной. К ним относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, секреторные пузырьки, пероксисомы.
Двумембранные, стенка которых образована двумя мембранами. Это митохондрии и пластиды (хлоропласты, хромопласты и лейкопласты).
Немембранные. Это цитоскелет, клеточный центр, рибосомы.
одномембранные органеллы
Эти органеллы, как предполагается, в ходе эволюции образовались путем впячивания наружной мембраны внутрь и отпочковывания этих впячиваний. Почти все эти органеллы связаны между собой — прежде всего системой пузырькового (везикулярного) транспорта, когда пузырьки отпочковываются от одной органеллы и сливаются с другой, перенося содержимое и компоненты мембраны. Все вместе эти органеллы называются вакуолярной системой (эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, секреторные пузырьки, пероксисомы). Ядерная оболочка, по сути, тоже является частью вакуолярной системы — одной из цистерн ЭПС, которая «охватывает» наследственный материал. На наружной мембране часто сидят рибосомы, как и на шероховатой ЭПС. Внутренняя поверхность внутренней мембраны ядра имеет специфический состав и взаимодействует с ДНК внутри ядра.
Одна из гипотез происхождения ядра (рис. 6).

Рис. 6. Формирование ядра
Эндоплазматическая сеть

Рис. 7. Эндоплазматическая сеть

Рис. 8. Эндоплазматическая сеть в клетке легкого, трансмиссионная электронная микроскопия. Справа видно ядро с ядрышком, между цистернами ЭПС — митохондрии
Внутри эукариотической клетки мы видим сложные мембранные системы, образующие клеточные органеллы. Прежде всего, это эндоплазматическая сеть, или эндоплазматический ретикулум (рис. 7, 8). Он представляет собой систему мембран, образующих соединенные между собой цистерны, полость которых не сообщается с окружающей цитоплазмой. Различают два вида эндоплазматического ретикулума: гладкий и шероховатый. На шероховатом расположены многочисленные гранулы, представляющие собой рибосомы. Они находятся снаружи полости, с цитоплазматической стороны, и синтезируют белки, которые по специальному каналу сразу направляются в полость ретикулума или встраиваются в его мембрану.
На гладком ретикулуме расположены ферменты, синтезирующие мембранные липиды. Таким образом, эндоплазматический ретикулум образует все компоненты, нужные для образования мембран (то есть роста их площади). От эндоплазматического ретикулума отделяются мембранные пузырьки, внутри которых белки, синтезированные на шероховатом ретикулуме, переносятся в следующую органеллу — аппарат, или комплекс, Гольджи.
Аппарат Гольджи
Аппарат, или комплекс, Гольджи — система уплощенных мембранных цистерн, основная функция которых — сортировка и модификация (прежде всего гликозилирование) белков, направляемых на экспорт из клетки или встроенных в мембрану (рис. 9). Каждая группа белков, синтезированных на шероховатом ретикулуме, собирается в определенном участке на периферии аппарата Гольджи. В этих участках от него отделяются мембранные пузырьки, часть из которых дает начало клеточным органеллам, таким как лизосомы. Другая направляется к цитоплазматической мембране, сливается с ней и выделяет свое содержимое наружу. Таким образом осуществляется секреция из клетки таких белков, как пищеварительные ферменты, гормоны, белки межклеточного матрикса и гликокаликса.

Рис. 9. Аппарат Гольджи. Слева — трансмиссионная электронная микроскопия, справа — модель
лизосомы
Лизосомы представляют собой мембранные пузырьки, внутри которых находятся гидролитические ферменты, расщепляющие белки, жиры, полисахариды. В лизосомах кислая среда (рН 4,5–5,0), что отличает их от других органелл клетки. Эта среда создается действием специального фермента — Н+- АТФазы, перекачивающей протоны из цитоплазмы в лизосомы. Лизосомы выполняют функцию клеточного пищеварения, расщепляя отработавшие компоненты клетки или вещества, поглощенные в результате фагоцитоза и пиноцитоза.
Пероксисомы
 Рис.
10. Пероксисомы в клетках растений, ТЭМ.
С — хлоропласты, Р — пероксисомы, m —
митохондрия
Рис.
10. Пероксисомы в клетках растений, ТЭМ.
С — хлоропласты, Р — пероксисомы, m —
митохондрия
Пероксисомы среди одномембранных органелл стоят особняком, т. к. способны делиться самостоятельно, хотя не содержат ДНК. В них находятся ферменты, катализирующие некоторые окислительно-восстановительные реакции, в которых участвуют перекиси. Они также играют важную роль в обезвреживании многих токсичных веществ. Белки, которые находятся в пероксисомах, поступают туда из ЭПС и кодируются в геноме ядра. На электронных микрофотографиях пероксисом часто можно видеть в них кристаллы ферментов (рис. 10).
растительная и животная клетка

Рис. 11. Сравнение животной и растительной клеток
В растительных клетках существуют некоторые дополнительные органеллы (рис. 11). Это пластиды и крупная центральная вакуоль. В зрелых клетках растений центральная вакуоль занимает значительную часть клетки. Она поддерживает определенное осмотическое давление, служит местом запасания некоторых веществ, например органических кислот, и вывода ненужных веществ, например некоторых ионов, таких как кальций. Пластиды, как и митохондрии, представляют собой двумембранные структуры, имеющие собственные ДНК и рибосомы. Различают зеленые пластиды — хлоропласты, выполняющие функцию фотосинтеза, окрашенные в цвета от красного до желтого хромопласты и бесцветные пластиды — лейкопласты, в которых осуществляется синтез и запасание крахмала.
Транспорт веществ через мембрану. Эндоцитоз и экзоцитоз
В клеточные мембраны встроены различные специфические рецепторы — белки или гликопротеины, распознающие определенные молекулы (лиганды). В многоклеточном организме каждый тип клеток имеет свой уникальный набор поверхностных рецепторов.
Разветвленные цепи гликопротеинов, выступающие из клеточной мембраны, участвуют в распознавании факторов внешней среды, а также во взаимном узнавании клеток (например, взаимное узнавание яйцеклетки и сперматозоида при оплодотворении или правильная ориентация клеток в процессе образования тканей). С распознаванием связана и регуляция транспорта молекул и ионов через мембрану, а также иммунологический ответ, в котором гликопротеины и белки мембран играют роль антигенов.
Мембранные белки участвуют в транспорте определенных молекул внутрь клетки или из нее, осуществляют структурную связь цитоскелета с клеточными мембранами, служат в качестве рецепторов для получения и преобразования химических сигналов из окружающей среды.
ОСНОВНЫЕ СВОЙСТВА КЛЕТОЧНОЙ МЕМБРАНЫ
Обладает избирательной проницаемостью, которая изменяется при различных состояниях клетки
Имеет каналы, через которые проникают ионы:
потенциалзависимые каналы — открываются при изменении разности потенциалов;
потенциалнезависимые (лигандозависимые, гормонрегулируемые) — открываются при взаимодействии рецепторов с веществами.
Важнейшим свойством мембраны является избирательная проницаемость. Это значит, что молекулы и ионы проходят через нее с различной скоростью. Это свойство определяет плазматическую мембрану как осмотический барьер. Максимальной проникающей способностью обладают неполярные простые вещества, например, азот, кислород; значительно медленнее проходят сквозь мембрану полярные частицы, такие, как вода, и практически не проходят заряженные частицы — ионы.
МЕХАНИЗМЫ МЕМБРАННОГО ТРАНСПОРТА
Транспорт веществ через мембрану клетки осуществляется диффузией через липидный бислой или посредством двух классов мембранных белков — переносчиков или каналов.
Проницаемость плазматической мембраны
Если бы в плазматической мембране не присутствовали белки, легкость, с которой молекулы проходят сквозь фосфолипидный бислой по градиенту концентрации, зависела бы от размера молекулы, растворимости ее в жире и электрического заряда.
Малые жирорастворимые (неполярные) молекулы диффундируют быстро. Примеры: O2, N2. Крупные жирорастворимые молекулы, например, стероидные и тиреоидные гормоны, проходят через бислой с меньшей, но заметной скоростью.
Жиронерастворимые (полярные) молекулы способны проникать через бислой при условии малого размера и отсутствия полных зарядов. Примеры: H2O, CO2, мочевина. Большая проницаемость мембраны для воды обусловлена также наличием белковых каналов — аквапоринов. Также способны проходить через липидный бислой небольшие полярные молекулы этанола и глицерина.
Заряженные молекулы (ионы), даже при условии небольшого размера (Na+, K+, Cl–) практически не проникают через липидный бислой при отсутствии специальных транспортных механизмов.
Транспорт ионов и больших полярных молекул обеспечивается специальными трансмембранными белками. Так переносятся большие незаряженные полярные молекулы типа глюкозы и сахарозы.
Белки-переносчики передают вещества физическим движением одной части белковой молекулы относительно другой. Транспорт с помощью переносчиков может быть пассивным или активным (требует источник энергии).
Каналы транспортируют вещества по их электрохимическому градиенту. Такой транспорт не требует прямого расхода энергии и поэтому называется пассивным транспортом.

Пассивный транспорт
Пассивный транспорт может происходить непосредственно через фосфолипидный слой, через белки- переносчики или через белковые каналы.
Движущая сила может обеспечиваться:
разностью концентрации транспортируемого вещества (диффузия) или осмотического давления (осмос) на разных сторонах мембраны.
транспорт воды обеспечивается разностью осмотического давления с помощью белков-аквапоринов.
разностью электрического потенциала на мембране (если транспортируемое вещество несет заряд);
простая диффузия
Простая диффузия — перенос веществ через мембрану по градиенту концентрации (из области высокой концентрации в область низкой концентрации) без затрат энергии.
происходит по электрохимическому градиенту;
скорость линейно зависит от градиента концентрации вещества;
не насыщаемый процесс, то есть может ускоряться неограниченно;
не расходуется энергия.
Путём простой диффузии в клетку проникают гидрофобные вещества (кислород, азот, бензол) и полярные маленькие молекулы (вода, углекислый газ, мочевина). Не проникают полярные относительно крупные молекулы (аминокислоты, моносахариды), заряженные частицы (ионы) и макромолекулы (ДНК, белки).
осмос
Диффузия воды через мембрану называется осмосом (см. тему "Осмос и осморегуляция").
Осмос — односторонняя диффузия растворителя (воды) через полупроницаемую мембрану в более концентрированный раствор.
Из-за того, что более концентрированный раствор содержит меньшую концентрацию молекул растворителя, в него путем диффузии просачивается растворитель из менее концентрированного раствора и разбавляет его до тех пор, пока концентрация не станет равной по обе стороны мембраны.
В случае, когда мембрана плохо проницаема, либо непроницаема для данного вещества, она подвергается действию осмотических сил. При более низкой концентрации вещества в клетке вода из нее выходит, и клетка сжимается, при более высокой концентрации — впускает внутрь воду, и клетка увеличивается в объеме.
Облегченная диффузия
Облегченная диффузия — процесс трансмембранного переноса веществ по градиенту концентрации с участием мембранных белков-переносчиков без затраты энергии.
Каналы — белковые молекулы в мембране, создающие гидрофильный проход (облегченная диффузия). Характеристика каналов:
селективны (специфичны);
изменяют свое состояние;
чувствительны к лигандам и заряду мембраны.


С помощью облегченной диффузии в клетку попадают органические вещества с высокой полярностью.

Особенности облегченной диффузии
быстрее, чем простая диффузия;
происходит по электрохимическому градиенту;
с помощью белка-переносчика или белков трансмембранных каналов;
ограниченный по скорости и насыщаемый процесс;
можно блокировать, связав переносчик — чувствительна к ингибиторам;
конкуренция переносимых веществ за переносчик;
не расходуется энергия.
Активный транспорт
Активный транспорт — процесс трансмембранного переноса веществ против их градиента концентрации с затратами энергии.
Активный транспорт всегда происходит посредством белков-носителей, называемых транспортерами. Деятельность белкового насоса:
зависит от источника метаболической энергии:
первичные транспортеры требуют прямого использования АТФ, например, транспортные механизмы для Na+, K+, Са2+;
 Структура Na+/K+-АТФазы.
Структура Na+/K+-АТФазы.
вторичные транспортеры — белки, чьи транспортные функции требуют одновременного перемещения иона (обычно Na+) по градиенту концентрации, который поддерживается первичными транспортерами (например, транспортные механизмы для глюкозы и аминокислот);
2. вещества транспортируются против их электрохимического градиента
3. происходит только в одном направлении через плазматическую мембрану.
Унипорт — перенос через мембрану отдельного растворенного вещества.
Более сложная функция переносчиков-транспортировать два растворенных вещества совместно, так что перенос одного зависит от одновременного переноса другого в том же самом направлении (симпорт) или в противоположном направлении (антипорт).

Основные схемы совместного транспорта ионов
Энергия для активного транспорта выделяется при гидролизе АТФ:
АТФ = АДФ + Ф + энергия
С помощью активного транспорта в клетку проникают ионы (К+, Na+, Са2+, Mg2+) и мономеры (простые сахара, аминокислоты, нуклеотиды).
Осуществляется специальными белками-переносчиками, образующими так называемые ионные насосы, или помпы. Наиболее изученным является натрий-калиевый насос в клетках животных, активно выкачивающих ионы Na+ наружу, поглощая при этом ионы К+. Благодаря этому в клетке поддерживается большая концентрация К+ и меньшая Na+ по сравнению с окружающей средой. На этот процесс затрачивается энергия АТФ.
В результате активного транспорта с помощью мембранного насоса в клетке происходит также регуляция концентрации Mg2+ и Са2+.
ионные каналы
Ионный канал образуется одной или несколькими белковыми субъединицами, окружающими центральную пору.
Транспорт через ионные каналы всегда пассивен, и его максимальная скорость приблизительно в 1 000 раз больше, чем у белков-переносчиков.
Ионные каналы высоко селективны для определенных ионов
Ионные каналы изменяют свою активность (открываются в ответ на определенный стимул (раздражитель), который может быть электрическим, химическим или механическим).
Электрочувствительные каналы, осуществляя перенос Na+, К+ или Са2+, имеют большое значение для возбудимых клеток(например, нервных и мышечных).
Лигандчувствительные каналы распространены во всех органах и тканях и участвуют в восприятии клетками химических раздражителей.

1. 2.
Физическая (1) и химическая (2) активация ионных каналов.
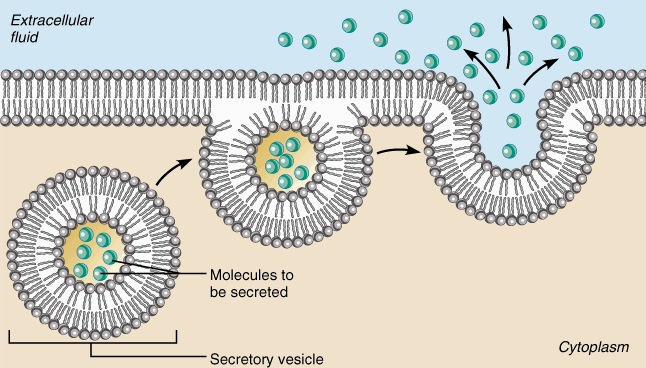
эндоцитоз
Транспорт макромолекул, их комплексов и крупных частиц внутрь клетки происходит совершенно иным путем — посредством эндоцитоза. Выведение тех же грузов из клетки называется экзоцитозом.
При эндоцитозе (эндо — внутрь) определенный участок плазмалеммы захватывает и как бы обволакивает внеклеточный материал, заключая его в мембранную вакуоль, возникшую вследствие впя-чивания мембраны. В дальнейшем такая вакуоль соединяется с лизосомой, ферменты которой расщепляют макромолекулы до мономеров.

экзоцитоз
Экзоцитоз (экзо — наружу) — процесс, обратный эндоцитозу. Благодаря ему клетка выводит внутриклеточные продукты или непереваренные остатки, заключенные в вакуоли или пузырьки. Пузырек подходит к цитоплазматической мембране, сливается с ней, а его содержимое выделяется в окружающую среду. Так выводятся пищеварительные ферменты, гормоны, гемицеллюлоза и др.
Таким образом, биологические мембраны как основные структурные элементы клетки служат не просто физическими границами, а представляют собой динамичные функциональные поверхности. На мембранах органелл осуществляются многочисленные биохимические процессы, такие как активное поглощение веществ, преобразование энергии, синтез АТФ и др.
Функции биологических мембран
Отграничивают содержимое клетки от внешней среды и содержимое органелл от цитоплазмы.
Обеспечивают транспорт веществ в клетку и из нее, из цитоплазмы в органеллы и наоборот.
Содержат рецепторы, необходимые для получения и преобразования сигналов из окружающей среды, узнавания веществ и других клеток и т. д.
Могут содержать ферменты — катализаторы реакций (обеспечение примембранных химических процессов).
Участвуют в преобразовании энергии (например, необходимы для электронного транспорта и синтеза АТФ в митохондриях и хлоропластах).
Строение клеток эукариот. Двумембранные органеллы
Двумя мембранами окружены особые органеллы эукариот — митохондрии и пластиды.
Митохондрии осуществляют кислородный этап дыхания, играя роль «энергетических станций» клетки. Пластиды бывают трех типов: хлоропласты, хромопласты и лейкопласты. Хлоропласты содержат зеленый пигмент хлорофилл и осуществляют фотосинтез. Хромопласты содержат пигменты из группы каротиноидов, которые придают им окраску от красного до желтого; их наличие обеспечивает окраску плодов и других органов растений. Лейкопласты — бесцветные пластиды, накапливающие зерна крахмала.
Двумембранные органеллы называют также полуавтономными, подчеркивая большую степень их самостоятельности в клетке. Прежде всего, это означает, что они могут делиться. Новые митохондрии и пластиды образуются только путем деления существующих. У этих органелл имеется собственный геном — кольцевая молекула ДНК, напоминающая ДНК бактерий. Она содержит гены, кодирующие часть белков органеллы. Другая часть кодируется в ядре и поступает из цитоплазмы (поэтому митохондрии и пластиды не могут жить свободно, вне клетки). Также эти органеллы имеют свой собственный аппарат синтеза белка, то есть рибосомы. Эти рибосомы более мелкие, чем в цитоплазме, и также похожи на рибосомы прокариот.
Все эти факты указывают на то, что двумембранные органеллы — это и есть прокариоты, которые когда-то вступили в симбиоз с древней эукариотической клеткой, поселившись внутри нее. Эукариоты часто фагоцитируют те или иные организмы, но не переваривают их, и они остаются жить в вакуолях. Часто в таких случаях встречается симбиоз, например у кораллов с некоторыми типами водорослей. Подобным же образом в эукариотической клетке поселились прокариоты — предки митохондрий, аэробные (способные к кислородному дыханию) бактерии из группы альфа-протеобактерий, а у растений — еще и предки хлоропластов — цианобактерии (зеленые прокариоты-фотосинтетики).
Когда в атмосфере накопился кислород, использование его для дыхания стало давать конкурентное преимущество. Кислородное окисление дает огромный выигрыш в энергии по сравнению с бескислородным. Поэтому симбиоз с предками митохондрий оказался полезным. Митохондрии имеются в клетках большинства эукариот (кроме тех, что утратили их вторично при переходе в бескислородные экологические ниши).
Цианобактерии обладали прогрессивным типом фотосинтеза, что дало преимущество «приютившим» их организмам. Все растения имеют тот же механизм фотосинтеза, что и цианобактерии, так как именно за счет симбиоза с ними «научились» фотосинтезировать.
Внешняя мембрана двумембранных органелл сходна по составу с мембранами эукариот, внутренняя сходна с мембранами прокариот. Это согласуется с гипотезой о том, что внешняя мембрана органеллы — это бывшая мембрана пищеварительной вакуоли (фагосомы), где оказался прокариотический симбионт, а внутренняя — это его собственная мембрана.
Такой внутренний симбиоз называют эндосимбиозом, а эту теорию происхождения двумембранных органелл — теорией эндосимбиогенеза.

