
Липиды
.pdf
|
|
|
ЛИПИДЫ |
|
|
|
|
|
|
|||
Терминлипидыобъединяеторганическиевещества,(масла, жирыили воска), |
||||||||||||
обладающие общимфизическимсвойством-гидрофобностью,т.е.нерастворимостьюв |
||||||||||||
воде,онихорошорастворяютсяворганическихрастворителяхПоструктурелипиды |
||||||||||||
настолькоразнообразны,чтоунихотсутствуетобщийпризнакхимическогостроения. |
||||||||||||
|
Липидыимеютдвеглавныефункции-1. |
|
|
-влипидахзапасаетсябольшая |
||||||||
часть энергии, выделяющейся в результате химических реакций. Основную массу таких |
||||||||||||
|
|
|
|
|
|
|
Запастопл ва |
|
|
|
|
|
липидов в организме составляютжиры — три-ацилглицеролы (триглицериды), служащие |
||||||||||||
формой депонирования энергии. Жиры располагаются преимущественно в подкожной |
||||||||||||
жировой ткани и выполняют также функции теплоизоляционной и механической |
||||||||||||
защиты. |
|
- липидысоставляютглавныекомпонентыклеточныхмембран,кним |
||||||||||
2. |
|
|
||||||||||
относятся Фосфолипиды—большойкласслипидов,получившийсвоёназваниеиз-за |
||||||||||||
|
Структурная |
|
|
|
|
|
|
|
|
|
|
|
остаткафосфорнойкислоты,придающегоимполярныесвойства.Благодаряэтому |
||||||||||||
свойствуфосфолипидыформируютбислойнуюструктурумембран. |
|
|
|
|||||||||
|
Нарушения обмена липидов приводят к развитию многих заболеваний, но среди |
|||||||||||
людей наиболеераспространеныдваизних—ожирение иатеросклероз. |
|
|||||||||||
СТРУКТУРА,СОСТАВ ИСВОЙСТВАЖИРНЫХкислот |
|
|
|
|
||||||||
Жирныекислотысамыераспространенныеинеобходимыекомпонентыбольшинства |
||||||||||||
липидов,Жирныекислоты–этодлинноцепочечныеорганическиекислоты,содержащиеот |
||||||||||||
4до24углеродныхатомов.Жирные кислоты представляютсобойуглеводородную |
||||||||||||
неразветвлённую цепь,наодномконце которойнаходитсякарбоксильнаягруппа, |
||||||||||||
(-СООН)ана другом—метильнаягруппа(-СН3).Из-задлинногонеполярного«хвоста» |
||||||||||||
большинстволипидовнерастворимывводеипроявляютсвойствамасел илижиров. |
||||||||||||
Жирные кислоты,не содержащиедвойныхсвязей,называют |
|
|
.при |
|||||||||
температуретелаонинаходятсяв |
|
|
состоянии. Основнойнасыщенной |
|||||||||
|
|
|
|
|
|
|
|
|
|
насыщенными |
|
|
жирнойкислотой влипидах человекаявляетсяпальмитиновая,содержащая16 |
||||||||||||
|
|
|
|
|
|
твердом |
|
|
|
|
|
|
атомовуглерода(до30—35%).Жирные кислоты, содержащие двойные связи, |
|
|||||||||||
называют |
|
|
Они представляютсобой |
|
|
Ненасыщенные |
||||||
жирные кислотыпредставленымоноеновыми(соднойдвойнойсвязью)и |
|
|||||||||||
|
|
ненасыщ нными. |
|
|
|
|
|
жидкости. |
|
|
||
полиеновыми(с двумяибольшимчислом двойных связей). Все встречающиесяв |
||||||||||||
естественныхусловияхжирные кислотысодержатчетное число атомовуглерода, 16 |
||||||||||||
или18углеродныхатомов. Общую формулу жирныхкислот можновыразитьтак: |
||||||||||||
насыщеннаяжирнаякислота: |
|
CH3(CH2)n (CH=СН)m.COOH |
|
|
|
|||||||
ненасыщеннаяжирнаякислота: |
CH3(CH2 )n COOH |
|
|
|
|
|||||||
Какправило,ненасыщенные жирные кислотывстречаютсяиу животных,и у растенийвдваразачаще,чемнасыщенные.