Рис. 1. Теория эндосимбиогенеза: мембрана эукариот обозначена синим цветом, а мембраны прокариот-симбионтов — красным и зеленым; черным обозначена ДНК, точками — рибосомы

Рис. 2. Митохондрия: мембрана эукариотного происхождения обозначена синим цветом, а мембрана прокариоты-симбионта — красным; черным обозначена ДНК, точками — рибосомы

Рис. 3. Хлоропласт: мембрана эукариотного происхождения обозначена синим цветом, а мембрана цианобактерии-симбионта — зеленым. У цианобактерии уже имелись внутренние впячивания, образовавшие мешочки — тилакоиды, на мембране которых происходит фотосинтез. Черным обозначена ДНК, точками — рибосомы
Митохондрии
Снаружи митохондрии покрыты внешней мембраной, сходной с другими мембранами клетки. Расположенная под ней внутренняя мембрана отличается по липидному и белковому составу, напоминая мембрану бактерий. В ней высоко содержание белков, именно в ней находятся ферменты, осуществляющие окисление веществ кислородом, и синтез за счет этого АТФ. Для увеличения площади внутренняя мембрана митохондрий образует многочисленные складки — кристы. Между двумя мембранами находится изолированное от цитоплазмы межмембранное пространство. Внутри митохондрии находится пространство, называемое матрикс, в котором осуществляется расщепление различных веществ: жирных кислот, аминокислот и продуктов распада углеводов. В митохондриях имеются собственные рибосомы и ДНК. Они ответственны за образование части белков митохондрий.

Рис. 4
Пластиды
Пластиды имеют наружную и внутреннюю мембраны и могут иметь внутренние мембранные мешочки (тилакоиды и ламеллы), которые представляют собой отпочковавшиеся внутрь впячивания внутренней мембраны. Все типы пластид образуются из пропластид, а также могут превращаться друг в друга.

Рис. 5. Взаимопревращения пластид
Система внутренних мембран хлоропласта включает в себя совокупность мембранных каналов, называемых ламеллами, и интенсивно окрашенные образования, называемые гранами. Граны представляют собой стопки плоских мембранных пузырьков — тилакоидов, в мембранах которых содержится хлорофилл и осуществляются светозависимые реакции фотосинтеза. Внутренняя полость хлоропласта называется строма. Она заполнена раствором ферментов, осуществляющих темновые (светонезависимые) реакции фотосинтеза.

Рис. 6
Строение клеток эукариот. Немембранные органеллы
В клетке эукариот присутствуют следующие основные немембранные органеллы:
рибосомы (молекулярные машины для синтеза белка);
цитоскелетные нити;
построенные на основе цитоскелетных нитей структуры:
клеточный центр;
органоиды движения: жгутики, реснички, миофибриллы (сократимые нити поперечно-полосатых мышечных волокон).
РИбосомы
Рибосомы представляют собой мелкие органеллы, состоящие из рибосомных РНК (рРНК) и рибосомных белков. Они видимы в электронный, но не световой микроскоп, т. к. имеют размеры порядка. Функция рибосом — биосинтез белка. Работающая рибосома включает в себя 2 субъединицы — большую и малую. Они соединяются при синтезе белка. До начала синтеза белка субъединицы должны быть диссоциированы, то есть плавать в цитоплазме отдельно друг от друга. Они могут быть и соединены, но начать синтез белка могут, только предварительно разъединившись.
 Рис.
1. Структура рибосомы археи Thermus
thermofilus
с атомарным разрешением. Малая субъединица
— желтая, большая — голубая. Видны цепи
рРНК и «облепляющие» их белки
Рис.
1. Структура рибосомы археи Thermus
thermofilus
с атомарным разрешением. Малая субъединица
— желтая, большая — голубая. Видны цепи
рРНК и «облепляющие» их белки
Рибосомы про- и эукариот различаются по форме и массе. Экспериментально это можно установить путем центрифугирования. Константа седиментации — величина, характеризующая скорость осаждения частиц (измеряется в S — единицах Сведберга). Как правило, у частиц большего размера константа седиментации больше, но она зависит и от формы частицы.
Рибосомы прокариот имеют константу седиментации 70S, состоят из малой — 30S-субъединицы и большой — 50S-субъединицы. Константа седиментации не аддитивна: 70S не равно 30S + 50S. Это объясняется тем, что при соединении субъединиц меняется общая площадь их поверхности и, соответственно, сопротивление раствора при осаждении. В малой субъединице рибосом прокариот 1 молекула рРНК, в большой — 2 (все молекулы рРНК в рибосоме разные и характеризуются особой структурой и массой).
Рибосомы эукариот имеют константу седиментации 80S, состоят из малой — 40S-субъединицы и большой — 60S-субъединицы. В малой субъединице рибосом эукариот 1 молекула рРНК, в большой — 3 (на одну больше, чем у прокариот).

Рис. 2
|
Прокариоты |
Эукариоты |
Малая субъединица |
30S, 1 рРНК |
40S, 1 рРНК |
Большая субъединица |
50S, 2 рРНК |
60S, 3 рРНК |
Полная рибосома |
70S |
80S |
Рибосомы в клетках эукариот могут быть свободными, то есть находиться в цитоплазме, или быть ассоциированными с внешней (цитоплазматической) стороной шероховатой ЭПС. Во втором случае они ассоциированы с ЭПС только в ходе синтеза белка, который должен встроиться в мембрану или пройти в полость ЭПС, и после окончания синтеза такого белка рибосома «отваливается» от ЭПС и снова уходит в цитоплазму.
цитоскелет
Эукариотическую клетку пронизывает система структур, называемая цитоскелетом. Он выполняет функции опоры, поддержания формы клетки, движения (как всей клетки, так и различных грузов внутри нее, в том числе пузырьков и органелл).
Цитоскелет эукариотической клетки включает белковые волокна 3 типов:
микротрубочки;
актиновые нити (тонкие филаменты);
промежуточные филаменты.
 Рис.
3. Три типа цитоскелетных нитей, слева
направо: актиновые филаменты
(микрофиламенты), промежуточные филаменты,
микротрубочки
Рис.
3. Три типа цитоскелетных нитей, слева
направо: актиновые филаменты
(микрофиламенты), промежуточные филаменты,
микротрубочки
микротрубочки
Самые толстые из цитоскелетных нитей — микротрубочки, их диаметр порядка 25 нм. Это полые трубки, которые построены из димеров белка тубулина (от лат. «тубула» — трубочка), уложенных по спирали. Тубулин для построения микротрубочки должен быть связан с ГТФ.

Рис. 4
Микротрубочка имеет плюс-конец, на котором преимущественно происходит ее рост, и минус-конец, где преимущественно происходит разборка. Чтобы микротрубочка не разбиралась с минус-конца, она должна быть прикреплена к центру организации микротрубочек, который блокирует минус-конец и не дает микротрубочке разбираться.
клеточный центр. центриоли
В клетке микротрубочки радиально (звездообразно) расходятся в стороны от клеточного центра, где находятся центры организации микротрубочек.

Рис. 5. Клеточный центр, электронная микроскопия. C, C' — две центриоли, МТ — микротрубочки
В клетках животных в клеточном центре находятся парные образования, называемые центриолями. Центриоли представляют собой полые цилиндры, расположенные перпендикулярно друг другу. Эти цилиндры построены из микротрубочек. В каждом из них 9*3 — 9 триплетов, то есть троек, микротрубочек. В клетках растений и высших грибов центриолей нет.

Рис. 6

Рис. 7
Начало сборки микротрубочек из тубулиновых димеров происходит в клеточном центре. Микротрубочки составляют основу жгутиков и ресничек. По ним осуществляется транспорт клеточных органелл.
Клеточный центр способен удваиваться — каждая из центриолей достраивает возле себя дочернюю. Два образовавшихся клеточных центра расходятся и становятся полюсами так называемого веретена деления, организуя микротрубочки, которые растаскивают хромосомы эукариот по двум дочерним клеткам.

Рис. 8. Удвоение центриолей
Центриоли также обязательно находятся в основании жгутиков и ресничек эукариот. Такие центриоли называются базальным телом жгутика или реснички.
моторные белки
Моторные белки — сложные молекулярные машины, благодаря которым движутся организмы, перемещаются пузырьки и другие «грузы» внутри клеток, происходят изменения формы клеток.
Моторные белки способны расщеплять АТФ или ГТФ и за счет выделяющейся при этом энергии «шагать» по цитоскелетным нитям — актину или микротрубочкам. Все они устроены похожим образом. По актиновым нитям (микрофиламентам) способны «шагать» миозины — неотъемлемый компонент мышечных волокон, а по микротрубочкам — динеины и кинезины. За счет работы динеина, «шагающего» по микротрубочкам, бьются жгутики и реснички.

Рис. 9
ЖГУТИКИ И РЕСНИЧКИ
У эукариот жгутики и реснички внутренние, то есть покрыты мембраной снаружи. Они построены из микротрубочек. В основе жгутика либо реснички — микротрубочковая структура 9*2 + 2 (9 дублетов — двоек — микротрубочек по периферии + 2 свободные микротрубочки в центре). Такая структура называется аксонемой.

Рис. 10

Рис. 11. Реснички эпителия бронхиол, электронная микроскопия
Жгутик способен волнообразно изгибаться, а ресничка — биться, как хлыст. Движение происходит за счет перемещения по микротрубочкам аксонемы моторного белка динеина. Он как бы стремится вытолкнуть микротрубочку, по которой идет, вверх, но так как длина жгутика ограничена, он при этом вынужден изгибаться. Попеременное изгибание в разные стороны достигается активацией работы динеина то на одной, то на другой стороне жгутика.

Рис. 12. Биение ресничек
микрофиламенты — актиновые нити
Второй вид — микрофиламенты, построенные из глобул белка актина, связанного с АТФ. Диаметр микрофиламентов — порядка 6 нм, это самые тонкие цитоскелетные нити.

Рис. 13
Актиновые нити не организованы радиально вокруг центра, как микротрубочки, а образуют трехмерную сеть, особенно плотную под мембраной клетки. Они необходимы для поддержания формы поверхности клетки (субмембранный кортекс). Сборка и разборка актинового цитоскелета лежит в основе амебоидного движения, ползания клетки по субстрату, циркулярного движения цитоплазмы у растений.
По актину способны перемещаться («ходить») моторные белки — миозины. Актиново-миозиновые сократимые комплексы обеспечивают деление клетки животных и некоторых простейших путем перетяжки, а также сокращение всех видов мышц (гладких и поперечно-полосатых).
промежуточные филаменты
Третий вид волокон – промежуточные филаменты. Они называются так потому, что имеют диаметр около 10 нм — промежуточный между актиновыми нитями и микротрубочками.
Белковый состав промежуточных филаментов тканеспецифичен. Например, к ним относятся белки кератины, характерные для эпителиев и входящие в состав роговых производных эпидермиса. Другие белки промежуточных филаментов — десмин, виментин, а также ламины — белки внутренней выстилки ядерной оболочки. Важно отметить, что все мономеры промежуточных филаментов — фибриллярные белки, то есть белки, молекула которых имеет вид волокна вытянутой структуры. Этим они отличаются от микротрубочек и микрофиламентов, мономеры которых — глобулярные (округлые) белки актин и тубулин.
Промежуточные филаменты стабильны (в отличие от динамичных микротрубочек и тонких филаментов, которые подвержены постоянной сборке-разборке) и в основном отвечают за поддержание формы клеток. Участие в движениях для них нехарактерно. В клетке все эти типы цитоскелета существуют параллельно и функционируют координированно.

Рис. 14. Цитоскелетные нити в одной и той же клетке — флуоресцентная микроскопия

Рис. 15. Электронная микрофотография цитоскелета
Клеточный цикл. Понятие о дифференцировке. Стволовые клетки
клеточный цикл
Деление клетки может происходить только после удвоения (репликации) ее ДНК, так как каждая из дочерних клеток должна получить свою копию генетического материала. У прокариот ДНК кольцевая и имеет, как правило, только одну точку начала репликации. Когда движущиеся в противоположные стороны репликативные вилки встречаются, это означает окончание репликации кольцевой ДНК.
У эукариот геном гораздо больше, чем у прокариот, и представлен совокупностью отдельных линейных хромосом. В каждой из них находится много точек начала репликации. Соответственно, очень важно наличие механизма, контролирующего, чтобы весь геном был отреплицирован один и только один раз. При этом клеточное деление не должно начинаться до завершения репликации. Репликация генома эукариот занимает довольно много времени (гораздо больше, чем деление). К делению и к репликации также требуется подготовка. Все эти ограничения привели к возникновению сложной системы контроля событий в жизни эукариотической клетки — клеточного цикла.
 Клеточный
цикл состоит из четырёх фаз (см. рис.).
После деления клетка вступает в фазу
G1, или
предсинтетическую фазу. В это время
осуществляется рост клетки образование
новых органелл, создаётся запас
питательных веществ. Затем наступает
S фаза, в течение которой синтезируется
ДНК. Надо отметить, что две дочерние
молекулы ДНК, образовавшиеся в результате
репликации, не расходятся, а остаются
связанными между собой специальными
белками. После завершения репликации
начинается постсинтетическая фаза G2,
в течение которой клетка готовится к
делению: синтезируются белки, необходимые
для процесса деления, накапливается
запас богатых энергией веществ. И только
тогда, когда все эти процессы завершены,
клетка вступает в стадию деления,
называемую митозом.
Клеточный
цикл состоит из четырёх фаз (см. рис.).
После деления клетка вступает в фазу
G1, или
предсинтетическую фазу. В это время
осуществляется рост клетки образование
новых органелл, создаётся запас
питательных веществ. Затем наступает
S фаза, в течение которой синтезируется
ДНК. Надо отметить, что две дочерние
молекулы ДНК, образовавшиеся в результате
репликации, не расходятся, а остаются
связанными между собой специальными
белками. После завершения репликации
начинается постсинтетическая фаза G2,
в течение которой клетка готовится к
делению: синтезируются белки, необходимые
для процесса деления, накапливается
запас богатых энергией веществ. И только
тогда, когда все эти процессы завершены,
клетка вступает в стадию деления,
называемую митозом.
Прохождение клеткой фаз клеточного цикла регулируется специальными белками — циклинами. Определенный набор циклинов характерен для каждой фазы. Они взаимодействуют с белками, осуществляющими те или иные эффекты в ходе данной фазы.
На границе некоторых фаз клеточного цикла, а именно, перед делением и перед репликацией, существуют критические точки — так называемые чекпойнты. В этот момент специальные системы контроля проверяют, завершена ли полностью предыдущая фаза (например, репликация) и не произошло ли в ДНК клетки накопления большого количества повреждений (они должны быть исправлены до того, как клетка начнет деление). Также имеется чекпойнт в метафазе митоза, когда все хромосомы должны обязательно расположиться в экваториальной плоскости клетки.
дифференцировка клеток
В многоклеточном организме все клетки приспособлены для выполнения определенных функций. Нервные клетки, клетки крови, кости, эпителия, хрусталика — все они совершенно разные. При этом геном всех этих клеток одинаков, за редкими исключениями (например, эритроциты человека не имеют ядра, а в иммунных клетках происходят генетические перестройки). В каждой клетке работают далеко не все гены, а только их определенный набор. В зависимости от того, какой набор генов включен в той или иной клетке (или, как говорят, экспрессируется в данной клетке), она принадлежит к тому или иному клеточному типу. Все клетки многоклеточного организма появляются в результате деления одной-единственной клетки — оплодотворенной яйцеклетки, или зиготы. По мере формирования многоклеточного организма происходит дифференцировка клеток (англ. to differ — различаться) — включение и выключение определенных генов и специализация клетки для выполнения тех или иных функций, приобретение ею черт того или иного клеточного типа. В некоторых случаях этот процесс обратим, и клетка может вернуться в менее дифференцированное состояние (дедифференцироваться), после чего возможно превращение в другой клеточный тип, но по мере дальнейшей дифференцировки она, как правило, становится необратимой. Выделяют терминально дифференцированные клетки — те, которые уже не могут дедифференцироваться, например, поперечно-полосатые мышечные волокна, большинство нейронов. Как правило, дифференцированные клетки не могут делиться (представьте, как смог бы поделиться нейрон со всеми его отростками и синапсами, или мышечное волокно с его сократимым скелетом). По мере дифференцировки способность клетки к делению угасает. То есть в организме делятся в основном клетки, еще не приобретшие специфических черт определенного клеточного типа. Исключение составляют, например, гепатоциты (клетки печени), способные делиться в дифференцированном состоянии. С этим связывают высокую способность к регенерации, характерную для печени. В основном же делятся в многоклеточном организме так называемые стволовые клетки — это недифференцированные предшественники одного или нескольких клеточных типов. При делении стволовой клетки образуется две дочерних клетки, одна из которых становится снова стволовой (за счет чего не уменьшается общее число стволовых клеток), а вторая вступает на путь дифференцировки и в конце концов превращается в терминально дифференцированную клетку того или иного типа. За исследования стволовых клеток Гёрдону и Яманаке в 2012 году присуждена Нобелевская премия. Их изучение крайне важно для регенеративной медицины, прежде всего, для выращивания искусственных органов. Ученые уже существенно продвинулись по этому пути и научились получать из стволовых клеток различные искусственные органы и ткани.
Клеточное деление. Митоз
Одним из положений клеточной теории является постулат, что все клетки образуются делением ранее существующих клеток. Механизмы деления клеток про- и эукариот различаются.
бинарное деление клеток прокариот
В случае прокариот деление осуществляется довольно просто. Клетка растет за счет поглощения веществ окружающей среды, при наличии достаточного количества веществ начинается репликация ДНК. Так же, как и родительская молекула ДНК, дочерние прикреплены к клеточной мембране, но в разных точках. Дальнейший рост клетки приводит к росту мембраны, что приводит к растаскиванию дочерних молекул ДНК, после чего клетка делится пополам простой перетяжкой.