Дополнительные двойные связиобычнорасположенымежду 9-ойдвойнойсвязью и метильнымконцомцепи.
Есливсоставе жирнойкислоты содержатся две иболее двойных связей,тоони располагаютсячерез СН2-группу.
Двойные связи–CHв жирных= CH –CHкислотах–CH= CHв орга– низме человека имеют цис-конфигурацию. Этоозначает, что ацильные2фрагменты находятся пооднусторонудвойнойсвязи.Цисконфигурация двойной связи делает алифатическую (открытую) цепь жирной кислоты изогнутой, что нарушает упорядоченное расположение насыщенных радикалов жирных кислот в фосфолипидах мембраниснижаеттемпературуплавления.Чем больше двойных связей в жирных кислотах липидов, тем ниже температура их плавления. чем длиннее углеводороднаяцепь,темвышетемператураплавленияжирныхкислот.
Жирные кислоты взаимодействуют с щелочами (NaOH и KOH), вследствие чего образуются соли жирных кислот, имеющие свойства мыла. Мыла обладают амфипатическими свойствами, т.е. одна их часть гидрофильна, а другая (углеводородный хвост) полностью гидрофобна. Мыла формируют гидрофильную оболочку вокруг капель жира, образуя мицеллу. Углеводородные хвосты мыла встраиваются в капли жира, а полярные головы взаимодействуют с водой. Мыла способныэмульгироватьнерастворимыевводемаслаижиры.
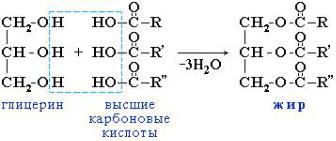
1. КЛАССЫЛИПИДОВ : Представляют.
собойсложныенейтральныеэфирыжирытрехатомного- Триацилглспиртацерины–глицерина, или триглицеридыи трех молекулжирных кислот (сложный эфир - это вещество, которое образуются при взаимодействии органической кислотыиспирта)
где |
,остаткижирныхкислот |
|
|
Триглицериды – это |
, гидрофобные вещества, поскольку они не содержат |
||
|
R,R',R” |
|
. |
заряженных или сильнонеполярныеполярных функциональных групп. В их состав часто входят разные жирные кислоты, насыщенные и ненасыщенные. Живые организмы запасают «топливо» в виде триглицеридов. В растительных триглицеридах преобладают ненасыщенные жирные кислоты, поэтому они являются жидкими при комнатной температуре, (подсолнечное. оливковое). Животные жиры содержат в составе триглицеридов преимущественно насыщенные жирные кислоты и поэтому имеют твердую консистенцию. В растительных клетках триацилглицеролы находятся в цитозоле в виде мелкодисперсных эмульгированных маслянистых капелек. У позвоночных животных в жирной ткани имеются специализированные клетки, адипоциты, которые полностью заполнены триглицеридами. Во многих животных, обитающих в холодных условиях (пингвины. моржи, тюлени) триглицериды образуют толстыйтеплоизоляционныйслой,защищающийорганизмотпереохлаждения.
2. – это сложные эфиры, образуемые длинноцепочечными (14-36 C), насыщеннымиВоска или ненасыщеннымижирнымикислотами и длинноцепочечными спиртами (счисломуглеродныхатомовот16до30).
У позвоночных воска выполняют функцию защитного покрытия, смазывающего и смягчающего кожу, а также предохраняющую перья, шерсть, мех от воды. Воска имеют высокуютемпературуплавления.Книмотносятсяпчелиныйвоск,спермацет,ланолини т.д.
3. основные липидные компоненты мембран. Структурная основа глицерофосфолипидовФосфолипиды - — глицерол. Глицерофосфолипиды представляют собой молекулы, в которых с глицеролом в первой и второй позициях связаныдвежирные кислотысложноэфирнойсвязью,а втретьейпозициинаходится остатокфосфорной кислоты, к которому, в свою очередь, могут быть присоединены различные заместители,чаще всегоаминоспирты.

можетбыть: этаноламин,холин,Хсеринили
инозитол. Жирные кислотывсоставе фосфолипидовбывают16или18углеродными, однанасыщенная,а другаяненасыщенная, последняя обычносвязянасовторымуглеродом глицерола.
Поскольку в составе фосфолипидов остаток фосфорной кислоты заряжен |
||||||||||||
отрицательно, также заряжены спиртовые остатки, каждая молекула фосфолипида |
||||||||||||
состоит из полярной (взаимодействующей с водой) «головы» и двух неполярных |
||||||||||||
хвостов из длинноцепочечных жирных кислот. т.е. Все фосфолипиды являются |
||||||||||||
|
|
|
|
|
, поэтому они участвуют в строении мембран. Благодаря |
|||||||
своим полярным свойствам фосфолипиды не только являются основой всех |
||||||||||||
полярными соединениями |
|
|
|
|
|
|
|
|
||||
клеточных мембран, но и выполняют другие функции: образуют поверхностный |
||||||||||||
гидрофильный слой липопротеинов крови, выстилают поверхность альвеол, |
||||||||||||
предотвращая слипание стенок во время выдоха. Некоторые фосфолипиды |
||||||||||||
участвуютвпередачегормональногосигналавклетки |
их |
|
|
|
|
, которые |
||||||
4. Еще один класс полярных липидов - |
|
основе |
е |
|
|
|||||||
также- |
являются важными компонентами мембран. В |
|
|
трехатомный |
||||||||
спирт глицерин, |
|
|
Сфинголипиды гликолипиды |
|
||||||||
как в случае фосфолипидов а аминоспиртсфингозин.Сфинголипиды |
||||||||||||
построены из одного остатка длинноцепочечной жирной кислоты, одного остатка |
||||||||||||
длинноцепочечного аминоспирта сфингозина и одного остатка спирта полярной головы. |
||||||||||||
Гликолипиды добавочно содержат остатки сахаров (галактозу). Гликосфинголипиды |
||||||||||||
локализуются в основном на поверхности клеток и участвуют в процессе «узнавания» |
||||||||||||
клеткамидругдруга,вадгезии,атакжеопределяютгруппукрови. |
|
|
|
четырех |
||||||||
5. |
|
|
- |
класс липидов, которые являются |
производными |
|||||||
конденсированныхциклическихсистем —циклопентанпергидрофенантренов. |
||||||||||||
|
Стероиды |
|
|
|
|
|
|
|
|
|
|
|
|
В организме человека основной стероид — холестерол,остальные стероиды —его |
|||||||||||
производные. Растения, грибы и дрожжи не синтезируют холестерол, но образуют |
||||||||||||
разнообразные фитостеролы и микостеролы, не усваиваемые организмом человека. |
||||||||||||
Бактериине способнысинтезироватьстероиды. |
|
|
|
|
|
|
||||||
|
Холестерол входит в состав мембран и влияет на структуру бислоя, увеличивая её |
|||||||||||
жёсткость. Из холестерола синтезируются жёлчные кислоты, стероидные гормоны - |
||||||||||||
мужские и женские половые гормоны, гормоны надпочечников, а также. |
витамин D3. |
|||||||||||
Нарушение обменахолестеролаприводитк развитию атеросклероза |
|
|
|
|
||||||||
Холестеролпредставляетсобоймолекулу,содержащую 4 конденсированныхкольца, обозначаемые латинскими буквами А, В, С,D, разветвлённую боковую цепь из 8 углеродных атомовв положении 17, 2 метильные группы (18 и 19) и гидроксильную группу в положении 3. Наличие
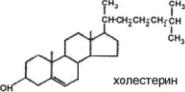
гидроксильной группы позволяет относить холестерол к спиртам, поэтому его правильное химическое название «холестерол», однако в медицинской литературе часто используют термин«холестерин».
Присоединение жирных кислот сложноэфирной связью к гидроксильной группе приводит к образованиюэфиров холестерола.