Рис. 1
деление клеток эукариот
В случае эукариотической клетки, которая содержит множество разнообразных органелл, возникает проблема распределения между дочерними клетками различных клеточных структур. Компоненты цитоскелета и одномембранные структуры могут разбираться на фрагменты и собираться заново, поэтому нет проблемы передачи этих структур дочерним клеткам: они возникают заново. Митохондрии и пластиды содержат свой собственный геном, поэтому могут образовываться путем деления предсуществующих митохондрий и пластид. Но они делятся независимо от деления ядра клетки, и количество таких органелл в клетке достаточно велико (клетка печени содержит более 1000 митохондрий), поэтому при случайном распределении они попадают в обе дочерние клетки.
Проблема возникает при распределении наследственного вещества. Оно представлено у эукариот несколькими или многими линейными молекулами, каждая их которых образует хромосому. Каждая дочерняя клетка при делении должна получить одну полную копию генетического материала, представленного всеми хромосомами. Если какая-то из дочерних клеток после деления получит не весь набор хромосом или лишние хромосомы, то это означает, что в ней произошла геномная мутация — изменение генетического материала. Это может иметь весьма серьезные последствия для клетки, в том числе ее гибель или превращение в раковую клетку. Поэтому в ходе эволюции в клетках эукариот сформировался высокоточный механизм расхождения линейных хромосом к разным полюсам клетки при помощи микротрубочек — белковых нитей цитоскелета. Микротрубочки при делении клетки образуют своеобразную структуру, называемую веретеном деления. Полюса веретена деления представлены клеточными центрами, от которых расходятся микротрубочки. Клеточные центры играют роль центров организации микротрубочек. У животных и части одноклеточных в каждом из клеточных центров находится по две центриоли (это структуры из микротрубочек, см. тему «Немембранные органеллы»). Часть микротрубочек, отходящих от каждого из полюсов, встречается и контактирует посередине клетки. Они участвуют в расталкивании полюсов друг от друга, чтобы они вместе с отошедшими к ним хромосомами в результате деления попали в разные клетки. Часть микротрубочек присоединяется к первичным перетяжкам, или центромерам, хромосом. Каждая хромосома имеет в области центромеры специфический белковый комплекс для прикрепления микротрубочек — кинетохор.

Рис. 2: 1 — хромосомы; 2 — клеточный центр (2 центриоли); 3–5 — микротрубочки веретена

Рис. 3
Наличие веретена деления является отличительной особенностью деления эукариотических клеток по сравнению с прокариотами.
Центриоли удваиваются между делениями клетки. После деления в клетке есть 1 клеточный центр, содержащий 2 центриоли. Затем центриоли немного расходятся, и около каждой из них начинается формирование перпендикулярной дочерней центриоли. В результате перед делением в клетке оказывается 2 пары центриолей (т. е. всего 4) в виде двух клеточных центров, которые расходятся и становятся полюсами веретена деления.
Существует два типа деления клеток эукариот: митоз и мейоз. При митозе хромосомный набор (плоидность) клетки не меняется, обе дочерние клетки полностью генетически идентичны исходной. Это обычный способ деления клеток, например, при формировании тел многоклеточных животных и растений. В результате мейоза, который включает в себя 2 деления, из одной диплоидной клетки получается 4 гаплоидных, причем все они генетически отличаются друг от друга. Это может происходить при формировании гамет или спор. В некоторых книгах можно встретить упоминание третьего типа деления эукариот — амитоза, под которым подразумевается прямое деление ядра клетки без участия микротрубочек и веретена деления. В настоящее время все явления, которые описывались ранее как амитоз, считаются либо артефактами (то есть следствиями повреждения клеток при приготовлении препаратов), либо следствиями апоптоза (запрограммированной клеточной смерти) и других известных процессов.
В неделящейся клетке хромосомы не различимы, хотя они присутствуют в ядре. Так происходит потому, что плотность упаковки генетического материала в клетке может быть разной. Максимально плотными хромосомы становятся в начале деления. Поэтому при делении хромосомы в клетке различимы даже в световой микроскоп. После деления упаковка хромосом становится менее плотной, поэтому мы перестаем видеть их границы, хотя они сохраняют свою целостность, но находятся в состоянии «размотанного клубка». Однако их все-таки можно увидеть при помощи специфических методов визуализации хромосом. Совокупность ДНК и связанных с ней белков в неделящейся клетке, которая представляет собой деспирализованные хромосомы, называется хроматином. Почему так происходит? Дело в том, что в том состоянии, в котором хромосомы находятся при делении, генетический материал упакован настолько плотно, что он не может работать. Не может происходить транскрипция, репликация. Да это и не нужно. Главная задача при делении — быстро распределить хромосомы по дочерним клеткам. После деления хромосомы уже не могут оставаться в таком плотно упакованном состоянии, иначе клетка не сможет ими «пользоваться», и ее жизнедеятельность будет невозможна. Поэтому хромосомы «распаковываются», декомпактизуются. Можно сравнить эти процессы с упаковкой вещей при переезде и их распаковкой после переезда в новую квартиру. В начале деления происходит, таким образом, конденсация хроматина, или спирализация хромосом (это одно и то же), а в конце деления — деконденсация хроматина, или деспирализация хромосом.
фазы митоза
Митоз состоит из четырех фаз: профазы, метафазы, анафазы и телофазы.
Профаза является фазой подготовки к разделению хромосом (см. рис. 4). Происходит разборка ядрышка, оно исчезает. Хроматиновые нити укорачиваются и утолщаются, происходит процесс, называемый конденсацией хроматина. В результате образуются хромосомы, каждая из которых состоит из двух сестринских хроматид. Каждая из хроматид содержит всегда одну молекулу ДНК, при этом молекулы ДНК сестринских хроматид — это две точные копии, образовавшиеся в результате репликации ДНК (поэтому они и называются сестринскими). Сестринские хроматиды остаются соединенными только в области центромеры, образуя Х-образную структуру. Два клеточных центра расходятся к противоположным концам клетки, образуя полюса веретена деления. От каждого из них начинают расти микротрубочки. Встречаясь с хромосомами, микротрубочки прикрепляются к их центромерам, причем к каждой хромосоме прикрепляется две микротрубочки: одна от одного полюса, а вторая — от другого. Микротрубочки, которые не прикрепились к хромосомам, соединяются между собой в середине клетки. Образуется структура, называемая веретеном деления, а микротрубочки называются нитями веретена. Последним событием профазы является распад ядерной оболочки на фрагменты (пузырьки).

Рис. 4
Затем наступает метафаза (см. рис. 5). За счет изменения длины нитей веретена хромосомы перемещаются в среднюю часть клетки, образуя экватор деления. В это время они все находятся в одной плоскости, что позволяет провести исследование их набора, формы и размеров. Приводимые обычно фотографии хромосомных наборов делаются именно на этой стадии, образуя так называемую метафазную пластинку. Хромосомы находятся в экваториальной плоскости довольно длительное время. Как показано в последние годы, в это время происходит смена белков в центромерах хромосом, что позволяет в дальнейшем разделить их.

Рис. 5
За метафазой наступает анафаза (см. рис. 6). Центромеры сестринских хроматид разделяются, нити веретена укорачиваются, в результате дочерние хроматиды расходятся к противоположным полюсам. Такое движение продолжается до тех пор, пока хроматиды, ставшие самостоятельными хромосомами, не достигнут полюсов. В результате у каждого полюса деления образуется полный набор хромосом, характерный для данного вида.

Рис. 6
Митоз завершается телофазой, в которой восстанавливается исходная структура ядер (см. рис. 7). Вокруг каждого набора хромосом у полюсов деления формируется новая ядерная оболочка. После этого начинается удлинение и уменьшение толщины хромосом, называемое деконденсацией хроматина. В результате хромосомы превращаются в хроматиновые нити. Формируются новые ядрышки. На этом завершается митоз и начинается деление клетки — цитокинез.

Рис. 7
Он происходит по-разному у животных и растений. У животных в средней части образуется бороздка, опоясывающая клетку, — экваториальная борозда. В этой области перетяжки формируется сократимое кольцо из белков актина и миозина (подобно мышечным волокнам). Борозда постепенно углубляется, образуя перетяжку, которая в конце концов делит клетку на две равные части.
У растений жесткая клеточная стенка не позволяет образовать перетяжку. После расхождения хромосом и образования ядер к нитям веретена прикрепляются мембранные пузырьки, находящиеся в цитоплазме. Они перемещаются по нитям веретена на экватор клетки. Там происходит вскрытие пузырьков (см. рис. 8). Их содержимое застывает, образуя срединную пластинку (фрагмопласт), а мембраны пузырьков формируют с двух сторон от нее две новые клеточные мембраны. Эти мембраны затем синтезируют целлюлозные волокна, формирующие две новые клеточные стенки и склеенные срединной пластинкой.

Рис. 8
Результатом митоза является образование двух новых генетически идентичных клеток. У одноклеточных эукариот митоз является способом бесполого размножения. У многоклеточных митоз, приводящий к увеличению числа клеток, является основой роста. Кроме того, после митоза одна из образующихся клеток может начать превращение в специализированную клетку, выполняющую новую функцию. Такой процесс называется дифференцировкой клеток и лежит в основе процессов развития многоклеточных организмов.
Мейоз и его биологическое значение
Определение
Мейоз (редукционное деление клетки) — деление, в процессе которого из одной диплоидной (2n) клетки получаются 4 гаплоидные (n) клетки.
Так как у дочерних клеток происходит уменьшение (редукция) числа хромосом с 2n до n, такое деление названо редукционным.
Схема мейоза

Мейоз у животных наблюдается при формировании гамет (гаметогенезе). Мейоз у растений и грибов, как правило, происходит при образовании гаплоидных спор. У различных одноклеточных эукариот мейоз может наблюдаться на разных стадиях жизненного цикла. Для восстановления диплоидности в цикле всегда необходимо слияние гаплоидных клеток (оплодотворение).
Мейоз состоит из двух делений. Первое из них является собственно редукционным, то есть именно в ходе первого деления уменьшается плоидность клетки. Причиной этого служит расхождение гомологичных хромосом («материнской» и «отцовской») по двум разным дочерним клеткам. Второе деление аналогично митозу и называется эквационным (то есть «равным»). Плоидность в результате второго деления не меняется. В ходе этого деления, как и при митозе, расходятся сестринские хроматиды (копии ДНК). Между двумя делениями мейоза отсутствует репликация ДНК (так как «цель» мейоза — уменьшить плоидность клетки, увеличивать количество ДНК здесь незачем).
В профазе I деления мейоза происходит важнейший процесс, относящийся к генетической рекомбинации — кроссинговер, то есть обмен участками гомологичных хромосом. В результате этого процесса создаются новые комбинации генов в потомстве. Хромосомы как целое не передаются напрямую от бабушек и дедушек внукам, а «реконструируются» в каждом поколении в процессе кроссинговера.
В нижеследующей таблице приведено описание фаз мейоза в клетке, для которой n=3, 2n=6. В каждом наборе есть три хромосомы, различающиеся по размеру. Материнский и отцовский хромосомные наборы выделены синим и красным.
Фаза |
Процесс |
Описание |
Профаза I 2n4c |
|
конденсация (сверхспирализация) хромосом (видны в электронный микроскоп); конъюгация (соединение) гомологичных хромосом с образованием бивалентов; кроссинговер — обмен участками между гомологичными хромосомами; гомологичные хромосомы остаются соединенными между собой; ядерная оболочка растворяется; центриоли расходятся к полюсам |
Метафаза I 2n4c |
|
биваленты выстраиваются вдоль экватора клетки |
Анафаза I 2n4c |
|
микротрубочки веретена деления сокращаются, биваленты делятся; к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая |
Телофаза I n2c x 2 |
|
хромосомы деспирализуются («раскручиваются»); формируется ядерная оболочка |
Второе деление мейоза следует непосредственно за первым, без интерфазы: репликации (удвоения) ДНК не происходит. |
||
Профаза II n2c x 2 |
|
конденсация (сверхспирализация) хромосом; клеточный центр делится, центриоли расходятся к полюсам ядра; разрушается ядерная оболочка; образуется веретено деления |
Метафаза II n2c x 2 |
|
двухроматидные хромосомы располагаются в плоскости «экватора» (метафазная пластинка) |
Анафаза II 2nc x 2 |
|
центромеры делятся; однохроматидные хромосомы расходятся к полюсам |
Телофаза II nc x 4 |
|
хромосомы деспирализуются; формируется ядерная оболочка |








Протекание мейоза, как правило, нарушается в клетках гибридных организмов, т. к. в профазе I должно происходить попарное слияние (конъюгация) гомологичных хромосом, а у гибридов набор материнских генов не гомологичен отцовскому.
Данный механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и в конечном счете к нежизнеспособности гамет, а следовательно, к стерильности (бесплодию) гибридов.
В селекции для преодоления стерильности гибридов искусственно вызывают полиплоидность (кратное увеличение) хромосомных наборов. В этом случае каждая хромосома конъюгирует с соответствующей хромосомой своего набора.
Значение мейоза
Половые клетки родителей, образовавшиеся путем мейоза, обладают гаплоидным набором (n) хромосом. В зиготе при объединении двух таких наборов число хромосом становится диплоидным (2n). Формирование нового организма происходит путем митотических делений зиготы, и каждая его клетка содержит диплоидный (2n) набор хромосом. Каждая пара гомологичных хромосом содержит одну отцовскую и одну материнскую хромосому. Исходя из этого:
Мейоз является основой комбинативной изменчивости благодаря кроссинговеру (профаза I) и независимому расхождению гомологичных хромосом (анафаза I и II).
Благодаря уменьшению количества хромосом в гаметах в новых организмах поддерживается постоянный диплоидный (2n) набор хромосом.
профаза I деления мейоза
Профаза I деления мейоза своеобразна, включает в себя множество процессов и подразделяется на стадии:
Лептотена
Зиготена
Пахитена
Диплотена
Диакинез







Основные типы тканей животных
Ткань — сложившаяся в процессе развития совокупность клеток и межклеточного вещества, имеющих сходное строение, происхождение и функции.
Ткани животных, в отличие от тканей растений, содержат много межклеточного вещества.
Существуют 4 основных типа животных тканей: эпителиальная (покровная), мышечная, соединительная, нервная.
эпителиальная ткань (эпителий)
Клетки эпителия -- эпителиоциты — лежат на тонкой базальной мембране, они лишены кровеносных сосудов, их питание осуществляется за счет лежащей под базальной мембраной соединительной ткани.
В покровном эпителии много нервных окончаний, передающих в центральную нервную систему сигналы о различных раздражениях.
виды |
особенности |
функции |
нахождение |
покровный
|
клетки мелкие, постоянно делятся - высокая способность к регенерации; клетка соединены с помощью плотных контактов; очень мало межклеточного вещества клетки ориентированы в пространстве (есть базальная и апекальная часть) |
отделяет внутреннюю среду от внешней; защитная; всасывание и выделение продуктов обмена; регенерация |
кожные покровы; слизистые оболочки полостей, сосудов и внутренних органов; серозные оболочки |
железистый |
секреторные клетки — гландулоциты: экзокринные — выделяют свой секрет во внешнюю среду или просвет органа; эндокринные — выделяют свой секрет непосредственно в кровоток. |
секреторная |
в железах кожи, кишечнике, слюнных железах, железах внутренней секреции и др. |
Классификация по строению:
Однослойный эпителий: один слой клеток, прикрепленных к базальной мембране.
однорядный: клетки одинаковой формы, ядра всех клеток лежат на одном уровне;
многорядный: клетки разной формы, ядра клеток лежат на разных уровнях.
Эндотелий и мезотелий
реснитчатый (мерцательный) эпителий
Многослойный эпителий: несколько слоев клеток, к базальной мембране прикреплен только самый глубокий слой.
ороговевающий: образует наружный слой кожи (эпидермис)
неороговевающий
переходный (уроэпителий): в органах, которые меняют форму — мочевой пузырь, аллантоис и др. При изменении объёма органа толщина и строение эпителия также изменяется. Эпителий способен выделять секрет, защищающий его клетки от воздействия мочи.
многослойный плоский неороговевающий эпителий
Развивается из эктодермы.
Выстилает роговицу, ротовую полость, преддверие анального отверстия и влагалище.
Клетки располагаются в несколько слоёв:
Слой стволовых клеток — на базальной мембране. Они делятся и превращаются в шиповатые клетки.
Слой шиповатых клеток (полигональной формы с выростами и шипами). Они постепенно уплощаются.
Поверхностный слой плоских клеток, которые с поверхности отторгаются во внешнюю среду.
кожный эпидермис
Многослойный плоский ороговевающий эпителий — эпидермис кожных покровов.
В толстой коже (ладонные поверхности), которая постоянно испытывает нагрузку, эпидермис содержит 5 слоёв:
1 — базальный слой — содержит стволовые клетки, дифференцированные цилиндрические и пигментные клетки (меланоциты).
2 — шиповатый слой — клетки неправильной формы с многочисленными выростами; содержат тонофибриллы — нитчатые образования, придающее коже механическую прочность;
3 — зернистый слой — клетки ромбовидной формы; в них начинается процесс ороговения;
4 — блестящий слой — клетки становятся плоскими, они постепенно утрачивают внутриклеточную структуру;
5 — роговой слой — содержит роговые чешуйки, которые полностью утратили строение клеток, содержат белок кератин.
В тонкой коже, которая не испытывает нагрузки, отсутствует блестящий слой.

По форме эпителиоциты бывают плоские, кубические, призматические, цилиндрические и т.п.