В неэтерифицированной форме холестерол входит в состав мембран различных клеток. Гидроксильная группа холестерола обращена к водному слою, а жёсткая гидрофобная часть молекулыпогруженавовнутреннийгидрофобныйслоймембраны
В крови 2/3 холестерола находится в этерифицированной форме и 1/3 — в виде свободного холестерола. Эфиры холестерола служат формой его депонирования в некоторых клетках (например, печени, коры надпочечников, половых желёз). Из этих депо холестерол исполь-
зуетсядлясинтезажёлчн |
х |
|
истероидных горм нов. |
|
|||
|
|
|
|
кислоты |
|
липопротеидами. |
|
Полярные липиды, жирные |
|
|
|||||
|
, холестерол и его эфиры образуют комплексы |
||||||
со |
специфическими |
белками, |
|
которые называются |
|
||
Липопротеиды имеют внешнюю гидрофильную оболочку, благодаря чему они |
|||||||
растворимы в воде и приспособлены для транспорта липидов в плазме крови |
|||||||
Холестерин и триглицериды циркулируют в плазме крови в составе липопротеинов. |
|||||||
Гидрофобное ядро липопротеинов содержит .триглицериды и эфиры холестерина, а |
|||||||
оболочка -фосфолипиды, холестеринибелки |
|
||||||
Липопротеиды плазмы различаются липидным составом и, соответственно, |
|||||||
функциями. Определение липопротеидного состава крови имеет диагностическое |
|||||||
|
Терпены |
|
|
|
|
|
|
значение дляпревенцииатеросклероза. |
|
||||||
6. |
|
— класс природных липидов, которые в больших количествах |
|||||
содержатся в растениях. Терпены не растворимы в воде, но хорошо растворимы в |
|||||||
неполярных растворителях и жирах. Терпены и их производные летучие вещества, |
|||||||
они входят в состав эфирных масел, придают характерный запах плодам, цветкам и |
|||||||
листьям растений. Основой терпенов является изопрен. Производными изопрена |
|||||||
являются такжежирорастворимые витамины A,Eи K. |
|
||||||
эйкозаноиды |
|
жирной |
кислоты, арахидоновой кислоты, происходят |
||||
7. |
Из 20 углеродной |
||||||
– гормоноподобные вещества, обладающие высокой биологической активностью, причем они не переносятся кровью. а действуют местно. К ним относятсяПростагландины, Тромбоксаны, Лейкотриены.
Простагландины вызывают сокращение или расслабление гладких мышц, регулируют артериальное давление. Синтез простагландинов возрастает во время воспалительных процессов. Они вызывают повышение температуры тела и болевые ощущения. Тромбоксаны способствуют агрегации тромбоцитов. они участвуют в процессе свертывания крови. Лейкотриены синтезируются в лейкоцитах. Они стимулируютподвижность,хемотаксисимиграцию лейкоцитов вочаг воспаления.
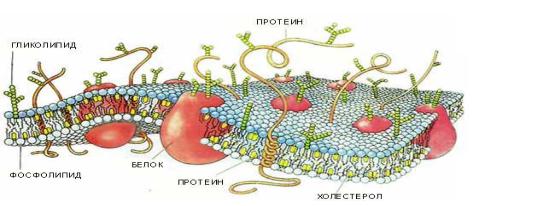
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса |
|||||||||||||
липидов, большинство из которых представляет собой так называемые сложные |
|||||||||||||
липиды— |
фосфолипиды. |
Молекулы фосфолипидов |
имеют |
гидрофильную |
|||||||||
(«головка») игидрофобную («хвост») часть. При образовании мембран гидрофобные |
|||||||||||||
участкимолекулоказываютсяобращены внутрь,а гидрофильные — наружу. |
|
||||||||||||
Биологическая мембрана, кроме этого состоит из |
, молекулы которых как бы |
||||||||||||
«растворены» в |
липидном |
бислое. |
Основной |
взаимодействующей силой между |
|||||||||
мембранными |
макромолекулами |
является |
белков |
|
|
|
. |
||||||
Ковалентные связи |
|
образуются ни между липидами, |
ни между липидами и |
||||||||||
белками. |
|
|
|
не |
|
|
|
|
гидрофоб ое взаимодействие |
|
|||
|
|
|
БИОЛОГИЧЕСКАЯМЕМБРАНА |
|
|
|
|
||||||
Как и мыла, полярные липиды обладают амфипатическими свойствами и в воде |
|||||||||||||
спонтанно формируют мицеллы. Полярные липиды растекаются по поверхности |
|||||||||||||
растворов |
|
|
|
|
|
|
|
|
|
|
бислои. |
||
образуя слой толщиной в одну молекулу – монослой. На поверхности |
|||||||||||||
Поля ные |
липидные |
бислои |
|
|
|
|
|
|
|
||||
раздела |
двух водных фаз они самопроизвольно формируют очень тонкие |
|
|
||||||||||
мембран. |
|
|
|
|
|
|
составляют Структурную |
основу |
биологических |
||||
Основные принципы структурной организации всех мембран одинаковы, однако одна из |
||||||||||||||||||
самых характерных особенностей — огромное их разнообразие. Мембраны органелл |
||||||||||||||||||
эукариотических клеток уникальны по своему составу и по характеру выполняемых |
||||||||||||||||||
функций. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Липидныйсоставмембранразличен,содержаниетогоилидругоголипида,по- |
|||||||||||||||||
|
|
|
|
|
|
|
амфифильные |
|
|
молекулы, |
|
|||||||
видимому,определяетсяразнообразием функций,выполняемых этимилипидамив |
||||||||||||||||||
мембранах. Мембранныелипиды— |
|
|
|
|
|
(амфипатические) |
|
|
|
|
т.е.в |
|||||||
молекулеестькакгидрофильные группы(полярные «головки»), таки |
|
|
|
|||||||||||||||
алифатические радикалы(гидрофобные «хвосты»),самопроизвольноформирующие |
||||||||||||||||||
бислой.Вбольшинствеэукариотическихклетоконисоставляютоколо30—70% |
|
|||||||||||||||||
фосфолипиды,гликолипиды ихолестерол(холестерин). |
|
|
|
|
|
|||||||||||||
массымембраны.Вмембранахприсутствуютлипидытрёхглавныхтипов— |
— |
|||||||||||||||||
|
Наиболее |
распространённые |
|
глицерофосфолипиды |
|
мембран |
||||||||||||
фосфатидилхолины и фосфатидилэтаноламины. Специфические фосфолипиды |
||||||||||||||||||
внутренней мембраны митохондрий — кардиолипины (дифосфатидилглицеролы), |
||||||||||||||||||
построенные на основе глицерола и |
в |
двух остатков |
фосфатидной кислоты. В |
|||||||||||||||
плазматических |
|
мембранах |
клеток |
|
значительных |
количествах содержатся |
||||||||||||
сфингомиелины. Сфингомиелины — главные липиды миелиновой оболочки нервных |
||||||||||||||||||
волокон. |
|
|
гидрофобная часть представлена церамидом. Гидрофильная группа — |
|||||||||||||||
углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у |
||||||||||||||||||
|
В гликолипидах |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
первого углеродного атома церамида. В зависимости от длины и строения углевод- |
||||||||||||||||||
нойчасти различают |
|
|
|
содержащиемоноили олигосахаридный остаток, |
||||||||||||||
и |
|
|
|
к |
-группе которых |
присоединён |
сложный, |
разветвлённый |
||||||||||
|
|
|
|
|
цереброзиды, |
|
|
|
|
|
|
|
|
|
|
|
||
олигосахарид. Полярные «головки» гликосфинголипидов находятся на наружной |
||||||||||||||||||
|
ганглиозиды, |
|
|
ОН |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
поверхности плазматических мембран. Ганглиозиды являются важными |
||||||||||||||||||
компонентами расположенных на поверхности мембран специфических |
|
|
|
|||||||||||||||
|
потому что их отрицательно заряженные полярные группы (головы), служат |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
рецепторных |
||
выступающими на поверхность клетки «антеннами», распознающими. |
молекулы |
|||||||||||||||||
участков, |
сигнальных |
веществ, в |
|
частности гормонов |
В |
|
значительных |
|||||||||||
определенных |
|
|
||||||||||||||||
количествах гликолипиды содержатся в мембранах клеток мозга, эритроцитов, эпителиальных клеток. Ганглиозиды эритроцитов разных индивидуумов различаютсястроениемолигосахаридныхцепей,проявляющихантигенные свойства.