Строение покровного эпителия
А - однослойный плоский эпителий;
Б - однослойный кубический эпителий;
В - однослойный столбчатый эпителий;
Г - реснитчатый эпителий;
Д - переходный эпителий;
Е - неороговевающий многослойный плоский эпителий.
мышечная ткань
Виды мышечной ткани:

Гладкая мышечная ткань
Состоит из одноядерных клеток — миоцитов веретеновидной формы.
Свойства: сокращается ритмично, медленно и на протяжении длительного времени, способна при этом развивать большую силу без значительных затрат энергии и без утомления. Является непроизвольной (то есть ее деятельность не управляется по воле человека).
Входит в состав стенок внутренних органов: кровеносных и лимфатических сосудов, мочевыводящих путей, пищеварительного тракта (сокращение стенок желудка и кишечника).
Эти клетки имеют тонкие миофибриллы, которые тянутся от одного конца клетки к другому. Гладкие мышечные клетки объединяются в пучки, состоящие из 10 — 12 клеток. Это объединение возникает благодаря особенностям иннервации гладкой мускулатуры и облегчает прохождение нервного импульса на всю группу гладких мышечных клеток.
У низших многоклеточных животных из гладкой мышечной ткани состоят все мышцы. У позвоночных животных гладкая мышечная ткань входит в состав внутренних органов (кроме сердца).

Поперечно-полосатая скелетная мышечная ткань
Состоит из длинных нитевидных многоядерных миоцитов.
Свойства: высокая скорость сокращения и расслабления; характеризуется произвольным сокращением (сокращение в ответ на импульсы, идущие из коры больших полушарий). Скорость сокращения этой ткани в 10 — 25 раз выше, чем в гладкой мышечной ткани.
Входит в состав скелетных мышц, а также стенки глотки, верхней части пищевода, образует язык, глазодвигательные мышцы.
Мышечное волокно поперечно-полосатой ткани покрыто оболочкой — сарколеммой. Под оболочкой находится цитоплазма с большим количеством ядер, расположенных по периферии цитоплазмы, и сократительными нитями — миофибриллами. Миофибриллы состоят из белков: актина (тонкие нити) и миозина (толстые нити).
При сокращении мышечного волокна происходит возбуждение сократимых белков, тонкие протофибриллы скользят по толстым. Актин реагирует с миозином, и возникает единая актомиозиновая система.

Поперечно-полосатая сердечная мышечная ткань
Состоит из многоядерных кардиомиоцитов, имеющих поперечную исчерченность цитоплазмы. Кардиомиоциты разветвлены и образуют между собой соединения — вставочные диски, в которых объединяется их цитоплазма.
Свойство: автоматия — способность ритмично сокращаться и расслабляться под действием возбуждения, возникающего в самих клетках. Эта ткань является непроизвольной.
Образует миокард сердца.

соединительная ткань
Выполняет вспомогательная роль во всех органах.
Составляя 60-90 % от их массы.
Функция: опорная, защитная, трофическая, терморегуляция.
К соединительной ткани относят костную, хрящевую, жировую, кровь и лимфу. Поэтому соединительная ткань — единственная ткань, которая присутствует в организме в 4-х видах — волокнистом (связки), твёрдом (кости), гелеобразном (хрящи) и жидком (кровь и лимфа).
Общими свойствами всех соединительных тканей является происхождение из мезенхимы, а также выполнение опорных функций и структурное сходство.

1 2 3 4 5
1 — рыхлая соединительная ткань, 2 — плотная соединительная ткань, 3 — хрящ, 4 — кость, 5 — кровь.
Состав соединительной ткани:
межклеточное вещество;
клетки различного типа (фибробласты, хондробласты, остеобласты, тучные клетки, макрофаги);
волокнистые структуры. Соединительная ткань:
собственно соединительная ткань рыхлую волокнистую соединительную ткань: во всех органах: рыхлая сеть волокон и клеток; плотную неоформленную соединительную ткань: неправильно расположенные пучки волокон; плотную оформленную соединительную ткань: параллельные пучки волокон (сухожилия, связки);
скелетная ткань: костная, хрящевая, цемент и дентин зуба.
 Костная
ткань.
Костная
ткань.
соединительная ткань со специфическими свойствами: жировая, слизистая, пигментная, ретикулярная.

Нервная ткань
Состоит из нейронов.
Нейрон — нервная клетка, структурно-функциональные единицы нервной системы.

В состав нейрона входят:
дендриты — отростки, воспринимающие раздражения
аксон — отросток, передающий нервные сигналы от тела другим клеткам.
Дендритов у нейрона может быть много, аксон только один.
Функция: осуществляет связь организма с окружающей средой; обеспечивает взаимодействие тканей, органов и систем органов организма.
Общий обзор организма человека (органы и системы органов)
Орган — это часть тела, имеющая определенное строение и выполняющая специфические функции.
Орган состоит из нескольких видов тканей, часто одной — основной и вспомогательных.
Органы, входящие в состав тела человека, можно разделить на трубчато-полые и паренхиматозные.
трубчато-полые органы: органы сердечно-сосудистой системы, пищеварительного тракта, воздухоносных и мочевыводящих путей, большая часть органов мужского и женского половых аппаратов (кроме желез).
паренхиматозные органы: кроветворные органы, железы внутренней и внешней секреции, легкие, почки, органы нервной системы.
Стенка трубчато-полых органов имеет обычно три оболочки:
слизистая оболочка: разделяет внешнюю и внутреннюю среду (в артериях, венах и лимфатических сосудах — интима; в сердце — эндокард);
мышечная — средняя оболочка;
соединительнотканная — наружная оболочка. Стенки некоторых полостных внутренних органов покрывает серозная соединительнотканная оболочка. Благодаря своей гладкости и влажности серозная оболочка уменьшает трение между органами и окружающими их частями при движении.
Паренхиматозные органы составляют 2 основных структурных компонента:
паренхима — ткань, выполняющая специфические функции;
строма — обычно соединительная ткань, выполняющая опорную и трофическую функцию.
Система органов — несколько органов, имеющих общее происхождение и выполняющих определенную функцию.
Например, к пищеварительной системе относятся трубчато-полые органы пищеварительного тракта (пищевод, желудок, тонкий и толстый кишечник), а также паренхиматозные органы (слюнные железы, печень, поджелудочная железа).
функциональные системы
Некоторые авторы (П.К. Анохин) выделяют также функциональные системы организма.
Функциональная система организма — рабочие органы и связанная с ними часть нервной системы, обеспечивающие выполнение определенной функции.
В образовании функциональных систем могут участвовать органы, принадлежащие к разным анатомических систем. Например, в функциональную дыхательную систему входят органы дыхательной системы (легкие и воздухоносные пути), нервные окончания, афферентные и эфферентные нервные волокна, дыхательный центр в продолговатом мозге, дыхательные мышцы, ребра и соответствующие части сердечно-сосудистой системы.
Аппарат — группа органов, выполняющих общую функцию, но имеющих разное происхождение. Например, к опорно-двигательному аппарату относятся костная, мышечная и соматическая нервная система.
система / аппарат |
органы |
функции |
опорно-двигательный аппарат |
кости скелета и мышцы |
опора, движение, защита |
пищеварительная система |
кишечная трубка (ротовая полость, пищевод, желудок, тонкий и толстый кишечник) и пищеварительные железы |
измельчение, продвижение, химическая обработка пищи, всасывание питательных веществ, удаление непереваренных остатков |
дыхательная система |
воздухоносные пути (носовая и ротовая полость, трахея, бронхи) и альвеолярные легкие |
поступление в организм кислорода,выведение из организма углекислого газа и газообразных продуктов метаболизма |
мочевыделительная система |
почки, мочеточники, мочевой пузырь, мочеиспускательный канал |
выведение продуктов обмена веществ, излишка воды, токсинов, солей, регуляция артериального давления крови |
нервная система |
головной и спинной мозг, нервы, нервные узлы, рецепторы |
связь с внешней средой, координация работы органов |
половая система |
половые железы и выводные протоки |
размножение |
эндокринная система |
железы внутренней секреции |
регуляция работы органов, обмен веществ |
сердечно-сосудистая система |
сердце и сосуды |
питание, транспорт, защита, регуляция |
покровная система |
кожные покровы и кожные (экзокринные) железы |
защита от механический повреждений, УФ лучей, проникновения чужеродных тел; выведение продуктов метаболизма; терморегуляция |
иммунная система |
органы иммунной защиты (костный мозг, тимус, лимфатические узлы, селезенка) |
защита от чужеродных тел, инфекционных агентов (бактерий, вирусов, простейших); уничтожение измененных клеток, опухолей и т.п. |
Системы органов формируют целостный организм человека.
Нервная система. Общие сведения
Нервная система — иерархически организованная нервная ткань, пронизывающая весь организм и связывающая его в единое целое.
Нервная ткань состоит из нейронов и глии.
нейрон
Определение
Нейрон (нервная клетка) — основной структурный и функциональный элемент нервной системы.
У человека насчитывается более 100 млрд нейронов.
Взаимодействие между нейронами представляет собой передачу нервных сигналов (нервного возбуждения).
Свойства нервных клеток: возбудимость и проводимость.
Строение нейрона

Рис. 1. Нейрон
Нейрон состоит из тела (сомы) и отростков.
Тело нейрона содержит ядро (с большим количеством ядерных пор) и органеллы. Органеллы в нервной клетке те же, что и в других клетках.
Нейрон имеет развитый цитоскелет, проникающий в его отростки. Цитоскелет состоит из микрофиламентов и микротрубочек. Его функция: поддержание формы клетки, транспорт органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов — молекул — передатчиков нервных импульсов).
Из специфических органелл присутствует тигроид (тельца Ниссля) и нейрофибриллы.
Тигроид состоит из сильно развитой шероховатой ЭПС с активными рибосомами и аппарата Гольджи; его функция — синтез специфических белков. Выглядит эта структура как «мелкая зернистость и полосатость» в теле и дендритах нейрона (отсюда и название). Длительное голодание или стресс приводит к разрушению тигроида и прекращению синтеза специфических белков.
Рис. 2. Внутреннее строение нейрона
Нейрофибриллы (нейрофиламенты) состоят из микротрубочек и являются основным структурным компонентом цитоскелета. Их функция — аксональный транспорт (перемещение веществ по аксону).
Аксональный транспорт
Помимо своей специфической функции в качестве проводника нервных импульсов аксон является каналом для транспорта веществ.
Аксональный (аксонный) транспорт — это перемещение веществ по аксону. Белки, синтезированные в теле клетки, нейромедиаторы и низкомолекулярные соединения перемещаются по аксону вместе с клеточными органеллами, в частности митохондриями. Для большинства веществ и органелл обнаружен также транспорт в обратном направлении. Вирусы и токсины могут проникать в аксон на его периферии и перемещаться по нему. Аксональный транспорт — активный процесс — зависит от энергии АТФ. При снижении уровня АТФ вдвое аксональный транспорт блокируется.
Различают антероградный (от тела нейрона) и ретроградный (к телу нейрона) аксонный транспорт.
Выделяют два вида отростков: короткие ветвящиеся дендриты и один длинный не ветвящийся аксон.
Дендриты ветвятся дихотомически (надвое), аксоны же дают коллатерали (боковые ответвления). В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки. У большинства аксонов миелиновая оболочка имеется.
Миелиновая оболочка
Миелиновая оболочка — электроизолирующая оболочка, покрывающая аксоны многих нейронов. Миелиновая оболочка формируется из плоского выроста тела глиальной клетки, многократно оборачивающего аксон подобно изоляционной ленте.
В периферической нервной системе миелиновую оболочку аксонов образуют шванновские клетки (несколько шванновских клеток на один аксон). В ЦНС один олигодендроцит образует миелиновую оболочку нескольким нервным клеткам.

Рис. 3. Образование миелиновой оболочки в ЦНС
Цитоплазма шванновской клетки вытесняется из пространства между спиральными витками и остается только на внутренней и наружной поверхностях миелиновой оболочки, в результате чего миелиновая оболочка представляет собой, по сути, множество слоев клеточной мембраны.

Рис. 4. Миелиновая оболочка (микрофотография)
Химический состав миелина: 70–75 % липиды, 25–30 % белки. Такое высокое содержание липидов отличает миелин от других биологических мембран.
Миелин прерывается только в области перехватов Ранвье, которые встречаются через правильные промежутки длиной примерно 1 мм (расстояние между перехватами Ранвье прямо пропорционально толщине аксона). В связи с тем что ионные токи не могут проходить сквозь миелин, вход и выход ионов осуществляется лишь в области перехватов. Это ведет к увеличению скорости проведения нервного импульса. Таким образом, по миелинизированным волокнам импульс проводится приблизительно в 5–10 раз быстрее, чем по безмиелиновым.
Благодаря наличию миелиновой оболочки и совершенству метаболизма на всем протяжении мембраны в покое поддерживается одинаковый заряд, который быстро восстанавливается после прохождения возбуждения.
Цвет миелинизированных нейронов белый, отсюда название «белого вещества» мозга.
Безмиелиновые волокна изолированы по другой схеме. Несколько аксонов частично погружены в изолирующую шванновскую клетку, которая не смыкается вокруг них до конца. Возбуждение постепенно охватывает соседние участки мембраны и так распространяется до конца аксона с постепенным ослаблением (т. к. происходят большие затраты энергии).
Место нейрона, от которого начинается аксон, называется аксонным холмиком. Здесь генерируется потенциал действия — специфический электрический ответ возбудившейся нервной клетки. Аксон, выходя из сомы клетки, постепенно утончается и может давать ответвления — коллатерали.
Функция аксона — передача нервного импульса к аксонным терминалиям. В месте отхождения коллатерали импульс «дублируется» и распространяется как по основному ходу — аксону, так и по коллатералям. В конце аксона имеются синаптичекие окончания — аксонные терминалии.
В цитоплазме аксона отсутствует ЭПС и аппарат Гольджи. Нейрофиламенты и микротрубочки располагаются вдоль аксона и обеспечивают транспорт белков и других веществ.
Серое вещество мозга состоит из тел нейронов и дендритов. Белое вещество мозга состоит из аксонов.
функционирование нейрона
В нейроне нервные импульсы по дендритам проходят к соме клетки.
В аксонном холмике происходит генерация потенциала действия (нервный импульс).
Нервный импульс по аксону достигает аксонных терминалий, а с них переходит сразу на несколько нейронов или рабочих органов.
Благодаря отросткам нейроны контактируют друг с другом и образуют нейронные сети и круги, по которым циркулируют нервные импульсы. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Синапс
Определение
Синапс — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой (клеткой рабочего органа).
Термин был введен в 1897 г. английским физиологом Чарльзом Шеррингтоном.
Служит для передачи нервного импульса между двумя клетками.
Синапсы могут возникнуть между аксоном и телом нервной клетки, аксоном и дендритом, аксоном и аксоном.

Рис. 5. Нервные синапсы
Синапсы:
возбуждающие: усиливают нервный импульс;
тормозные: ослабляют нервный импульс.
Вдоль нервного волокна нервные импульсы распространяются в виде волн электрических потенциалов.

Рис. 6. Мембранный потенциал
В синапсе происходит смена механизма распространения. Когда нервный импульс достигает пресинаптического окончания, в синаптическую щель выделяется активное химическое вещество — нейромедиатор (нейротрансмиттер). Нейромедиатор проходит через синаптическую щель и меняет проницаемость постсинаптической мембраны, в результате чего на ней возникает потенциал, вновь генерирующий распространяющийся импульс. Так действует химический синапс.

Рис. 7. Химический синапс
Встречается также электрический синапс, когда нейрон возбуждается электрически.
виды нейронов
Нейроны различаются по форме, числу отростков и функциям.
По количеству отростков нейроны бывают (см. рис. 8):
1 — униполярные нейроны (нет дендритов, только аксон);
2 — биполярные нейроны (аксон и один дендрит);
3 — псевдоуниполярные нейроны (один аксон имеет Т-образную форму);
4 — мультиполярные нейроны (один аксон и много дендритов).

Рис. 8. Виды нейронов
Вид нейрона |
Местоположение и путь |
Функция |
Чувствительные нейроны |
от рецептора к ЦНС |
воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг |
Эффекторные нейроны (двигательные, секреторные) |
от ЦНС к исполнительному органу |
вырабатывают и посылают команды к рабочим органам |
Вставочные нейроны (интернейроны) |
в ЦНС |
осуществляют связь: между чувствительными и двигательными нейронами, между сегментами спинного мозга, между спинным и головным мозгом; участвуют в обработке информации и выработке команд |
Нейроны составляют лишь 25 % от всех клеток мозга, остальные 75 % клеток относятся к нейроглии (греч. glia — клей).
нейроглия (глия)
Глиальные клетки впервые описал в 1846 г. Рудольф Вирхов. Он считал, что они «склеивают» нервные клетки.
Определение
Нейроглия — совокупность вспомогательных клеток нервной ткани.
Глиальные клетки активно делятся в течение всей жизни, и число клеток глии значительно превышает число нейронов (в 10 раз у взрослого и в 15 раз у пожилого человека). Увеличение массы мозга у ребенка в течение постнатального развития осуществляется за счет увеличения массы дендритов и клеток глии.

Рис. 9. Астроциты нейроглии
Астроциты (зеленым цветом) — один из видов глиальных клеток.
Функции нейроглии:
трофическая функция (питание нейронов);
опорная функция;
транспортная (обмен веществ между кровью и нейронами);
секреторная функция (образование спинномозговой жидкости);
разграничительная функция;
защитная функция (гематоэнцефалический барьер).
Клетки нейроглии
Нейроглия делится на два генетически и функционально разных вида.
Макроглия (глиоциты)
Состав:
Эпендимоциты: выстилают спинномозговой канал и желудочки мозга.
Функция: образование спинномозговой жидкости (ликвора).
Астроциты: имеют извитые отростки, переплетающиеся с отростками нейронов.
Функции:
осуществляет транспорт веществ из крови в нейроны и обратно (астроциты располагаются между капиллярами и телами нейронов);
выполняют опорную функцию, заполняя промежутки между нейронами;
образуют гематоэнцефалический барьер (физиологический барьер между кровеносной системой и ЦНС), защищающий мозг от инфекций и чужеродных веществ.
Олигодендроциты: окружают тела нейронов; входят в состав оболочек нервных волокон и нервных окончаний.

Рис. 10. Клетки нейроглии
Функции:
формируют миелиновую оболочку вокруг аксонов;
участвуют в дегенерации и регенерации нервных волокон;
обеспечивают обмен веществ.
Микроглия
Состав:
Макрофаги: имеют небольшой размер, активно передвигаются, распределены по всей ЦНС.
Функция: фагоцитоз продуктов обмена веществ и погибших нейронов.
нейрогенез
Определение
Нейрогенез — процесс образования нервных клеток.
Нейрогенез включает в себя несколько этапов:
деление (= пролиферация) клеток-предшественниц;
миграция новообразованных клеток в определенный отдел мозга;
дифференцировка новообразованных клеток;
образование нового функционирующего нейрона.
Долгое время ученые полагали, что нейрогенез у млекопитающих происходит только в эмбриональный период («нервные клетки не восстанавливаются»). Однако в последнее время исследования показали, что нейрогенез происходит в течение всей жизни человека.
Функции нервной системы
регуляция жизнедеятельности тканей, органов и их систем;
объединение (интеграция) организма в единое целое;
осуществление взаимосвязи организма с внешней средой и приспособления его к меняющимся условиям среды;
определение психической деятельности человека как основы его социального существования.
Особенности работы нервной системы
В отличие от гуморальной регуляции процессов жизнедеятельности, осуществляемой железами внутренней секреции, нервная система обеспечивает быструю передачу информации (возбуждения) вполне определенным клеткам, тканям, органам.
Отделы нервной системы
Центральная нервная система (ЦНС): головной и спинной мозг.
Периферическая нервная система: нервы, нервные узлы (ганглии), рецепторы.
Определение
Нерв — пучок аксонов.
По виду аксонов нервы делятся на:
чувствительные нервы: из аксонов чувствительных нейронов;
двигательные нервы: из аксонов двигательных нейронов;
смешанные нервы: из аксонов чувствительных и двигательных нейронов.
Соматическая нервная система — часть нервной системы, регулирующая деятельность скелетной (произвольной) мускулатуры.
Вегетативная нервная система — часть нервной системы, регулирующая деятельность внутренних органов, гладкой мускулатуры и обмен веществ.
Строение спинного мозга человека
Спинной мозг имеет вид длинного белого шнура (около 40 см), заостренного внизу. На уровне большого затылочного отверстия он переходит в головной мозг, а на уровне 1–2 поясничного позвонка заканчивается пучком нервов, получившим название «конский хвост».
Расположен спинной мозг в позвоночном канале под защитой позвоночника.

Рис. 1
ОБОЛОЧКИ СПИННОГО МОЗГА
Спинной мозг покрыт тремя оболочками:
твердая оболочка спинного мозга: плотная соединительнотканная оболочка, которая несет кровеносные и лимфатические сосуды; Она не прилегает вплотную к стенкам позвоночного канала, которые покрыты надкостницей;
между надкостницей и твердой оболочкой находится эпидуральное пространство. В нем залегают жировая клетчатка и венозные сплетения;
субдуральное пространство — между твердой и паутинной оболочкой;
паутинная оболочка спинного мозга представлена тонкой полупрозрачной соединительнотканной пластинкой, расположенной кнутри от твердой оболочки; образует сеть перекладин, состоящих из тонких пучков коллагеновых и эластических волокон;
субарахноидальное пространство: между паутинной и мягкой оболочкой. Заполнено ликвором (обеспечивает питание и обмен веществ нервных клеток);
мягкая сосудистая оболочка спинного мозга покрывает поверхность спинного мозга и соединяется с ним кровеносными сосудами, обеспечивая обмен веществ между ликвором и мозгом, а также фиксирует мозг в полости позвоночника зубчатыми связками.

Рис. 2
Кровоснабжение спинного мозга
Сосуды спинного мозга, спускаясь вдоль спинного мозга, соединяются между собой многочисленными ветвями, образуя на поверхности мозга сосудистую сеть. От этой сети отходят веточки, проникающие вместе с отростками мягкой оболочки в вещество мозга.
К лимфатическим сосудам спинного мозга можно отнести периваскулярные пространства вокруг сосудов, сообщающиеся с субарахноидальным пространством.
ФУНКЦИИ СПИННОГО МОЗГА
Рефлекторная функция (находится под контролем головного мозга):
координация простых безусловных рефлексов (коленного рефлекса, отдергивание руки от горячего предмета и т. п.);
координация некоторых вегетативных рефлексов (сосудодвигательных, пищевых, дыхательных, половых, дефекации, мочеиспускания).
Проводниковая функция:
осуществляет связь между спинным и головным мозгом за счет восходящих и нисходящих путей белого вещества. По восходящим путям возбуждение от мышц и внутренних органов передается в головной мозг, по нисходящим — от головного мозга к органам.
СТРОЕНИЕ СПИННОГО МОЗГА
Передняя и задняя продольные борозды делят спинной мозг на две симметричные половинки. В центре проходит спинномозговой канал, в котором находится спинномозговая жидкость (ликвор). Функции ликвора: механическая защита (амортизация) и питание (обмен веществ) спинного мозга.
В средней части спинного мозга около спинномозгового канала расположено серое вещество, на поперечном срезе напоминающее контур бабочки. Серое вещество образовано телами нейронов и дендритами, в нем различают передние и задние рога. Вокруг серого вещества расположено белое вещество, образованное аксонами нервных клеток.
В задних рогах спинного мозга расположены тела вставочных нейронов.
В передних рогах — тела двигательных нейронов.

Рис. 3. Рога спинного мозга
В составе задних корешков в спинной мозг вступают аксоны чувствительных нейронов, тела которых находятся в ганглиях задних корешков, расположенных рядом со спинным мозгом и образующих вздутия. В спинном мозге эти аксоны направляются в задние рога серого вещества, где они образуют синапсы со вставочными нейронами. Последние в свою очередь образуют синапсы с двигательными нейронами (мотонейронами), лежащими в передних рогах спинного мозга, аксоны которых покидают спинной мозг в составе передних корешков.

Рис. 4. 1 — задние корешки (чувствительные нейроны); 2 — спинномозговой смешанный нерв (аксоны чувствительных и двигательных нейронов); 3 — передние корешки (аксоны двигательных нейронов); 4 — спинномозговой нервный узел (скопление тел чувствительных нейронов)
В грудном, верхнепоясничном и крестцовом отделах спинного мозга серое вещество образует боковые рога спинного мозга, содержащие тела нейронов вегетативной нервной системы.
У каждого человека имеется 31 сегмент спинного мозга: 8 шейных; 12 грудных; 5 поясничных; 5 крестцовых; 1 копчиковый.
Номера сегментов спинного мозга не совпадают с номерами позвонков.
По бокам каждого сегмента передние (двигательные) и задние (чувствительные) корешки попарно сливаются, образуя 31 пару спинномозговых смешанных нервов.

Рис. 5
Проводящие пути спинного мозга
Основные характеристики локомоции, т. е. перемещения человека или животного в окружающей среде при помощи координированных движений конечностей, запрограммированы на уровне спинного мозга. Подобные независимые от внешней стимуляции двигательные программы шире представлены в высших двигательных центрах. Некоторые из них (например, дыхание) врожденные, другие же (например, езда на велосипеде) приобретаются в процессе научения.
В спинном мозге действуют восходящие и нисходящие межсегментарные нервные пути, образованные вставочными нейронами. Их тела находятся в сером веществе спинного мозга, а аксоны поднимаются или спускаются на различные расстояния в составе белого вещества, никогда не покидая спинной мозг.
Таким образом спинной мозг осуществляет интегративную (объединяющую) функцию. У млекопитающих возрастает регуляция спинальных функций высшими отделами центральной нервной системы (процесс энцефализации).
Белое вещество спинного мозга состоит из пучков нервных волокон (аксонов), образующих проводящие пути спинного мозга.
Различают три системы пучков:
короткие пучки ассоциативных (вставочных) волокон связывают сегменты спинного мозга, расположенные на различных уровнях;
восходящие (афферентные, чувствительные) пути направляются к центрам головного мозга;
нисходящие (эфферентные, двигательные) пути идут от головного мозга к клеткам передних рогов спинного мозга.
Белое вещество образует продольные тяжи спинного мозга (канатики).

Рис. 6
В белом веществе передних канатиков проходят в основном нисходящие проводящие пути: в боковых канатиках — восходящие и нисходящие; в задних канатиках — восходящие проводящие пути.
Спинальные локомоторные центры
В опытах показано, что болевое раздражение какой-либо конечности спинального животного (экспериментальное животное, у которого разъединены нервные пути между спинным и головным мозгом) вызывает рефлекторные движения всех четырех; если же такая стимуляция продолжается достаточно долго, могут возникнуть ритмичные сгибательные и разгибательные движения не подвергающихся раздражению конечностей. Если такое животное поставить на бегущую дорожку, то при некоторых условиях оно будет совершать координированные шагательные движения, весьма сходные с естественными.
Предполагают, что у человека тоже есть спинальные локомоторные центры. По-видимому, их активация при раздражении кожи проявляется в виде шагательного рефлекса новорожденного. У взрослого человека спинальные локомоторные центры полностью подчинены головному мозгу. Тем не менее активизация локомоторных центров путем интенсивной тренировки лежит в основе различных методик восстановления ходьбы у больных с повреждением спинного мозга.
Строение головного мозга человека
Головной мозг человека занимает всю полость мозгового отдела черепа.
Кости черепа защищают головной мозг от внешних механических повреждений.
От головного мозга отходят 12 пар черепно-мозговых нервов.
оболочки головного мозга
Снаружи головной мозг покрыт тремя оболочками: сосудистой (мягкой), паутинной и твердой. Это те же оболочки, которые защищают спинной мозг. Оболочки спинного мозга переходят в оболочки головного мозга. Все оболочки снаружи выстланы однослойным плоским эпителием.
Мягкая сосудистая оболочка состоит из двух пластинок, между которыми располагаются мозговые артерии и вены. Эта оболочка сращена с тканью мозга, она принимает участие в образовании сосудистых сплетений желудочков головного мозга, продуцирующих спинномозговую жидкость (ликвор).
Гистология сосудистой оболочки
Строма (внутренняя часть) мягкой оболочки представлена рыхлой неоформленной соединительной тканью с большим количеством кровеносных сосудов и нервных волокон. Снаружи строма покрыта однослойным плоским эпителием нейроглиального происхождения — менинготелием. Сосуды стромы, проникающие в мозг, окружены элементами гематоэнцефалического барьера — астроцитами, ножки которых вокруг сосудов формируют непрерывную муфту. Таким образом, ножки астроцитов и их базальная мембрана являются границей между нервной тканью и мозговыми оболочками (наружная глиальная мембрана).
Паутинная оболочка имеет вид тонкой паутины, образованной соединительной тканью, содержит большое количество фибробластов. От паутинной оболочки отходят множественные нитевидные ветвящиеся тяжи, которые вплетаются в мягкую мозговую оболочку, а с другой стороны — выросты, соединяющиеся с твердой оболочкой.
Пространство между паутинной и мягкой сосудистой оболочкой называется субарахноидальным (подпаутинным) пространством. Оно заполнено ликвором.
Функция паутинной оболочки — поддержание биохимического состава и регуляция давления ликвора (способствует оттоку ликвора в сосуды твердой оболочки).
Твердая оболочка выстилает внутреннюю поверхность черепа. С надкостницей твердая оболочка срастается неравномерно, местами образуя эпидуральное пространство, заполненное жировой тканью. Наиболее плотное срастание наблюдается в районе черепных швов, нервных каналов и основания черепа. Содержит большое количество кровеносных сосудов. В отличие от мягкой, твердая оболочка обладает болевой чувствительностью.

Рис. 1. Схема строения мозговых оболочек полушарий головного мозга: 1 — фрагмент кости свода черепа; 2 — твердая оболочка мозга; 3 — паутинная оболочка; 4 — мягкая (сосудистая) оболочка; 5 — головной мозг; 6 — эпидуральное пространство; 7 — субдуральное пространство; 8 — субарахноидальное пространство; 9 — система ликвороносных каналов; 10 — субарахноидальные ячеи; 11 — артерии в ликвороносных каналах; 12 — вены; 13 — струны конструкции, стабилизирующие артерии в просвете ликвороносных каналов: стрелки указывают направление оттока эпидуральной жидкости в наружную (а) и внутреннюю (б) капиллярную сеть твердой мозговой оболочки
Кровоснабжение головного мозга
Кровеносные сосуды, проникающие в ткань головного мозга, идут по каналам, выстланным мягкой мозговой оболочкой. Вокруг крупных сосудов имеется периваскулярное пространство. Оно сообщается с субарахноидальным пространством и содержит ликвор. Вокруг кровеносных капилляров такого пространства нет. Содержимое кровеносных капилляров отделено от ткани головного мозга гематоэнцефалическим барьером.
Гематоэнцефалический барьер
Определение
Гематоэнцефалический барьер (ГЭБ) — это совокупность физиологических механизмов и анатомических образований в центральной нервной системе, участвующих в регулировании состава ликвора.
Существуют два механизма проникновения веществ в клетки мозга:
через ликвор (промежуточное звено между кровью и нервной или глиальной клеткой);
через стенку капилляра (основной путь у взрослого организма).
Проникновение веществ в мозг осуществляется главным образом через кровеносную систему на уровне капилляров — нервная клетка. Регулируя проницаемость клеточной стенки, ГЭБ контролирует динамику поступления в нервные клетки мозга физиологически активных веществ и препятствует поступлению в мозг чужеродных веществ, микроорганизмов и токсинов.
Структура ГЭБ
Основным элементом структуры ГЭБ являются эндотелиальные клетки. Особенностью церебральных сосудов (сосудов головного мозга) является наличие плотных контактов между эндотелиальными клетками.
В структуру ГЭБ также входят перициты (отростчатые клетки соединительной ткани стенки капилляров; способны сокращаться и фагоцитировать) и астроциты. Межклеточные промежутки между эндотелиальными клетками, перицитами и астроцитами нейроглии ГЭБ меньше, чем промежутки между клетками в других тканях организма.
Эти три вида клеток являются структурной основой ГЭБ не только у человека, но и у большинства позвоночных.


Рис. 2. Элементы гематоэнцефалического барьера
Две функции гематоэнцефалического барьера:
регуляторная: поддержание физико-химических показателей мозга в соответствии с его физиологической активностью;
защитная: защита головного мозга от поступления чужеродных и токсичных веществ.
Гематоэнцефалический барьер — важный компонент нейрогуморальной регуляции, т. к. через него реализуется принцип обратной химической связи в организме, например повышение концентрации определенного вещества в крови приводит к снижению проницаемости для него стенок мозговых капилляров.
Регуляция функций гематоэнцефалического барьера осуществляется высшими отделами ЦНС и гуморальными факторами, в т. ч. уровнем обмена веществ нервной ткани.
Черепномозговые нервы
От головного мозга отходят 12 пар черепномозговых нервов.
нерв |
путь |
функции |
I. Обонятельный |
От носа к головному мозгу |
Ощущение запаха |
II. Зрительный |
От глаза к головному мозгу |
Зрение |
III. Глазодвигательный |
От головного мозга к мышцам глаз |
Движения глаз |
IV. Блоковый |
От головного мозга к наружным мышцам глаз |
Движения глаз |
V. Тройничный |
От кожи головы, слизистых оболочек и зубов к головному мозгу; от головного мозга к жевательным мышцам |
Чувствительность лица, скальпа и зубов; жевательные движения |
VI. Отводящий |
От головного мозга к наружным мышцам глаз |
Поворот глаз кнаружи |
VII. Лицевой |
От вкусовых сосочков языка к головному мозгу; от головного мозга к мышцам лица |
Ощущение вкуса; движения мимической мускулатуры |
VIII. Преддверно-улитковый нерв |
От уха к головному мозгу |
Слух; чувство равновесия |
IX. Языкоглоточный |
От глотки и вкусовых сосочков языка к головному мозгу; от головного мозга к мышцам глотки и слюнным железам |
Чувствительность глотки, ощущение вкуса; глотательные движения, слюноотделение |
X. Блуждающий |
От глотки, гортани и органов грудной и брюшной полостей к головному мозгу; от головного мозга к мышцам глотки и органам грудной и брюшной полостей |
Чувствительность глотки, гортани, органов грудной и брюшной полостей; глотание, голосообразование, замедление сердцебиения, усиление перистальтики |
XI. Добавочный |
От головного мозга к определенным плечевым и шейным мышцам |
Движения плечами; повороты головы |
XII. Подъязычный |
От головного мозга к мышцам языка |
Движения языка |
Строение головного мозга
В отличие от спинного мозга серое вещество головного мозга находится на периферии, образуя кору больших полушарий и несколько подкорковых ядер (скоплений нервных клеток). Белое вещество находится в центральной части головного мозга.
В головном мозге различают пять отделов:
продолговатый мозг;
задний (мост и мозжечок);
средний мозг;
промежуточный мозг;
конечный мозг (большие полушария).
Формирование головного мозга в эмбриогенезе
Нервная система всех позвоночных формируется из нервной трубки.
В начальной стадии своего формирования головной отдел мозга образует три мозговых пузыря: передний, средний и задний.
Передний мозговой пузырь разделяется на зачаток большого и промежуточного мозга. Средний пузырь остается неразделенным и дает начало среднему мозгу. Задний мозговой пузырь подразделяется на зачаток мозжечка и моста (metencephalon) и зачаток продолговатого мозга (myelencephalon), без резкой границы переходящий в эмбриональный спинной мозг.Диаграмма, изображающая главные подотделы эмбрионального позвоночного мозга. Эти области позже разделяются на структуры передний мозг, средний мозг и задний мозг.

Рис. 3. Образование мозга из трех мозговых пузырей

Рис. 4. Отделы головного мозга
Наряду с приведенным выше делением на отделы весь мозг разделяют на три большие части:
ствол мозга;
мозжечок;
передний мозг (большие полушария (конечный мозг) и промежуточный мозг).
Ствол мозга
Состав:
продолговатый мозг;
мост;
средний мозг;
промежуточный мозг (мнения ученых расходятся по вопросу принадлежности промежуточного мозга к стволу).
Функции ствола мозга:
рефлекторная: поведенческие рефлексы;
проводниковая: восходящие и нисходящие нервные пути ЦНС;
ассоциативная: обеспечивает взаимодействие спинного мозга, ствола и больших полушарий головного мозга.
Продолговатый мозг
Является продолжением спинного мозга. В отличие от спинного мозга он не имеет метамерного, повторяемого строения, серое вещество в нем расположено не в центре, а в периферических ядрах.
В продолговатом мозге находятся перекресты нисходящих и восходящих путей, ретикулярная формация.
Ретикулярная формация
Определение
Ретикулярная формация — совокупность клеток и нервных волокон, расположенных на всем протяжении ствола мозга и в центральных отделах спинного мозга.