поверхность
|
|
|
|
|
|
поверхность |
|
Мембраны животных клеток также содержат холестерол. Единственная гидроксильная |
|||||||
группа его является «полярной головкой». Так как молекула холестерола состоит из |
|||||||
жёсткого гидрофобного ядра и гибкой углеводородной цепи, наличие холестерола в |
|||||||
мембранах уменьшает текучесть мембран, латеральную диффузию липидов и белков, |
|||||||
и поэтому может влиять на функции мембранных белков. В составе мембран |
|||||||
растений холестерола нет, а присутствуют растительные стероиды — ситостерол и |
|||||||
стигмастерол. |
|
|
|
|
|
|
|
Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю |
|||||||
поверхности, различающиеся по липидному и белковому составам — эту |
|||||||
особенность |
мембран называют |
|
(поперечной) |
|
|
||
Липиднаяасимметриявозникаетпреждевсегопотому,чтолипиды сболее объёмными |
|||||||
|
|
|
|
тран мембранной |
|
асимметрией. |
|
полярными «головками» стремятся находиться в наружном монослое, так как там |
|||||||
площадь поверхности, приходящаяся на полярную «головку», больше. |
|||||||
Фосфатидилхолины и сфингомиелины локализованы преимущественно в наружном |
|||||||
монослое, а фосфатидилэтаноламины и фосфатидилсерины в основном во |
|||||||
внутреннем. Липиды в некоторых биологических мембранах с довольно большой |
|||||||
частотой мигрируют с одной стороны мембраны на другую, т.е. совершают «флип- |
|||||||
флоп» (от англ. |
|
перескоки. Перемещение липидных молекул затрудняют |
|||||
полярные «головки», поэтому липиды, находящиеся на внутренней |
стороне |
||||||
|
|
flip—flop) |
|
|
|
|
|
мембраны, имеют относительно высокую скорость трансмембранной миграции по |
|||||||
сравнению с липидами наружной стороны мембраны, мигрирующих медленнее или |
|||||||
вообщене совершающими«флип-флоп» перескоки. |
|
|
|
||||
Для мембран характерна жидкостность (текучесть), которая, в свою очередь, |
|||||||
определяется относительным содержанием насыщенных и ненасыщенных жирных |
|||||||
кислот в составе липидов. Микровязкость меньше, если в составе липидов |
|||||||
преобладают ненасыщенные жирные кислоты, и больше при высоком содержании |
|||||||
насыщенных жирных кислот. Ацильные (алифатические) остатки ненасыщенных |
|||||||
жирных кислот имеют так называемые«изломы». Эти «изломы» препятствуют |
|||||||
слишком плотной упаковке молекул в мембране и делают её более рыхлой, а |
|||||||
следовательно и более «текучей». На текучесть мембран также влияют размеры |
|||||||
углеводородных «хвостов» липидов, с увеличением длины которых мембрана |
|||||||
становитсяболее «текучей». |
|
|
|
|
|||
Из за текучести мембран возможна латеральная диффузия липидов и белков: В |
|||||||
латеральной |
плоскости |
(т.е. на одной стороне |
бислоя) свободно |
могут |
|||
перемещаться как молекулы липидов, также и мембранные белки. Скорость перемещениямолекулзависитотмикровязкостимембран.
Е ли основная роль липидов в составе мембран заключается в стабилизац.Одниизних обеспечиваютбислоя,тобелкитранспортотвечаютопределённыхза функциональнуюмолекулактивностьи ионовмембран, другие являются ферментами, третьи участвуют в связывании цитоскелета с внеклеточным матриксом или служат рецепторами для гормонов и медиаторов. На долю белков приходится от 30 до 70% массы мембран. Белки определяют особенностифункционированиякаждой мембраны.
Белки мембран различаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его — интегральные белки, либо разными способами прикрепляться к мембране — поверхностные белки. Поверхностные белки или домены интегральных белков, расположенные на наружной поверхности всех мембран,почтивсегда гликозилированы(присоединены с остатками сахаров). Олигосахаридные остатки защищают белок от протеолиза, участвуютвузнаваниилигандовилиадгезии.
ФУНКЦИИМЕМБРАНЫ: |
|
|
|
|
|
I.Барьерная –мембранаотделяетклеткуотвнешнейсредыи делитее на |
|
||||
компартменты, предохраняяотвытекания ионовивеществ,отпроникновения |
|||||
токсиноввирусови т.д. |
|
|
|
|
|
II.Транспортнаяфункция. Мембраныимеютизбирательную проницаемость.Через |
|||||
нее вклеткупоступаютнеобходимые ейвеществаивыделяютсяпродуктыобмена. |
|||||
Любая молекула может пройти через липидныйбислой,однакоскорость |
|
||||
|
веществ, т.е. перехода вещества из области с большей концентрацией в |
||||
областьсменьшей,можетсильноотличаться. |
|
пассивной |
|||
диффузии |
|
|
|
через липидную мембрану малые |
|
Легче всего проходят |
|
|
|||
неполярные молекулы, такие как 02, стероиды, тиреоидные гормоны, а также жирные |
|||||
|
|
простой |
диффузией |
|
|
кислоты. Малые полярные незаряженные молекулы — С02,NH3, Н20, этанол, мочевина |
|||||
— также диффундируют с достаточно большой скоростью. Диффузия глицерола идёт |
|||||
значительно медленнее, а глюкоза практически не способна самостоятельно пройти |
|||||
через мембрану. |
|
|
|
|
|
|
Для всех заряженных молекул, независимо от размера, липидная |
||||
Транспорт таких молекул возможен благодаря наличию в мембранах либо белков, |
||||||
мембрананепроницаема. |
|
|
(поры), заполненные водой, через которые |
|||
формирующих в липидном слое |
|
|
||||
могут проходить вещества определённого размера простой диффузией, либо |
||||||
|
|
кана ы |
|
|
с |
|
специфических белков-переносчиков, которые избирательно взаимодействуя( |
||||||
определёнными лигандами, облегчают их перенос через мембрану |
облегчённая |
|||||
диффузия). |
|
|
|
|
|
|
Кроме пассивного транспорта веществ, в клетках есть белки, активно |
||||||
перекачивающие определённые растворённые в воде вещества против их |
||||||
|
активным тран портом, |
|
|
|
|
|
градиента, т.е. из меньшей концентрации в область большей. Этот процесс, на- |
||||||
зываемый |
|
|
осуществляется всегда с помощью белков- |
|||
переносчикови происходитсзатратойэнергии. |
|
|
||||
III.Ферментативная функция –большинствоферментоввклетке связаныс |
||
мембранами.Например,в мембранахмитохондрий сосредоточеныферменты |
||
дыхательнойцепи,в мембранахэндоплазматическогоретикулума –ферменты |
||
обменаксенобиотиков,синтезабелкаит.д. |
|
|
IV.Рецепторнаяфункция.Наповерхностиклеточныхмембранимеютсярецепторы |
||
(длягормонови другихрегуляторов).Благодаряналичию рецептороворганизм |
||
получает возможностьрегулировать функцию клеток, |
|
|
|
Похимическойприроде рецепторы |
|
|
|
благодаря рецепторам |
являютсябелками –чаще гликопротеинами.Многие заболеваниясвязаныс |
||
клетки о мениваются информацией. |
|
|
патологиейрецепторов. |
|
|
Например,еслижировые клетки имеютмало рецепторов дляадреналина,то это вызывает накоплениежира(поскольку адреналинактивируетлиполиз).Есликлеткиутрачивает рецепторыдлявеществ-регуляторов,которыетормозятихразмножение,то такиеклетки стают злокачественнымиибесконтрольноразмножаются.Механизмдействиямногих микробныхтоксинов связанстем,что они взаимодействуютсклеточнымирецепторами. Например,холерныйтоксин,связываетсяс рецептораминаэпителиальныхклетках слизистойкишечника;активируятранспортводывкишечник,обуславливаяпонос.
Многие лекарственные веществадействуютнаклеточныерецепторы.Например,морфин соединяетсясрецепторамив нервныхклеткахмозга.Вобычныхусловияхэтирецепторы связываютнейрогормоны,которыерегулируютэмоциональное состояние человека.Тоесть
морфинимитируетдействие нейрогормонов.
.Все многоклеточные организмыимеют неповторимую химическуюV.Антигеннаяиндивидуальностьфункция ,котораязакрепленагенетически,агенетическая чистотаконтролируетсясистемойиммунитета.Поверхностьвсехклетоксодержит множествоантигенов.
Например намембране эритроцитовимеетсяболее 250антигенов,которые определяю группыкрови,трансплантационныйиммунитет.
Антигенамиэритроцитов являютсямембранные белки –гликопротеины,а спецификуантигенаопределяетсостав углеводнойцепочки.В эритроцитах группы кровиВ этацепочказаканчиваетсяглюкозой,ав эритроцитах группы кровиА цепочкадлиннее наодиностаток N- ацетилгалактозамина,отщепив который можно превратитьэритроцитгруппы кровиАв эритроцитгруппы кровиВ.
1VI.Электрическая.Специализированныевозбудимостьфункциим–это передачамбран:информациис помощью изменения зарядамембран.Характернадлянервнойи мышечнойткани.