Рис. 5. Ретикулярная формация ствола, ее активирующие структуры и восходящие пути к коре больших полушарий (схема): 1 — ретикулярная формация ствола мозга и ее активирующие структуры; 2 — гипоталамус; 3 — таламус; 4 — кора большого мозга; 5 — мозжечок; 6 — афферентные (чувствительные) пути; 7 — продолговатый мозг; 8 — мост; 9 — средний мозг
Функции ретикулярной формации:
регуляция уровня сознания, например участие в цикле сон/бодрствование;
переработка и передача сенсорной информации лимбической системе;
защитные рефлексы: глотание, кашель, чихание и т. п.;
жизненно важные рефлексы: дыхательный и сосудодвигательный рефлекс;
двигательная активность.

Рис. 6. Продолговатый мозг
Функции продолговатого мозга:
участвует в реализации вегетативных (слюноотделение), соматических, вкусовых, слуховых, вестибулярных рефлексов;
обеспечивает выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, например при глотании и дыхании;
дыхательный и сосудодвигательный центр;
центр регуляции сердечной деятельности.
варолиев Мост
Мост лежит выше продолговатого мозга. Это утолщенный валик с поперечно расположенными волокнами, которые образуют его белое вещество.
Между волокнами расположены скопления серого вещества, которое образует ядра моста. Продолжаясь до мозжечка, нервные волокна образуют его средние ножки.

Рис. 7. Варолиев мост
Функция варолиева моста: передача информации из спинного мозга в отделы головного мозга.
Мозжечок
Мозжечок лежит на задней поверхности моста и продолговатого мозга в задней черепной ямке. Состоит из двух полушарий и червя, который соединяет полушария между собой. Белое вещество мозжечка покрыто корой из серого вещества. Поверхность мозжечка испещрена бороздами. Нервные ядра лежат внутри полушарий мозжечка, масса которых в основном представлена белым веществом.

Рис. 9. Мозжечок
Функции мозжечка:
координация движений;
поддержание мышечного тонуса.
Средний мозг
Средний мозг соединяет задний мозг с промежуточным.
На крыше среднего мозга находится четверохолмие:
2 зрительных холмика — центры ориентировочных рефлексов на зрительные раздражители;
2 слуховых холмика — центры ориентировочных рефлексов на звуковые раздражители.

Рис. 10. Средний мозг
Функции:
сенсорная функция: проведение зрительной и слуховой информации; ориентировочные рефлексы;
проводниковая функция: через него проходят все восходящие пути к вышележащим таламусу, большим полушариям и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу;
двигательная функция: например движение глазных яблок.
Передний мозг включает в себя промежуточный мозг и конечный мозг, состоящий из больших полушарий.
Промежуточный мозг
Состав: гипоталамус, таламус, метаталамус, эпиталамус.

Рис. 11. Промежуточный мозг
Метаталамус — подкорковый центр зрения и слуха.
Эпиталамус — надбугорная область промежуточного мозга.
К эпиталамусу относится эпифиз (шишковидная железа). Это эндокринная железа, функционально связанная с гипофизом и надпочечниками.

Рис. 12. Эпифиз
Функции эпифиза:
развитие половых признаков (особенно в детском и пубертатном возрасте);
регуляция гормональной функции надпочечников (управление выведением калия и натрия из организма);
регуляция сна (синтез гормона мелатонина).
Таламус (зрительный бугор)
В таламусе можно выделить четыре основных ядра серого вещества:
ядро, перераспределяющее зрительную информацию;
ядро, перераспределяющее слуховую информацию;
ядро, перераспределяющее тактильную информацию;
ядро, перераспределяющее чувство равновесия и баланса.
После того как информация о каком-либо ощущении поступила в ядро таламуса, там происходит ее первичная обработка, то есть впервые осознается температура, зрительный образ и т. д.
Функции таламуса:
первичная обработка зрительных, слуховых и вкусовых сигналов;
запоминание;
двигательные реакции: сосание, жевание, глотание, смех;
центр организации и реализации инстинктов, влечений, эмоций.
Повреждение таламуса может привести к амнезии, вызвать тремор (непроизвольную дрожь конечностей в состоянии покоя).
С таламусом связано редкое заболевание, называемое фатальная семейная бессонница.
Гипоталамус
Особенности нейронов гипоталамуса:
чувствительны к составу омывающей их крови;
отсутствует гематоэнцефалический барьер между нейронами и кровью;
способны к нейросекреции пептидов, нейромедиаторов и др.

Рис. 13. Гипоталамус
Функции гипоталамуса:
является главным подкорковым центром регуляции вегетативных функций организма;
способен воздействовать на вегетативные функции организма с помощью гормонов и нервных импульсов;
в гипоталамусе располагаются центры гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости;
является также центром регуляции цикла бодрствование — сон. При этом задний гипоталамус активизирует бодрствование; передний — сон. Повреждение заднего гипоталамуса может вызвать так называемый летаргический сон;
регулирует деятельность гипофиза;
в гипоталамусе и гипофизе образуются нейрорегуляторные пептиды — энкефалины и эндорфины, обладающие морфиноподобным действием и способствующие снижению стресса.
Лабораторные исследования функций гипоталамуса
Олдс описал поведение крыс, которым вживляли электроды в ядра гипоталамуса и давали возможность самостоятельно стимулировать эти ядра. Оказалось, что стимуляция некоторых ядер приводила к негативной реакции. Животные после однократной самостимуляции больше не подходили к педали, замыкающей стимулирующий ток. При самостимуляции других ядер животные нажимали на педаль часами, не обращая внимания на пищу, воду и др.
Исследования Дельгадо во время хирургических операций показали, что у человека раздражение аналогичных участков вызывало эйфорию, эротические переживания. В клинике показано также, что патологические процессы в гипоталамусе могут сопровождаться ускорением полового созревания, нарушением менструального цикла, половой функции.
Раздражение передних отделов гипоталамуса может вызывать у животных пассивно-оборонительную реакцию, ярость и страх.
Раздражение заднего гипоталамуса вызывает активную агрессию, расширение зрачков, повышение кровяного давления, сужение просвета артериальных сосудов, сокращение желчного, мочевого пузырей.
Гипофиз
Это нижний придаток мозга, расположенный в нижней части гипоталамуса.
Гипофиз является одной из важнейших эндокринных желез; в функциональном отношении он тесно связан с гипоталамусом.
В гипофизе различают переднюю долю (аденогипофиз), заднюю долю (нейрогипофиз).

Рис. 14. Гипофиз
Функции гипофиза:
рост;
обмен веществ;
репродуктивная функция.
конечный мозг
Конечный мозг составляет 80 % всей массы головного мозга и покрывает сверху все остальные отделы. Передний мозг состоит из двух полушарий.
Полушария мозга разделены продольной щелью, в углублении которой содержится мозолистое тело, которое их соединяет.

Рис. 15. Большие полушария конечного мозга
Состав полушарий:
серое вещество образует кору полушарий и подкорковые ядра;
белое вещество образует проводящие нервные пути.
Левое полушарие головного мозга управляет правой половиной тела, а правое — левой. Два полушария дополняют друг друга. Общая поверхность коры головного мозга увеличивается за счет многочисленных борозд, которые делят всю поверхность полушария на доли.
Три главные борозды — центральная, боковая и теменно-затылочная — делят каждое полушарие на четыре доли: лобную, теменную, затылочную и височную.

Рис. 16. Строение больших полушарий
Кора головного мозга функционально состоит из трех зон:
сенсорная зона получает сигналы от рецепторов и передает в ассоциативную зону;
моторная зона — управление двигательными актами, адекватными полученной информации;
ассоциативная зона связывает поступающую сенсорную информацию с хранящейся в памяти; сравнивает информацию, получаемую от разных рецепторов. Сенсорные сигналы интерпретируются и передаются в связанную с ней двигательную зону.
АССОЦИАТИВНЫЕ ЗОНЫ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ
У человека ассоциативная зона занимает около 75 % коры головного мозга.
Ассоциативная зона получает и перерабатывает информацию из сенсорной зоны и инициирует целенаправленное осмысленное поведение.
Лобная доля:
произвольные движения;
речь;
регуляция сложных форм поведения;
мышление.
Теменная доля:
восприятие и анализ кожно-мышечных раздражений;
пространственная ориентация;
регуляция целенаправленных движений.
Височная доля:
восприятие слуховых, вкусовых, обонятельных ощущений;
восприятие речи;
память.
Островок (закрытая долька) (расположен в глубине латеральной борозды):
восприятие вкуса.
Затылочная доля:
восприятие и переработкой зрительной информации.
Гиппокамп (подкорковая зона) (парная структура, расположен в глубине височных долей):
перекодировка информации краткосрочной памяти человека для ее последующей записи в долговременной памяти.
Таким образом, ассоциативные зоны участвуют в процессах мышления, запоминания и обучения.
К программе
Разделы конспекта
Рефлекторный механизм функционирования нервной системы
Вегетативная нервная система человека
Биология. Углубленный уровень. 8 класс.
Рефлекторный механизм функционирования нервной системы
В основе работы нервной системы лежит рефлекс.
Определение
Рефлекс — ответная реакция организма на изменения внутренней и внешней среды, осуществляемая при участии центральной нервной системы.
Предположение о рефлекторном характере деятельности высших отделов головного мозга впервые было развито ученым-физиологом И. М. Сеченовым.
Идеи И. М. Сеченова получили развитие в трудах И. П. Павлова, который открыл пути объективного экспериментального исследования функций коры, разработал метод выработки условных рефлексов и создал учение о высшей нервной деятельности.
И. П. Павлов в своих трудах разделил рефлексы на 2 группы:
безусловные рефлексы — рефлексы, которые осуществляются врожденными, наследственно закрепленными нервными путями;
условные рефлексы — это рефлексы, которые осуществляются посредством нервных связей, формирующихся в процессе индивидуальной жизни человека или животного.
Классификация рефлексов
По типу образования:
условные;
безусловные.
По эффекторам:
соматические, или двигательные, — рефлексы скелетных мышц;
вегетативные — рефлексы внутренних органов: пищеварительные, сердечно-сосудистые, выделительные, секреторные и др.
По биологической значимости:
оборонительные, или защитные;
пищевые;
половые;
ориентировочные.
По месту иннервации:
местные рефлексы осуществляются через периферические ганглии вегетативной нервной системы (например, иннервируемые метасимпатическим отделом вегетативной нервной системы). Обладают относительной автономностью от ЦНС;
центральные рефлексы протекают с обязательным вовлечением различных уровней центральной нервной системы (от спинного мозга до коры больших полушарий).
рефлекторная дуга
Рефлексы осуществляются посредством рефлекторной дуги.
Определение
Рефлекторная дуга — это путь, по которому раздражение (сигнал) от рецептора проходит к исполнительному органу.
Структурную основу рефлекторной дуги образуют нейронные цепи, состоящие из рецепторных, вставочных и эффекторных нейронов. Именно эти нейроны и их отростки образуют путь, по которому нервные импульсы от рецептора передаются исполнительному органу при осуществлении любого рефлекса.
Рефлекторная дуга состоит из пяти отделов:
рецептор;
чувствительный (центростремительный) нейрон;
вставочный нейрон;
двигательный (центробежный) нейрон;
эффектор (рабочий орган).
Типы нейронов
Афферентные (чувствительные, сенсорные) нейроны — нейроны, воспринимающие информацию. Как правило, афферентные нейроны имеют большую разветвленную сеть. Это характерно для всех уровней ЦНС. В задних рогах спинного мозга афферентными являются чувствительные нейроны малых размеров с большим числом дендритных отростков, в то время как в передних рогах спинного мозга эфферентные нейроны имеют тело большого размера, более грубые, менее ветвящиеся отростки. Эти различия нарастают по мере изменения уровня ЦНС к продолговатому, среднему, промежуточному, конечному мозгу. Наибольшие различия афферентных и эфферентных нейронов отмечаются в коре больших полушарий мозга.
Эфферентные (двигательные, мотонейроны) нейроны — это нейроны, передающие информацию от нервного центра к исполнительным органам или другим центрам нервной системы. Например, эфферентные нейроны двигательной зоны коры больших полушарий мозга посылают импульсы к мотонейронам передних рогов спинного мозга, т. е. они являются эфферентными для этого отдела коры больших полушарий. В свою очередь мотонейроны спинного мозга являются эфферентными для его передних рогов и посылают сигналы к мышцам. Основной особенностью эфферентных нейронов является наличие длинного аксона, обладающего большой скоростью проведения возбуждения.
Эфферентные нейроны разных отделов коры больших полушарий связывают между собой эти отделы. Такие связи обеспечивают внутриполушарные и межполушарные отношения в процессе обучения, утомления, при распознавании образов и т. д. Все нисходящие пути спинного мозга образованы аксонами эфферентных нейронов соответствующих отделов центральной нервной системы.
Нейроны вегетативной нервной системы, например ядер блуждающего нерва, боковых рогов спинного мозга, также относятся к эфферентным.
Вставочные нейроны (ассоциативные, интернейроны) — нейроны, которые обрабатывают информацию, полученную от афферентных нейронов, и передают ее на другие вставочные или на эфферентные нейроны.
Вставочные нейроны, как правило, имеют аксоны, терминали которых заканчиваются на нейронах своего же центра, обеспечивая прежде всего их взаимосвязь. Одни вставочные нейроны получают активацию от нейронов других центров и затем распространяют эту информацию на нейроны своего центра. Это обеспечивает усиление влияния сигнала за счет его повторения в параллельных путях и удлиняет время сохранения информации в центре. В итоге центр, куда пришел сигнал, повышает надежность воздействия на исполнительную структуру.
Другие вставочные нейроны передают информацию от эфферентных нейронов своего центра обратно в свой же центр, образуя обратные связи. Так организуются сети, позволяющие длительно сохранять информацию в нервном центре.
Вставочные нейроны могут быть возбуждающими или тормозными.
Рецепторы воспринимают раздражение и отвечают на него возбуждением. Рецепторами могут быть отростки чувствительных нейронов или различные рецепторные эпителиальные клетки. Рецепторы расположены в коже, во всех внутренних органах, скопления рецепторов образуют органы чувств (глаз, ухо и т. д.).
Чувствительный нейрон передает возбуждение к центру.
Тела чувствительных нейронов находятся за пределами центральной нервной системы — в спинномозговых нервных узлах.
Через вставочный нейрон в ЦНС происходит переключение возбуждения с чувствительных нейронов на двигательные.
Центры большинства двигательных рефлексов находятся в спинном мозге. В головном мозге расположены центры сложных рефлексов, таких как защитный, пищевой, ориентировочный и т. д. В нервном центре происходит синаптическое соединение чувствительного и двигательного нейрона.
Двигательный нейрон несет возбуждение от центральной нервной системы к рабочему органу. Двигательным называется нейрон, который передает рабочему органу сигнал из центра.
Эффектор — рабочий орган, который осуществляет эффект, реакцию в ответ на раздражение рецептора.
Эффекторами могут быть мышцы, сокращающиеся при поступлении к ним возбуждения из центра, клетки железы, которые выделяют сок под влиянием нервного возбуждения, или другие органы.
простейшая рефлекторная дуга
Простейшую рефлекторную дугу можно схематически представить как образованную всего двумя нейронами — рецепторным и эффекторным, между которыми имеется один синапс.
Такую рефлекторную дугу называют двухнейронной и моносинаптической.
Моносинаптические рефлекторные дуги встречаются весьма редко. Примером их может служить дуга коленного рефлекса.
Двухнейронная рефлекторная дуга:
первый нейрон — чувствительный нейрон, тело которого находится в спинномозговом ганглии;
второй нейрон — двигательный нейрон, тело которого находится в переднем роге спинного мозга.
Дендрит клетки спинномозгового ганглия имеет значительную длину, он следует на периферию в составе чувствительного нерва. Заканчивается дендрит особым приспособлением для восприятия раздражения — рецептором.
Возбуждение от рецептора по нервному волокну центростремительно передается в спинномозговой ганглий. Аксон нейрона спинномозгового ганглия входит в состав заднего (чувствительного) корешка; это волокно доходит до мотонейрона переднего рога и с помощью синапса, в котором передача сигнала происходит при помощи химического вещества — медиатора, устанавливает контакт с телом мотонейрона или с одним из его дендритов. Аксон этого мотонейрона входит в состав переднего (двигательного) корешка, по которому центробежно сигнал поступает к исполнительному органу, где соответствующий двигательный нерв заканчивается двигательной бляшкой в мышце. В результате происходит сокращение мышцы.

Рис. 1. Схема коленного рефлекса
полисинаптические рефлекторные дуги
В большинстве случаев рефлекторные дуги включают не два, а большее число нейронов: чувствительный, один или несколько вставочных и двигательный нейрон. Такие рефлекторные дуги называют многонейронными и полисинаптическими.
Примером полисинаптической рефлекторной дуги является рефлекс отдергивания конечности в ответ на болевое раздражение.

Рис. 2. Рефлекторная дуга соматического рефлекса: 1 — свеча; 2 — рецептор; 3 — дендрит чувствительного нейрона; 4 — тело чувствительного нейрона в спинномозговом ганглии; 5 — аксон чувствительного нейрона; 6 — тело вставочного нейрона; 7 — спинной мозг; 8 — тело двигательного нейрона; 9 — аксон двигательного нейрона; 10 — рабочая мышца
рефлекторная дуга вегетативного рефлекса
Иначе выглядит дуга вегетативного рефлекса.
Афферентный (чувствительный) нейрон находится в корешке спинного нерва. Синапс, соединяющий чувствительный и I эфферентный (двигательный) нейрон, находится в боковых рогах спинного мозга. Синапс между I и II эфферентными нейронами расположен в либо в боковом симпатическом стволе, либо в парасимпатическом ганглии в районе органа-эффектора.
Таким образом, эфферентный путь вегетативного рефлекса состоит из двух эфферентных нейронов. Вставочный нейрон отсутствует.

Рис. 3. Рефлекторные дуги соматического (А) и вегетативного (Б) рефлекса: 1 — рецептор; 2 — чувствительный нейрон; 3 — спинной мозг; 4 — двигательный нейрон; 5 — рабочий орган: мышца, железа; 6 — вставочный нейрон; 7 — вегетативный ганглий
Любая рефлекторная реакция зависит от взаимоотношения двух основных нервных процессов, из которых состоит всякая нервная деятельность, — возбуждения и торможения.
Возбуждение в нервных центрах стимулирует работу определенного органа.
Торможение в нервных центрах замедляет или прекращает работу связанного с ними органа.
В рефлекторных актах непременно участвуют и возбуждение, и торможение.
При рефлекторном сгибании конечности, например, одновременно с сокращением мышц-сгибателей происходит расслабление мышц-разгибателей. При рефлекторном разгибании конечности сокращение мышц-разгибателей неизменно вызывает одновременное расслабление мышц-сгибателей.
Между возбужденными и заторможенными центрами, совместно участвующими в реакции на раздражение, имеются антагонистические отношения. От них существенно зависит нормальное протекание любого рефлекторного акта.
Вегетативная нервная система человека
Вегетативная (автономная) нервная система — отдел нервной системы, регулирующий деятельность внутренних органов, желез внутренней и внешней секреции, кровеносных и лимфатических сосудов.
Вегетативная нервная система иннервирует весь организм, все органы и ткани. Деятельность вегетативной нервной системы не зависит от воли человека. Однако все вегетативные функции подчиняются центральной нервной системе,в первую очередь коре больших полушарий.
Функции:
нервная регуляция функций всех органов и тканей организма (кроме скелетных мышц);
регуляция обмена веществ;
поддержание гомеостаза — постоянства внутренней среды организма;
приспособительные реакции всех позвоночных.
Особенности вегетативной нервной системы:
очаговое расположение в мозге вегетативных нервных центров;
эффекторные (двигательные) нейроны расположены за пределами центральной нервной системы в узлах вегетативных нервных сплетений;
двухнейронный эфферентный нервный путь от мозга до рабочего органа;
отсутствие сегментарности в иннервации;
преобладают немиелинизированные нервные волокна, т.е. скорость проведения нервных импульсов ниже, чем в соматической нервной системе.
строение вегетативной нервной системы

Анатомически и функционально вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую.
Все структуры и системы организма иннервируются волокнами вегетативной нервной системы. Отделы вегетативной нервной системы находятся в относительном функциональном антагонизме, обеспечивая автоматическую регуляцию органов и систем без участия сознания человека.
Важнейшие органы имеют двойную иннервацию. Полые внутренние органы имеют тройную (симпатическую, парасимпатическую и метасимпатическую) иннервацию.
В симпатическом и парасимпатическом отделах имеются центральная и периферическая части.
Центральную часть вегетативной нервной системы образуют вегетативные ядра — тела нейронов, лежащих в спинном и головном мозге. Они осуществляют координацию работы всех трех частей вегетативной нервной системы.
Периферическую часть вегетативной нервной системы образуют отходящие от ядер нервные волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов.
Симпатические и парасимпатические центры находятся под контролем коры больших полушарий и гипоталамуса.
строение и особенности |
симпатический отдел |
парасимпатический отдел |
центральный отдел |
Ядра в боковых рогах спинного мозга:
|
4 ядра в стволе головного мозга:
Ядра во II — IV сегменте крестцового отделе спинного мозга |
периферический отдел |
парный симпатический ствол; нервные сплетения; нервы |
нервные узлы в стенках внутренних органов или рядом с органами; нервы |
медиаторы |
норадреналин |
ацетилхолин |

Симпатический отдел вегетативной нервной системы
Симпатические ядра расположены в спинном мозге на уровне грудных позвонков. Отходящие от ядер нервные волокна заканчиваются за пределами спинного мозга в симпатических узлах, расположенных по бокам позвоночника. От них берут начало нервные волокна, которые подходят ко всем органам.
Симпатическая нервная система усиливает обмен веществ, повышает возбуждаемость большинства тканей, мобилизует силы организма на активную деятельность.
Симпатический отдел возбуждается при воздействии адреналина.
параСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Парасимпатические ядра лежат в продолговатом мозге и в крестцовой части спинного мозга. Нервные волокна от ядер продолговатого мозга входят в состав блуждающих нервов. От ядер крестцовой части нервные волокна идут к кишечнику, органам выделения. Парасимпатические нервные узлы располагаются в стенках внутренних органов или возле органов.
Парасимпатическая система способствует восстановлению израсходованных запасов энергии, регулирует работу организма во время сна.
Парасимпатический отдел нервной системы возбуждается под воздействием ацетилхолина.
метаСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Метасимпатическая нервная система представлена нервными сплетениями и мелкими ганглиями в стенках пищеварительного тракта, мочевого пузыря, сердца и некоторых других органов.
Функция: осуществляет связь между внутренними органами (минуя головной мозг).
Известно, что многие внутренние органы, извлеченные из организма, продолжают выполнять присущие им функции. Например, сохраняется перистальтическая и всасывательная функция тонкой кишки. Такая относительная функциональная независимость объясняется наличием в стенках этих органов метасимпатического отдела вегетативной нервной системы. Метасимпатический отдел обладает собственным нейрогенным ритмом и имеет полный набор необходимых для самостоятельной рефлекторной деятельности звеньев: чувствительный, вставочный и эффекторный нейрон с соответствующим медиаторным обеспечением. В составе этой системы имеются собственные сенсорные элементы (механо-, хемо-, термо-, осморецепторы), которые посылают в свои внутренние сети информацию о состоянии иннервируемого органа, а также способны передавать сигналы в ЦНС. Метасимпатическая нервная система ограничена и охватывает только некоторые внутренние органы. Она не имеет своего центрального аппарата и ее эфферентные связи с центральными структурами опосредованы нейронами симпатического и парасимпатического отделов вегетативной нервной системы.
Существование специальных местных метасимпатических механизмов регуляции функций имеет определенный физиологический смысл. Их наличие увеличивает надежность регуляции функций. Эта регуляция может происходить в случае выключения связи с центральными структурами. При этом ЦНС освобождается от избыточной информации.
Органы с разрушенными метасимпатическими путями утрачивают способность к координированной моторной деятельности и другим функциям.
Влияние симпатического и парасимпатического отделов на отдельные органы
Симпатический отдел:
повышает частоту и силу сердечных сокращений;
стимулирует выброс адреналина;
повышает уровень глюкозы в крови;
повышает артериальное давление;
вызывает расширение артерий головного мозга, легких и коронарных артерий;
угнетает перистальтику кишечника и работу пищеварительных желез (в том числе слюнных), сокращает гладкомышечные сфинктеры;
угнетает перистальтику мочеточников, расслабляет мускулатуру и сокращает сфинктер мочевого пузыря;
расширяет бронхи и бронхиолы, усиливает вентиляцию легких;
расширяет зрачки.
Парасимпатический отдел:
уменьшает частоту и силу сердечных сокращений;
понижает уровень глюкозы в крови;
снижает артериальное давление;
усиливает перистальтику кишечника и стимулирует работу пищеварительных желез (в том числе слюнных), расслабляет гладкомышечные сфинктеры;
усиливает перистальтику мочеточников, сокращает мускулатуру и расслабляет сфинктер мочевого пузыря;
сужает бронхи и бронхиолы, уменьшает вентиляцию легких;
сужает зрачки.
РЕГУЛЯЦИЯ РАБОТЫ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Все механизмы регуляции деятельности внутренних органов условно объединены многоэтажной иерархической структурой.
Первый структурный уровень: внутриорганные рефлексы, имеющие метасимпатическую природу;
Второй структурный уровень: ганглии брыжеечных и солнечного (чревного) сплетений; Оба этих низших этажа обладают отчетливо выраженной автономностью и могут осуществлять регуляцию независимо от центральной нервной системы.
Третий структурный уровень: центры спинного мозга и ствола головного мозга.
Четвертый структурный уровень: гипоталамус, ретикулярная формация, лимбическая система, мозжечок, новая кора (верхний слой больших полушарий, отвечающий за высшую нервную деятельность).
Рефлекторные процессы в ядерных образованиях спинного, продолговатого, среднего мозга и моста находятся под постоянным влиянием гипоталамуса.
Гипоталамические центры: поддержание гомеостаза; регуляция метаболизма; регуляция функций эндокринных желез; интеграция нервной и гуморальной регуляции вегетативных функций (связь с гипофизом).
Лимбическая система («висцеральный мозг»): соматовисцеральная интеграция (объединение работы опорно-двигательной системы и внутренних органов): пищевое, сексуальное, оборонительное поведение, сон и бодрствование, внимание, эмоции, процессы памяти.
Мозжечок: стабилизирующее влияние на деятельность висцеральных (внутренних) органов.
Ретикулярная формация: повышение активности нервных центров, связанных с висцеральными функциями. Высоко чувствительна к гуморальным раздражениям. При стимуляции ретикулярной формации среднего и промежуточного мозга усиливается выброс гипофизарных гормонов, а при нарушении этих ретикулярных образований возникают эндокринные расстройства.
Кора больших полушарий мозга: контролирует работу всех внутренних органов. Известно, что в определенных условиях у человека гипнотическим внушением можно вызвать изменение сердечного ритма, усиление потоотделения и мочеотделения, изменение метаболизма.
Типы секреции. Экзокринные железы
Секреция — синтез и выделение клеткой определенных химических веществ.
У людей, так же как и во всех клетках эукариот, процесс секреции происходит путем экзоцитоза.
Секрет — жидкость, выделяемая клетками и содержащая биологически активные вещества.
С помощью секреции происходит образование и выделение молока, слюны, пота, желудочного, поджелудочного и кишечного сока, желчи, гормонов.
Секреторную функцию выполняют специализированные клетки — гландулоциты.
Гландулоциты — клетки железистого эпителия.
Форма гландулоцитов изменяется в зависимости от фазы секреции. Отличительной особенностью гландулоцитов являются крупные ядра, большое количество митохондрий и секреторные включения в цитоплазме. Хорошо развита гранулярная эндоплазматическая сеть, продуцирующая белковый секрет. В клетках, синтезирующих липиды, лучше развита гладкая эндоплазматическая сеть. Характерно наличие хорошо развитого комплекса Гольджи, где осуществляется формирование секреторных гранул. Важная роль в перемещении секреторных продуктов в гландулоцитах и их выделении принадлежит элементам цитоскелета — микротрубочкам и микрофиламентам.
Фазы секреции:
Поглощение гландулоцитами простых исходных веществ из крови и лимфы для синтеза секрета.
В эндоплазматической сети: синтез и накопление секрета.
В комплексе Гольджи: упаковка секрета в секреторные пузырьки. В секреторных пузырьках могут идти химические преобразования: например, синтез инсулина из проинсулина.
Экструзия — выделение секрета из гландулоцитов — происходит объединение секреторных пузырьков с клеточной мембраной поросомы (органоид, принимающий участие в экзоцитозе). В результате экзоцитоза содержимое пузырька выбрасывается из клетки.
Восстановление исходного состояния железистой клетки.
Периодические изменения железистой клетки, связанные с образованием, накоплением, выделением секрета и восстановлением ее для дальнейшей секреции, получили название секреторного цикла: поступление веществ — синтез и накопление секрета — выведение секрета.
Разновидностью секреции является нейросекреция.
железы
Железы — органы, выполняющие секреторную функцию.
Большинство желез являются производными железистого эпителия.
Железы делят на две большие группы:
экзокринные — железы внешней секреции.
эндокринные — железы внутренней секреции.
Кроме того существуют железы смешанной секреции — например, половые железы и поджелудочная железа — выделяющие секрет по протокам и гормоны непосредственно в кровь.
Экзокринные железы
Паренхима (основная ткань) экзокринной железы состоит из двух отделов: конечного (секреторного) отдела и выводного протока. По выводному протоку секрет экзокринных желез выделяется в полость или на поверхность тела. Экзокринные железы могут быть окружены соединительнотканной капсулой или содержать соединительнотканные перегородки — септы, разделяющие железу на доли и более мелкие дольки. Окружающие и поддерживающие их соединительнотканные элементы называются стромой железы.
В состав секреторного отдела, кроме железистых (секреторных) клеток, могут входить миоэпителиальные (мио — мышца) клетки. Они образуют длинные отростки, охватывающие снаружи концевые отделы. Сокращаясь, миоэпителиальные клетки облегчают продвижение секрета в выводной проток.
По количеству клеток: одноклеточные и многоклеточные (см. рис.).

По местонахождению:
эндоэпителиальные железы: одноклеточные; представлены бокаловидными клетками в составе реснитчатого многорядного эпителия воздухоносных путей (1) и однослойного призматического эпителия кишки (2).
 1.
1.
 2.
2.
экзоэпителиальные железы: многоклеточные; расположены в соединительной ткани (например, потовые железы в дерме) и связаны с эпителием выводными протоками.

По количеству выводных протоков подразделяют
простые: имеют один не ветвящийся выводной проток;
сложные: имеют разветвленный выводной проток.
По количеству секреторных отделов:
неразветвленные: имеют один конечный секреторный отдел;
разветвленные: имеют несколько конечных секреторных отделов (Г);
Сложные железы всегда разветвленные, потому что их многочисленные выводные протоки заканчиваются многими секреторными отделами.
По форме секреторных отделов железы подразделяют на:
трубчатые: конечный отдел имеет форму трубочки;
альвеолярные: конечный отдел имеет форму мешочка;
трубчато-альвеолярные: в железе есть оба типа концевых отделов.

А. Простая трубчатая неразветвлённая;
Б. Простая альвеолярная неразветвлённая;
В. Сложная альвеолярно-трубчатая неразветвлённая;
Г. Простая альвеолярная разветвлённая (несколько секреторных отделов на одном протоке);
Д. Сложная альвеолярная неразветвленная.
По химическому составу секрета экзокринные железы подразделяются на несколько типов:
белковые (или серозные),
слизистые,
белково-слизистые (или смешанные),
сальные,
солевые (например: потовые и слезные).
По способу выделения секрета железы делятся на следующие разновидности:
мерокриновые железы (А): секрет выделяется из клетки без нарушения ее целостности. К ним относятся большинство желез, например, слюнные железы;
апокриновые железы (Б): апикальная часть клетки отторгается вместе с секретом. К апокриновым железам относятся молочные и потовые железы;
голокриновые железы (В): после накопления секрета клетка полностью разрушается и ее остатки включаются в состав секрета. К голокриновым железам относятся сальные железы кожи.



регенерация железистых клеток
В железах в связи с их секреторной деятельностью постоянно происходят процессы физиологической регенерации.
В мерокриновых и апокриновых железах, в которых находятся долгоживущие клетки, восстановление исходного состояния гландулоцитов после выделения из них секрета происходит путем внутриклеточной регенерации или путем размножения.
В голокриновых железах восстановление осуществляется за счет размножения стволовых клеток. Вновь образовавшиеся из них клетки путем дифференцировки превращаются в железистые клетки.
В пожилом возрасте изменения в железах могут проявляться снижением секреторной активности железистых клеток и изменением состава вырабатываемых секретов, а также ослаблением процессов регенерации и разрастанием соединительной ткани.
виды экзокринных желез
В коже человека находятся потовые и сальные железы. Молочные железы — видоизмененные потовые железы — функционируют в период грудного вскармливания.
Кожные железы обеспечивают терморегуляцию (около 20% тепла отдается организмом путем испарения пота), защиту кожи от повреждений (жировая смазка предохраняет кожу от высыхания, а также от мацерации водой и влажным воздухом), обеспечивают выведение из организма продуктов обмена веществ (мочевина, мочевая кислота, аммиак и др.).
Потовые железы являются производными кожного эпителия. Больше всего потовых желез на коже лица, ладоней, ступней, подмышечных впадин и паховых складок.
Состав пота: 98% воды и 2% органических и неорганических веществ.
Функции:
терморегуляция за счет выделения воды на поверхность кожи;
выделение продуктов белкового обмена;
выделение солей (хлорида натрия и др.);
выделение специфических пахучих веществ.
По механизму секреции потовые железы подразделяются на эккриновые (мерокриновые) и апокриновые:
эккриновые железы распространены в коже повсеместно; функция — в основном терморегуляция.
апокриновые железы находятся лишь в определенных местах кожного покрова, например в подмышечных впадинах, коже лба, области заднего прохода, гениталиях; развиваются в период полового созревания организма. Разновидностью апокриновых потовых желез являются железы век и железы, выделяющие ушную серу.
Выводные протоки многих апокриновых желез впадают вместе с выводными протоками сальных желез в волосяные воронки.

Секрецией апокриновых потовых желез управляют половые железы (например, у женщин секреция апокриновых желез возрастает в предменструальный период и во время беременности).
Сальные железы достигают наибольшего развития во время полового созревания. Секретируют они по голокриновому типу.
Выводные протоки сальных желез связаны с волосами. Около одного корня волоса можно встретить до трех сальных желез. Больше всего сальных желез на голове, лице и верхней части спины. На ладонях и подошвах они отсутствуют.
Секрет сальных желез — кожное сало.

Функции:
уход за кожей и волосами: смягчает, придает эластичность;
облегчает трение соприкасающихся поверхностей кожи;
препятствует развитию микроорганизмов.
Концевые отделы состоят из двух видов себоцитов: малоспециализированных, способных к митотическому делению клеток, и клеток, находящихся в разных стадиях жирового перерождения. Первый вид клеток образует наружный (или базальный) ростковый слой концевого отдела. Кнутри от него располагаются более крупные клетки, в цитоплазме которых появляются капли жира. Постепенно процесс синтеза жира в них усиливается и одновременно клетки смещаются в сторону выводного протока. В результате удаления от источника питания, происходит гибель клеток, под влиянием ферментов лизосом клетки распадаются и превращаются в кожное сало. Последнее по выводному протоку поступает в воронку волоса и далее на поверхность его стержня и эпидермиса кожи.
Молочные железы по своему происхождению представляют видоизмененные кожные потовые железы.
Молочные железы закладываются у зародыша на 6—7-й неделе в виде двух уплотнений эпидермиса (т.н. «молочные линии»), тянущихся вдоль туловища. Из этих утолщений формируются так называемые «молочные точки». Затем они разветвляются на своих дистальных концах, формируют зачатки молочных желез. В течение детского возраста у обоих полов происходит разрастание разветвлений молочных ходов.
С наступлением половой зрелости возникают резкие половые различия в темпах развития молочных желез. У мальчиков образование новых ходов замедляется и затем прекращается. У девочек развитие железистых трубок значительно ускоряется и к началу менструаций на молочных ходах появляются первые концевые отделы. Однако молочная железа достигает окончательного развития только при беременности в период лактации. Интенсивная секреция полноценного молока устанавливается в течение первых дней после рождения ребенка.

Молоко — сложная водная эмульсия, в состав которой входят жировые капли, белки (казеин и др.), углеводы (лактоза, или молочный сахар) и соли.
Опорожнению альвеолы и переходу молока в млечные ходы способствует сокращение миоэпителиальных клеток.
По окончании периода лактации молочная железа претерпевает инволюцию, однако часть альвеол, образовавшихся во время предыдущей беременности, сохраняется.
Регуляция деятельности функционирующей молочной железы осуществляется двумя основными гормонами:
пролактин (гормон гипофиза) стимулирует железистые клетки альвеол (лактоциты) к биосинтезу молока, накопляющегося сначала в млечных ходах;
окситоцин (гормон гипоталамуса) стимулирует выведение молока из млечных протоков во время лактации.
Кроме кожных желез в организме человека есть пищеварительные и половые железы.
К пищеварительным железам относятся слюнные, желудочные, кишечные, поджелудочная железа и печень.
Слюнные железы расположены в ротовой полости и относятся к пищеварительной системе.
Выделяют три пары крупных слюнных желез и множество мелких железок, расположенных на языке и небе.
околоушные слюнные железы расположены ниже уровня уха;
подчелюстные слюнные железы расположены возле углов нижней челюсти;
подъязычные слюнные железы находятся под краями языка.