2.Синаптическаяпередача –преобразование электрическогоимпульсавхимический всинаптических мембранах.
3.Фоторецепция –мембраныспомощью специальногобелка –родопсина превращают световую энергию вхимическую,апотом –вэлектрическийимпульс. 4.Энергоспрягающаяфункция –мембраны митохондрий превращаютэнергию градиентаконцентрациипротоновобусловленаэнергию связейАТФ.
5.Функция подвижности – выростами мембраны: жгутиками, ворсинками.
6.Функциямежклеточныхконтактов.Благодарящелевымконтактам осуществляется переходвеществигормоновизоднойклеткив другую,что обеспечиваетсинхронностьработыклеток. Межклеточные контакты – обеспечиваетсябелкомфибронектином.
ПЕРЕНОСВЕЩЕСТВ ЧЕРЕЗМЕМБРАНЫ.Видытранспорта:
. осуществляетсязасчетградиентаконцентрации (1такПростаяпереносятся(па сивнаяО2, СО)2,диффузияН2Ои другие небольшие молекулы).Лучшедиффундируют те вещества,которые растворяютсявлипидахмембран.Например,диэтиловый эфир–человеквдыхаетчерезлегкие,ауже черезсекундыэфир проникаетвклетки мозга.
2.Облегченнаядиффузия осуществляетсяпоградиентуконцентрации,носболее
|
б лками-переносчиками |
|
||
высокойскоростью.Онаобусловлена: |
||||
а) |
|
|
|
–транслоказы. Белки-транслоказысоединяютсяс |
веществомнаоднойстороне мембраны,переносятегочерезмембрануиотдаютна |
||||
другой.Так, транспортерглюкозывэритроцитахускоряеттранспортглюкозы в10– |
||||
|
ионнымикан лами. |
|
|
|
100млн.раз. |
|
Ионные каналы –этобелковые образования,которые |
||
б) |
|
|
||
обеспечиваюттранспортионовNa+,K+,Ca2+. Например,натриевыйканалвнервных |
||||
клеткахсостоитиздвухбелков. Одинбелокобразуетионную пору,котораяимеет |
||||
селективныйфильтр дляNa+,а другойбелоквыполняетфункцию ворот -закрывает |
||||
илиоткрываетвходв канал. Воротаканалаоткрываютсязасчетэлектрического |
||||
импульса.Некоторые токсиныблокируютнатриевые каналы.Например, |
||||
тетрадотоксинрыбы фугу. Онзакрываетвходвканал(аналогичнокакбутылка |
||||
|
ионофорами. |
|
|
|
закрывается пробкой). |
|
|
||
в) |
|
Ионофоры–этоантибиотики,имеющие циклическую структуру,в |
||
середине кольцаесть полость,черезкоторую избирательномогутпроходитьионы. Например,валиномицинизбирательнопропускаетК+ (встраиваясьвмембрану,он образуетканал,через которыйпроисходит утечкаК+); амфотерицин –избирательно пропускаетанионы.
3. Активныйтранспорт осуществляется противгтранпортныхдиентаконцентрацииза
асчет)ПереносэнергииNa+АТФ,K+, Ca: 2+,анионовипротонов засчет АТФ-аз.Хорошо изученаNa+,K+-АТФ-аза –Na+,K+-насос,обеспечиваетперенос3-хионовнатрияиз клетки наружу вобменнапоступление внутрьклетки двухионовкалия,приэтом гидролизуется 1молекулаАТФ.ИмеетсяпротоннаяАТФ-аза,котораязасчет энергииАТФсоздаетградиент.
б)Симпортиантипорт –этотранспортодноговеществазасчетградиента концентрациидругоговещества. Симпорт – однонаправленный переносодного веществазасчетградиентаконцентрациидругого.Например, переносNa+ через мембранусопровождается транспортомвтомже направлении глюкозыили аминокислот(таквсасываетсяглюкозаи аминокислоты вкишечнике). Антипорт – противоположное движение ионов,вхождение одноговещества вызываетвыброс другого.Например,вхождение вклеткуNa+ сопровождается выбросом Ca2+–. переносвеществвместе счастью плазматической мембраныпутем образования4.Эндоцитозпузырьков:
-пиноцитоз – перенос пузырьков жидкости; -фагоцитоз –перенос твердыхнерастворимыхвеществ.