Слюнные железы секретируют слюну.
Состав слюны:
99 % — вода;
ферменты (амилазы): первичное переваривание углеводов;
лизоцим: антибактериальные и ранозаживляющие свойства.
Функции слюнных желез:
выделение пищеварительных ферментов;
защита слизистой оболочки полости рта;
формирование пищевого комка и стимуляция глотания.
Печень — самая крупная железа в организме человека. Расположена в брюшной полости и имеет дольчатое строение. Способна к регенерации.
Функции печени:
очистка крови от токсических веществ;
участие в процессе пищеварения: секретирует желчь, которая эмульгирует жиры и усиливает перистальтику кишечника;
принимает участие в метаболизме жирорастворимых витаминов;
депонирует кровь и гликоген;
в плодный период участвует в кроветворении.
Поджелудочная железа является железой смешанной секреции.
Вырабатывает панкреотический сок, содержащий ферменты для расщепления питательных веществ в тонком кишечнике.
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
Эндокринные железы не имеют выводных протоков, их продукты — гормоны — выделяются непосредственно в кровь или лимфу.
Гормоны — высоко активные биологические вещества, выполняющие регуляторную функцию.
ЭВОЛЮЦИЯ ЭНДОКРИННОГО АППАРАТА
Развитый эндокринный аппарат возник вслед за нервной системой на ранних этапах эволюции животного мира. При этом наиболее ранней формой эндокринных клеток, по-видимому, стали нейросекреторные нейроны , выявляемые отчетливо уже у кишечнополостных и имеющиеся у представителей более развитых типов беспозвоночных и всех позвоночных.
Можно предположить, что у древнейших примитивных metazoa нервные клетки выполняли двоякую функцию: контактно-медиаторную и нейросекреторную.
Поскольку эпителиальные эндокринные железы не могут быть обнаружены у большинства типов беспозвоночных и встречаются только у представителей некоторых моллюсков, ракообразных и насекомых, а также всех классов позвоночных, предполагается, что этот тип желез - более позднее достижение эволюции.
Вероятнее всего, эволюционное развитие эндокринной системы животных складывалось из следующих основных этапов:
появление нейросекреторных клеток;
скопление нейросекреторных клеток в определенных участках головного мозга и некоторых других ганглиев;
объединение нейросекреторных клеток в органы;
формирование из эпителиальных клеток эндокринных желез, чувствительных к нейрогормонам.
Следует отметить, что однотипные эндокринные железы закономерно встречаются внутри каждой крупной таксономической группы высокоорганизованных беспозвоночных и внутри всей группы позвоночных.
Развитие желез
А — Б — эпителиальные клетки усиленно размножаются и образуют вырост, постепенно углубляющийся в соединительную ткань.
В — формирование экзокринной железы: клетки в области верхушки выроста дифференцируются в секреторные, а остальные формируют выводной проток железы.
Г — Д — формирование эндокринной железы: при разрыве связи с эпителиальным пластом.

Эндокринная железа состоит из скоплений эндокринных клеток, окружённых соединительной тканью с многочисленными кровеносными капиллярами.
Два варианта организации эндокринной железы: островок и фолликул. В последнем случае гормоны из эндокринных клеток поступают в просвет фолликула, где они хранятся до введения в кровь.
регуляция секреции
Регуляция секреции идет через нервные и гуморальные механизмы. При этом в железистых клетках активизируются ферментные системы и метаболизм, сборка микротрубочек и сокращение микрофиламентов, участвующих во внутриклеточном транспорте и выведении секрета.
Секреция слюнных желез регулируется преимущественно нервными (рефлекторными) механизмами; секреция желез желудка — нервными и гуморальными механизмами;
секреция поджелудочной железы регулируется гормонально.
Между нейронами и железистыми клетками могут образовываться синапсы: некоторые нервные окончания выделяют нейромедиаторы в межклеточное пространство, откуда медиатор путем диффузии попадает к секреторным клеткам.
Секреторные клетки реагируют и на местные факторы: рН среды, пищевые продукты, отдельные компоненты секретов и т.д. Их значение особенно велико в регуляции деятельности желез пищеварительного тракта.
Эндокринные железы
Эндокринная система — система регуляции деятельности внутренних органов посредством гормонов, выделяемых эндокринными клетками непосредственно в кровь, лимфу или спинномозговую жидкость.
Гормоны — биологически активные вещества органической природы, вырабатывающиеся в специализированных клетках желёз внутренней секреции, поступающие в кровь и оказывающие регулирующее влияние на обмен веществ и физиологические функции.
Гормоны служат гуморальными регуляторами физиологических процессов в различных органах и системах.
По химическому составу гормоны делятся на:
стероидные гормоны (гормоны коркового слоя надпочечников и половые гормоны);
производные аминокислот (тироксин, адреналин, норадреналин);
пептидные гормоны (рилизинг-гормоны; инсулин, глюкагон, гормон роста).
Свойства гормонов:
высокая физиологическая активность;
каждый гормон регулирует определенный процесс;
каждый гормон действует на определенный орган-мишень.
Восприятие гормонов органами
Все гормоны воздействуют на отдельные органы-мишени при помощи специальных рецепторов к этим гормонам.
Если гормоны плохо проникают через клеточную мембрану (производные аминокислот и пептидные гормоны):
гормон — рецепторы мембраны — посредники: (простагландины) — фермент аденилатциклаза катализирует синтез цАМФ из АТФ — синтез необходимых белков-ферментов.

Если гормон легко проходит через клеточную мембрану (стероидные гормоны):
гормон — внутриклеточный рецептор — гормон-рецепторный комплекс — синтез белков-ферментов.

Все ткани и органы имеют механизм обратной связи, который участвует в саморегуляции восприимчивости органов к гормонам:
при низком уровне определённого гормона автоматически возрастает количество рецепторов в тканях и их чувствительность к этому гормону повышается;
при высоком уровне определённого гормона происходит автоматическое понижение количества рецепторов в тканях и их чувствительности к этому гормону понижается.
Увеличение или уменьшение выработки гормонов, а также снижение или увеличение чувствительности гормональных рецепторов и нарушение гормонального транспорта приводит к эндокринным заболеваниям.
Эндокринная система включает:
центральное звено: гипоталамус и гипофиз. Функция: регуляция работы эндокринных желез.
периферическое звено: эндокринные железы и эндокринные клетки. Функция: регуляция работы организма.
В эндокринную систему входят как специализированные железы (например, щитовидная железа, надпочечники), так и неспецифические органы, содержащие эндокринные клетки (диффузная эндокринная система): печень, почки, желудок, кишечник, сердце.
Хорошо изучена гормональная функция желудка и двенадцатиперстной кишки. В них синтезируются гормоны — регуляторы функций желудка, поджелудочной железы и печени (гастрин, секретин и др.). В печени синтезируется соматомедин, стимулирующий соматотропный гормон (соматотропин = гормон роста).
Простагландины, образующиеся практически во всех тканях организма, участвуют в регуляции внутриклеточного обмена веществ.
Функции эндокринной системы
гуморальная регуляция функций организма;
координация работы всех органов и систем;
гомеостаз организма при изменяющихся условиях внешней среды;
рост и развитие организма;
половая дифференцировка и репродуктивная функция;
обмен веществ и энергии;
эмоциональные реакции;
психическая деятельность человека.
нейрогуморальная регуляция
Выполняя роль регулятора физиологических функций, эндокринная система является составной частью более сложной системы нейрогуморальной регуляции.

эндокринные железы

гипоталамус
Гипоталамус входит в систему гипоталамус — гипофиз — надпочечники, где он выполняет роль высшего подкоркового эндокринного регулятора.
Одна из функций гипоталамуса — нейросекреция: выделение нервными клетками гипоталамуса физиологически-активных веществ (рилизинг-гормонов), регулирующих работу гипофиза.
Рилизинг-факторы:
статины — тормозят работу гипофиза;
либерины — стимулируют работу гипофиза.
Например, соматолиберин стимулирует, а соматостатин — наоборот, тормозит — выработку гипофизом соматотропного гормона (гормона роста).
гипофиз
Гипофиз анатомически и функционально тесно связан с гипоталамусом.
Гипофиз состоит из двух долей:
передняя доля — аденогипофиз
состоит из железистой ткани
связана с гипоталамусом сетью кровеносных сосудов
регулируется рилизинг-гормонами
задняя доля — нейрогипофиз
состоит из нервной ткани
связана аксонами с гипоталамусом
Гормоны передней доли гипофиза
Соматотропин (СТГ = гормон роста) стимулирует синтез белков, деление клеток, обмен веществ.
Гиперфункция: гигантизм (1) — рост тела выше 2,0 м; акромигалия (2) — патологическое увеличение отдельных частей тела.
Гипофункция: задержка роста и физического развития (гипофизарный нанизм = гипофизарная карликовость (1): рост мужчин — до 1,30 м, рост женщин до 1,20 м).
 1.
1.
 2.
2.
Группа тропных гормонов:
гонадотропные гормоны (ГТГ) стимулирую секреторную функцию половых желез;
тиреотропный гормон (ТТГ) увеличивает продукцию гормонов щитовидной железы;
адренокортикотропныйгормон (АКТГ) усиливает синтез адреналина корой надпочечников.
гормоны задней доли гипофиза (нейрогипофиза)
Вазопрессин (АДГ = антидиуретический гормон) усиливает реабсорбцию воды в почечных канальцах.
Гипофункция: несахарный диабет (симптом: жажда и усиление диуреза до 15 л мочи в сутки).
Гиперфункция: повышение артериального давления.
Окситоцин регулирует тонус мускулатуры матки и молочных желез.
надпочечники
Гормоны коркового слоя надпочечников
Половые гормоны вырабатываются надпочечниками на протяжении всей жизни человека. В детском возрасте и после наступления климактерического периода только надпочечники вырабатывают половые гормоны.
андрогены — стероидные мужские половые гормоны.
эстрогены — стероидные женские половые гормоны.
прогестерон — стероидный гормон жёлтого тела яичников.
Глюкокортикоиды регулируют углеводный обмен.
кортизон обладает противовоспалительной активностью.
кортикостерон и дегидрокортикостерон повышают уровень глюкозы в крови.
Минералкортикоиды регулируют водный и минеральный обмен.
альдостерон усиливает реабсорбцию ионов натрия и выведение ионов калия с мочой.
Общим предшественником кортикоидных и половых гормонов является холестерин.
Гормоны мозгового слоя надпочечников
адреналин и норадреналин оказывают выраженное стимулирующее влияние на мышечную работоспособность; стимулируют синтез стероидных гормонов.
Гипофункция коры надпочечников: бронзовая, или болезнь Аддисона возникает при недостатке кортикоидных гормонов (симптомы: хроническая усталость, истощение, раздражительность, гиперпигментация открытых частей тела).
 Болезнь Аддисона.
Болезнь Аддисона.
щитовидная железа
Расположена в передней части шеи в виде бабочки.
Вес: 20 — 30 г.
Синтезирует йодсодержащие гормоны: тироксин и трийодтиронин.
Тироксин (Т4) и трийодтиронин (Т3) регулируют обмен веществ, рост и развитие организма.
Активность трийодтиронина в десятки раз выше тироксина.
Тиреокальцитонин регулирует кальциевый обмен: поступление кальция из крови в костную ткань.
Гипофункция (гипотериоз): микседема (слизистый отек). Симптомы: в следствие нарушения белкового обмена возникает слизистый отек тканей; снижается обмен веществ; задерживается психическое развитие, угнетается половая функция.
Эндемический зоб — разрастание железистой ткани — возникает при недостатке йода в продуктах питания.
 Микседема.
Микседема.
 Эндемичный
зоб.
Эндемичный
зоб.
Гипофункция щитовидной железы в детском возрасте приводит к кретинизму — задержке роста и психического развития, инфантилизму; в более тяжелых случаях — к идиотии.
Гиперфункция щитовидной железы (тиреотоксикоз):
Базедова болезнь: увеличение щитовидной железы, увеличение скорости обмена веществ, астения, раздражительность, пучеглазие. Энергетические расходы организма при работе увеличиваются в 2 — 3 раза.
 Базедова
болезнь.
Базедова
болезнь.
Паращитовидные железы
Расположены симметрично на боковой поверхности щитовидной железы.
Гормон: паратиреоидин (паратгормон) возбуждает функцию остеокластов (костеразрушающих клеток) и способствует переходу кальция из костей в кровь. Является антагонистом тиреокальцитонина щитовидной железы.
Гипофункция паращитовидных желез: нарушение роста и развития костной ткани, скелета, зубов. Дефицит кальция в крови приводит к нарушению функций ЦНС и печени.
Гиперфункция паращитовидных желез: разрушение костной ткани (остеопороз), мышечная слабость, нарушение функций внутренних органов.
Тимус (вилочковая = зобная железа)
Функционирует как эндокринная железа до наступления половой зрелости, тормозя преждевременное половое созревание.
У половозрелого человека она представляет орган лимфопоэза человека: гормон тимозин регулируют созревание, дифференцировку и иммунологическое «обучение» Т-лимфоцитов.
Рост органа продолжается до начала полового созревания (в это время его размеры максимальны (до 7,5 — 16 см в длину), а масса достигает 20 — 30 грамм). С возрастом тимус подвергается атрофии и в старческом возрасте едва отличим от окружающей его жировой ткани.
Гипофункция тимуса: снижение иммунитета.
Поджелудочная железа
Расположена слева в районе желудка.
Гормоны регулирует углеводный обмен:
инсулин увеличивает способность клеточных мембран пропускать углеводы: глюкоза в виде гликогена запасается в клетках, т. о. снижается уровень глюкозы в крови;
глюкагон — прямой антагонист инсулина; усиливает выход гликогена из клеток в кровь и распад его до глюкозы, т. о. повышает уровень глюкозы в крови.
Гипофункция поджелудочной железы: сахарный диабет. Сахар не усваивается клетками, уровень глюкозы в крови возрастает и она выводится с мочой; недостаток сахара в клетках приводит к судорогам, потери сознания (диабетической коме) и смерти.
Половые железы
Мужские половые железы: семенники.
Женские половые железы: яичники.
До начала пубертатного периода мужские и женские половые гормоны вырабатываются примерно в одинаковых количествах у мальчиков и у девочек. К моменту наступления половой зрелости у девушек увеличивается секреция женских половых гормонов, а у юношей — мужских.
Мужские гормоны (андрогены) и женские гормоны (эстрогены) вызывают появление вторичных половых признаков.
Тестостерон — мужской половой гормон — регулирует развитие вторичных половых признаков, сперматогенез, уменьшает синтез гликогена в печени.
Эстрогены регулируют менструальный цикл и течение беременности.
Прогестерон, или гормон желтого тела (ЛГ) подготавливает стенку матки к имплантации оплодотворенной яйцеклетки, стимулирует развитие молочных желез; регулирует развитие беременности в ранние сроки (до 3 — 4 месяцев).
Эпифиз (шишковидная железа)
Находится в промежуточном мозге.
Вырабатывает гормоны мелатонин и серотонин.
Функции серотонина:
снижает болевую чувствительность;
нейромедиатор в ЦНС;
свертывание крови;
является исходным веществом для синтеза мелатонина.
Функции мелатонина:
торможение выделения гормонов роста;
торможение полового развития и полового поведения;
торможение развития опухолей;
влияние на половое развитие и сексуальное поведение.
У детей эпифиз имеет бо́льшие размеры, чем у взрослых; по достижении половой зрелости выработка мелатонина уменьшается.
Разрушение эпифиза приводит к преждевременному половому созреванию.
Опорно-двигательная система человека. Скелет
Опорно-двигательный аппарат состоит из костей скелета, их соединений и мышц.
Функции опорно-двигательного аппарата:
опорная: является опорой всего тела; к костям прикрепляются мягкие ткани и органы;
двигательная: система рычагов с подвижными соединениями, приводимых в движение мышцами;
защитная: образует полости для жизненно важных органов — позвоночный канал для спинного мозга; черепная коробка — для головного мозга; грудная полость — для сердца и легких; тазовые кости — для защиты органов мочеполовой системы;
минеральный обмен: кости являются депо для минеральных солей: фосфора, кальция, железа, меди; регулируют постоянство минерального состава внутренней среды организма;
кроветворная (гемопоэтическая функция): из стволовых гемопоэтических клеток костного мозга образуются клетки крови и иммунной системы.
Химический состав костной ткани
В состав костной ткани входят:
органические вещества (оссеин): придают костям гибкость и упругость;
неорганические вещества (вода, соли кальция, магния, фосфаты): минеральные соли придают костям твердость.
Оссеин — органическое вещество костной ткани.
В состав оссеина входят белки (коллаген и др.), небольшая доля липидов (лецитин и др.) и углеводов (гликоген).
Коллаген — основной белок костной ткани.
Специфической особенностью костной ткани является содержание в ней значительного количества солей лимонной кислоты — 70% от всего запаса ее в организме, что обусловлено особенностями биосинтеза ткани.
В детском возрасте количество органических веществ максимально, кости детей упругие, устойчивы к переломам, однако легко деформируются при чрезмерных нагрузках.
С возрастом количество органических веществ уменьшается, а доля минеральных солей увеличивается. Кости приобретают твердость и прочность.
У пожилых людей в костях уменьшается доля минеральных веществ, из-за этого их кости становятся более хрупкими.
При сжигании кость чернеет с выделением углерода, который остаётся после разложения органических веществ.
В растворах кислот минеральные соли костной ткани растворяются — остается оссеин, и кости становятся пористыми и эластичными, но сохраняют свою форму.

При удалении органических веществ путем сжигания кость также сохраняет первоначальную форму, но становится хрупкой и легко крошится.
Только правильное сочетание органических и неорганических веществ делает кость твердой и упругой. Прочность скелета значительно возрастает благодаря сложной архитектуре внутреннего строения костей.
строение костей
В состав скелета человека входит более 200 костей (у новорожденного — более 300 костей). Точное количество костей определить невозможно, т.к. в детском возрасте продолжается замена хрящевых частей костными. Количество копчиковых позвонков у людей варьирует от 3 до 5.
микроскопическое строение костей
Различают три типа клеток костной ткани: