

Календарный и биологический возраст, их соотношение.
Для точной оценки индивидуального развития рекомендуется учитывать возраст человека. Это связано с тем, что для каждого организма характерны только ему присущие темпы развития. Именно поэтому сроки отдельных возрастных этапов биологического развития не всегда совпадают с возрастом.
Если попытаться дать определение возрасту, то его можно обозначить как длительность существования организма от момента рождения до настоящего времени. Существует понятие календарного (паспортного) и биологического возраста. Календарный возраст измеряется количеством вращений Земли вокруг Солнца и выражается в календарной шкале. Однако, календарный возраст не может служить в полной мере показателем здоровья, трудоспособности, качества жизни лиц одного и того же года рождения. Хорошо известно, что даже внешний облик людей одного календарного возраста бывает настолько разным, что ориентировочное определение возраста может быть ошибочным.
В связи с этим для оценки жизнеспособности, функционального состояния организма используется не календарный, а биологический возраст, более адекватно отражающий меру возрастных изменений организма, его биологические возможности в отношении предстоящей жизни, темпы старения.
Под биологическим возрастом понимают достигнутый уровень морфологического и функционального развития организма. Биологический возраст оценивается по комплексу показателей: физическому развитию (рост, вес и т. д.), срокам окостенения скелета ("костный возраст"), степени половой зрелости, показатели артериального давления, состояние аккомодации зрения, слуховой порог и др.
Фактором, который еще более осложняет точную характеристику истинного возраста, является процесс, получивший наименование акселерация. Этот процесс характеризуется следующими основными особенностями: ускоренным физическим развитием, более ранними сроками полового созревания, увеличением размеров тела.
У долгожителей биологический возраст обычно меньше календарного. Если же биологический возраст опережает календарный, то говорят о преждевременном старении. Последнее является одной из важных проблем геронтологии.
Поэтому наряду с детьми-акселератами, т. е. ускоренно развивающимися, есть дети-ретарданты, развивающиеся медленно, которые отстают в физическом и половом развитии. Поэтому нередко один и тот же календарный возраст объединяет разный в биологическом отношении контингент детей.
В процессе физического воспитания и спортивного совершенствования необходимо учитывать не только календарный, но и биологический возраст занимающихся, индивидуальные особенности развития.
Критерии биологического возраста. При определении биологического возраста учитывают достигнутый отдельным индивидом уровень развития морфологических структур в сравнении со средним хронологическим возрастом той группы, которой он соответствует по уровню своего развития. Индивидуальные колебания процессов роста и развития послужили основанием для введения такого понятия, как биологический возраст или возраст развития.
Основными критериями биологического возраста являются:
зрелость, оцениваемая по степени развития вторичных половых признаков;
скелетная зрелость (порядок и сроки окостенения скелета)
зубная зрелость (сроки прорезывания зубов, стертость зубов);
4) показатели зрелости отдельных физиологических систем организма па основании возрастных изменений микроструктур различных органов;
5) морфологическая и психологическая зрелость.
Оценка биологического возраста производится при сопоставлении соответствующих показателей развития обследованного индивида со стандартами, характерными для данной возрастной, половой и этнической группы.
Половая зрелость. Оценивается на основании степени развития половых признаков, таких как волосы на лобке: Волосы в подмышечных впадинах: Набухание сосков у мальчиков :Выступание кадыка: Перелом голоса: Развитие молочных желез у девочек:.
Скелетная зрелость. Основными показателями возрастной дифференцировки костей скелета являются центры окостенения, сохранность или закрытие эпифизарных хрящевых зон роста, у младенцев — зарастание родничков на голове. У взрослых оцениваются сопряженные с процессом старения изменения компактного вещества кости (истончение), губчатого вещества (разрежение). В периоде старения как критерий биологического возраста используются проявления остеопороза и остеосклероза, остеофитоза (разрастания костной ткани в области фаланг кисти, стопы и на позвонках).
Оссификация скелета тесно связана с биологическим развитаем всего организма (длина тела, темпы роста, стадии полового созревания), а ее стадии ее приурочены к определенным; этапам онтогенеза.
Для оценки скелетной зрелости чаще всего используется определение индексов запястья и различных трубчатых костей на основе рентгенограмм.
Зубной возраст. Определяется на основании числа прорезавшихся и находящихся в состоянии замены молочных зубов у ребенка, у' взрослых — по состоянию стертости постоянных зубов.
Морфологическая зрелость. Оценивается на основании развития сопорно-мышечного аппарата — мышечной силы, выносливости, частоты движений.
Под психологической готовностью ребенка к школе, понимается уровень морфологического и функционального развития, при котором требования системы обучения, нагрузки и новый режим дня не будут обременительны для будущего школьника,
С морфологической и психологической зрелостью тесно связана школьная зрелость, под которой подразумевают уровень психофизиологической и морфологической зрелости, достаточный для рначала школьного обучения.
Лекция 4
Тема: «Возрастная анатомия и физиология нервной системы.
Общий план строения нервной системы.»
План.
Регуляторные системы организма
Общий план строения нервной системы.
Нейроглия
Структура нейрона, его свойства
Строение и свойства нервных волокон.
Связь между нейронами.
Регуляторные системы организма
Нервная система обеспечивает взаимосвязь между отдельными органами и системами органов и функционирование организма как единого целого. Она регулирует и координирует деятельность различных органов, приспосабливает деятельность всего организма как целостной системы к изменяющимся условиям внешней и внутренней среды. С помощью нервной системы осуществляются восприятие и анализ разнообразных раздражений из окружающей среды и внутренних органов, а также ответные реакции на эти раздражения. Вместе с тем следует иметь в виду, что вся полнота и тонкость приспособления организма к окружающей среде осуществляются при взаимодействии нервных и гуморальных механизмов регуляции.
Гуморальная регуляция представляет собой способ передачи регулирующей информации к эффекторам через жидкую внутреннюю среду организма с помощью молекул химических веществ, выделяемых клетками или специализированными тканями и органами. Этот вид регуляции жизнедеятельности может обеспечивать как относительно автономный местный обмен информацией об особенностях метаболизма и функции клеток и тканей, так и системный эфферентный канал информационной связи, находящийся в большей или меньшей зависимости от нервных процессов восприятия и переработки информации о состоянии внешней и внутренней среды.
Деление механизмов регуляции жизнедеятельности организма на нервные и гуморальные весьма условно и может использоваться только для аналитических целей как способ изучения. На самом деле, нервные и гуморальные механизмы регуляции неразделимы, так как информация о состоянии внешней и внутренней среды воспринимается почти всегда элементами нервной системы - рецепторами, обрабатывается в нервной системе, где может трансформироваться в сигналы исполнительных устройств либо нервной, либо гуморальной природы.
Управляющим «устройством» является, как правило, нервная система. Однако, сигналы, поступающие по управляющим каналам нервной системы передаются в местах окончания нервных проводников в виде химических молекул-посредников, поступающих в микроокружение клеток, т.е. гуморальным путем. А специализированные для гуморальной регуляции железы внутренней секреции управляются нервной системой.
Таким образом, следует говорить о единой нейро-гуморальной системе регуляции физиологических функций.
Общий план строения нервной системы.
Нервная система человека структурно подразделяется на центральную (ЦНС) и периферическую.
ЦНС состоит из нейронов и клеток нейроглии, периферическая – из отростков нейронов и периферических узлов – ганглиев.
К ЦНС относят спинной и головной мозг, к периферической – 12 пар черепно-мозговых нервов, 31 пару спинномозговых нервов и нервные узлы.
Функционально нервную систему делят на соматическую, регулирующую деятельность скелетных мышц и органов чувств и вегетативную (симпатическую, парасимпатическую), регулирующую деятельность внутренних органов, сосудов и желез.
Нейроглия
Нейроглия (от нейро... и греч. glía - клей), глия - клетки в мозге, своими телами и отростками заполняющие пространства между нервными клетками - нейронами - и мозговыми капиллярами. Каждый нейрон окружен несколькими клетками нейроглии, которая равномерно распределена по всему мозгу и составляет около 40% его объёма.
Число глиальных клеток их в центральной нервной системе млекопитающих около 140 млрд. - мельче нейронов в 3-4 раза и отличаются от них по морфологическим и биохимическим признакам.
С возрастом количество нейронов в ЦНС уменьшается, а клеток нейроглии - увеличивается, т.к. последние, в отличие от нейронов, сохраняют способность к делению.

Рис. 2.
Основные функции нейроглии: создание между кровью и нейронами барьера, необходимого как для защиты нейронов, так и главным образом для регуляции поступления веществ в ЦНС и их выведения в кровь; обеспечение реактивных свойств нервной ткани (образование рубцов после травмы, участие в реакциях воспаления, в образовании опухолей и др.).
Различают астроглию, олигоглию, или олигодендроглию, и эпендиму, которые вместе составляют макроглию, а также микроглию, занимающую особое положение среди клеток нейроглии.
Астроглия (около 60% от общего числа клеток) - звездообразные клетки с многочисленными тонкими отростками, оплетающими нейроны и стенки капилляров (рис.); основной элемент гемато-энцефалического барьера; регулирует водно-солевой обмен нервной ткани.
Олигоглия (около 25-30%) - более мелкие, округлые клетки с короткими отростками. Окружают тела нейронов и нервные проводники - аксоны. Отличаются высоким уровнем белкового и нуклеинового обмена; ответственны за транспорт веществ в нейроны. Участвуют в образовании миелиновых оболочек аксонов.
Эпендима состоит из клеток цилиндрической формы, выстилающих желудочки головного мозга и центральный канал спинного мозга. Играет роль барьера между кровью и спинномозговой жидкостью; выполняет, по-видимому, и секреторную функцию.
Нейроглия (главным образом олигоглия) участвует в происхождении медленной спонтанной биоэлектрической активности, к которой относят a-волны электроэнцефалограммы. Система "нейрон - нейроглия" - единый функционально-метаболический комплекс, отличающийся цикличностью работы, адаптивностью реакций, способностью переключения определённых обменных процессов преимущественно в нейроны или в глии в зависимости от характера и интенсивности физиологических и патологических воздействий на ЦНС.
Структура нейрона, его свойства.
Нейроны являются возбудимыми клетками нервной системы. В отличие от глиальных клеток они способны возбуждаться (генерировать потенциалы действия) и проводить возбуждение. Нейроны высокоспециализированные клетки и в течение жизни не делятся.
В нейроне выделяют тело (сому) и отростки. Сома нейрона имеет ядро и клеточные органоиды. Основной функцией сомы является осуществление метаболизма клетки.

Рис.3. Строение нейрона. 1 — сома (тело) нейрона; 2 — дендрит; 3 — тело Швановской клетки; 4 — миелинизированный аксон; 5 — коллатераль аксона; 6 — терминаль аксона; 7 — аксонный холмик; 8 — синапсы на теле нейрона
Число отростков у нейронов различно, но по строению и выполняемой функции их делят на два типа.
1. Одни — короткие, сильно ветвящиеся отростки, которые называются дендритами (от dendro — дерево, ветвь). Нервная клетка несет на себе от одного до множества дендритов. Основной функцией дендритов является сбор информации от множества других нейронов. Ребенок рождается с ограниченным числом дендритов (межнейронных связей), и увеличение массы мозга, которое происходит на этапах постнатального развития, реализуется за счет увеличения массы дендритов и глиальных элементов.
2. Другим типом отростков нервных клеток являются аксоны. Аксон в нейроне один и представляет собой более или менее длинный отросток, ветвящийся только на дальнем от сомы конце. Эти ветвления аксона называются аксонными терминалами (окончаниями). Место нейрона, от которого начинается аксон, имеет особое функциональное значение и называется аксонным холмиком. Здесь генерируется потенциал действия — специфический электрический ответ возбудившейся нервной клетки. Функцией же аксона является проведение нервного импульса к аксонным терминалям. По ходу аксона могут образовываться его ответвления.
Часть аксонов центральной нервной системы покрывается специальным электроизолирующим веществом — миелином. Миелинизацию аксонов осуществляют клетки глии. В центральной нервной системе эту роль выполняют олигодендроциты, в периферической — Шванновские клетки, являющиеся разновидностью олигодендроцитов. Олигодендроцит оборачивается вокруг аксона, образуя многослойную оболочку. Миелинизации не подвергается область аксонного холмика и терминали аксона. Цитоплазма глиальной клетки выдавливается из межмембранного пространства в процессе «обертывания». Таким образом, миелиновая оболочка аксона состоит из плотно упакованных, перемежающихся липидных и белковых мембранных слоев. Аксон не сплошь покрыт миелином. В миелиновой оболочке существуют регулярные перерывы — перехваты Ранвье. Ширина такого перехвата от 0,5 до 2, 5 мкм. Функция перехватов Ранвье — быстрое скачкообразное распространение потенциалов действия, осуществляющееся без затухания.
В центральной нервной системе аксоны различных нейронов, направляющиеся к одной структуре, образуют упорядоченные пучки — проводящие пути. В подобном проводящем пучке аксоны направляются «параллельным курсом» и часто одна глиальная клетка образует оболочку нескольких аксонов. Поскольку миелин является веществом белого цвета, то проводящие пути нервной системы, состоящие из плотно лежащих миелинизированных аксонов, образуют белое вещество мозга. В сером же веществе мозга локализуются тела клеток, дендриты и немиелинизированные части аксонов.

Рис.4.Строение миелиновой оболочки 1 — связь между телом клетки глии и миелиновой оболочкой; 2 — олигодендроцит; 3 — гребешок; 4 — плазматическая мембрана; 5 — цитоплазма олигодендроцита; 6 — аксон нейрона; 7 — перехват Ранвье; 8 — мезаксон; 9 — петля плазматической мембраны
Конфигурацию отдельного нейрона выявить очень трудно, поскольку они плотно упакованы. Все нейроны принято делить на несколько типов в зависимости от числа и формы, отходящих от их тела отростков. Различают три типа нейронов: униполярные, биполярные и мультиполярные.

Рис. 5. Виды нейронов. а — сенсорные нейроны: 1 — биполярный; 2 — псевдобиполярный; 3 — псевдоуниполярный; б — двигательные нейроны: 4 — пирамидная клетка; 5 — мотонейроны спинного мозга; 6 — нейрон двойного ядра; 7 — нейрон ядра подъязычного нерва; в — симпатические нейроны: 8 — нейрон звездчатого ганглия; 9 — нейрон верхнего шейного ганглия; 10 — нейрон бокового рога спинного мозга; г — парасимпатические нейроны: 11 — нейрон узла мышечного сплетения кишечной стенки; 12 — нейрон дорсального ядра блуждающего нерва; 13 — нейрон ресничного узла
Униполярные клетки. Клетки, от тела которых отходит только один отросток. На самом деле при выходе из сомы этот отросток разделяется на два: аксон и дендрит. Поэтому правильнее называть их псевдоуниполярными нейронами. Для этих клеток характерна определенная локализация. Они принадлежат неспецифическим сенсорным модальностям (болевая, температурная, тактильная, проприоцептивная).
Биполярные клетки — это клетки, которые имеют один аксон и один дендрит. Они характерны для зрительной, слуховой, обонятельной сенсорных систем.
Мультиполярные клетки имеют один аксон и множество дендритов. К такому типу нейронов принадлежит большинство нейронов ЦНС.
Исходя из особенностей формы этих клеток их делят на веретенообразные, корзинчатые, звездчатые, пирамидные. Только в коре головного мозга насчитывается до 60 вариантов форм тел нейронов.
Сведения о форме нейронов, их местоположении и направлении отростков очень важны, поскольку позволяют понять качество и количество связей, приходящих к ним (структура дендритного дерева), и пункты, в которые они посылают свои отростки.
Строение и свойства нервных волокон.
Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. По строению оболочек различают:
- миелиновые
- безмиелиновые нервные волокна.
Отросток нервной клетки в нервном волокне называют осевым цилиндром, или аксоном, так как чаще всего (за исключением чувствительных нервов) в составе нервных волокон находятся именно аксоны.
Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы.
Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. Миелиновый слой оболочки такого волокна содержит значительное количество липидов. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это так называемые узловатые перехваты, или перехваты Ранвье.
Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1—2 м/с, тогда как толстые миелиновые — со скоростью 5—120 м/с.
В безмиелиновом волокне волна деполяризации мембраны идет по всей аксолемме, не прерываясь, а в миелиновом возникает только в области перехватов.
Миелинизация нервных волокон играет важную роль в процессе развития нервной системы. Следы миелина обнаруживаются в нервных волокнах задних и передних корешков уже на 4-м месяце внутриутробной жизни плода. К концу 4-го месяца миелин выявляется в нервных волокнах, составляющих восходящие, или афферентные (чувствительные), системы боковых канатиков, тогда как в волокнах нисходящих, или эфферентных (двигательных), систем миелин обнаруживается на 6-м месяце. Приблизительно в это же время наступает миелинизация нервных волокон задних канатиков. Миелинизация нервных волокон корково-спинномозговых (пирамидных) путей начинается на последнем месяце внутриутробной жизни и продолжается в течение года после рождения. Это свидетельствует о том, что процесс миелинизации нервных волокон распространяется вначале на филогенетически более древние, а затем — на более молодые структуры. От последовательности миелинизации определенных нервных структур зависит очередность формирования их функций. Этим объясняется позднее созревание пирамидной системы и постепенное начало проявления ее функции в первые два года жизни ребенка. В это время бурно развиваются нервные элементы коры большого мозга, где происходит не только миелинизация нервных волокон, но и функциональная дифференциация клеточных элементов и их постепенное созревание, которое длится в течение первого десятилетия.
В постнатальном периоде постепенно происходит окончательное созревание всей нервной системы, в частности ее самого сложного отдела — коры большого мозга, играющей особую роль в мозговых механизмах условно-рефлекторной деятельности, формирующейся с первых дней жизни.
Пучки нервных волокон собраны в нервы. Нервы покрыты оболочкой из соединительной ткани. Собственная оболочка покрывает и каждое волокно в отдельности. Как и нейроны, нервы бывают сенсорными (афферентными) и моторными (эфферентными). Встречаются также смешанные нервы, передающие импульсы в обоих направлениях..
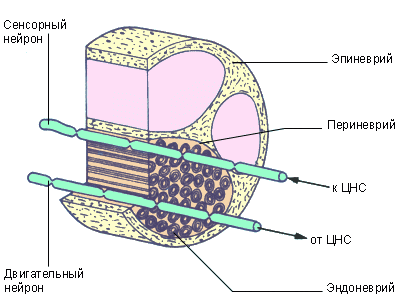
Рис.6. Поперечный срез нервного волокна
Связь между нейронами.
Передача информации от одного нейрона к другому происходит в синапсах. Обычно посредством синапсов связаны между собой аксон одного нейрона и дендриты или тело другого. Синапсами связаны с нейронами также окончания мышечных волокон. Число синапсов очень велико: некоторые клетки головного мозга могут иметь до 10 000 синапсов.

Рис.7 Строение синапса 1 — аксон пресинаптического нейрона; 2 — микротрубочки; 3 — синаптический пузырек (везикула); 4 — синаптическая щель; 5 — дендрит постсинаптического нейрона; 6 — рецептор для медиатора; 7 — постсинаптическая мембрана; 8 — пресинаптическая мембрана; 9 — митохондрия
В образовании синапса участвуют как пресинаптическая часть, так и мембрана последующей клетки (постсинаптическая часть). Синапс состоит из пресинаптической бляшки (расширение терминали аксона), оканчивающейся пресинаптической мембраной, и постсинаптической мембраны (участка мембраны постсинаптической клетки, лежащего под синаптической бляшкой). Между пресинаптической и постсинаптической мембранами расположена синаптическая щель. От ее величины зависит тип передачи информации через синапс. Если расстояние между мембранами нейронов не превышает 2—4 нм или они контактируют между собой, то такой синапс является электрическим, поскольку подобное соединение обеспечивает электрическую связь между этими клетками, позволяющую электрическому потенциалу непосредственно или электротонически передаваться от клетки к клетке. Доля электрических синапсов в ЦНС позвоночных очень мала.
Чаще всего мембраны нейронов расположены в непосредственной близости друг к другу и разделены обычным межклеточным пространством (щелью шириной примерно 20 нм) — смежное соединение. Такая смежность мембран облегчает перемещение из одной клетки в межклеточную щель химических веществ (ионов, метаболитов нейронов), которые оказывают влияние как на ту же самую клетку, так и на отростки соседних нейронов. Эти соединения нейронов относят к химическим синапсам.
В пресинаптическом окончании химического синапса находятся пузырьки, содержащие вещество — передатчик, называемое медиатором. В момент прихода к синаптической бляшке электрического импульса пузырьки открываются в пресинаптическую щель, выбрасывая туда медиатор. Медиатор диффундирует через щель и на постсинаптической мембране взаимодействует с рецептором, специфически чувствительным к медиатору, при этом возникает постсинаптический потенциал.
Таким образом, информация в нервной системе передается только в одном направлении (от пресинаптического нейрона к постсинаптическому) и в этом процессе участвует биологически активное вещество — медиатор.
Лекция 5
Тема: «Возрастная анатомия и физиология нервной системы.
Центральная нервная система.»
План.
ЦНС, ее строение, функции.
Процессы возбуждения и торможения в ЦНС и их взаимодействие.
Центральная нервная система , ее строение, функции.
Спинной мозг лежит в позвоночном канале и представляет собой тяж длиной 41 - 45 см (у взрослого), диаметром около 1 см, вес 30 г, несколько сплющенный спереди назад. Вверху он непосредственно переходит в головной мозг, а внизу заканчивается заострением - мозговым конусом - на уровне II поясничного позвонка. От мозгового конуса вниз отходит терминальная нить, представляющая собой атрофированную нижнюю часть спинного мозга.
Вначале, на 2 месяце внутриутробной жизни, спинной мозг занимает весь позвоночный канал, а затем вследствие более быстрого роста позвоночника отстает в росте и перемещается вверх.
Спинной мозг имеет два утолщения: шейное и поясничное, соответствующие местам выхода из него нервов, идущих к верхней и нижней конечностям. Передней срединной щелью и задней срединной бороздкой спинной мозг делится на две симметричные половины, каждая в свою очередь имеет по две слабовыраженные продольные борозды, из которых выходят передние и задние корешки - спинномозговые нервы. Эти борозды разделяют каждую половину на три продольных тяжа - канатика: передний, боковой и задний. В поясничном отделе корешки идут параллельно концевой нити и образуют пучок, носящий название конского хвоста.
Внутреннее строение спинного мозга. Спинной мозг состоит из серого и белого вещества.
Серое вещество заложено внутри и со всех сторон окружено белым. В каждой из половин спинного мозга оно образует два неправильной формы вертикальных тяжа с передними и задними выступами – столбами (напоминающих бабочку или букву Н), соединенных перемычкой - центральным промежуточным веществом, в середине которого заложен центральный канал, проходящий вдоль спинного мозга и содержащий спинномозговую жидкость. В грудном и верхнем поясничном отделах имеются также боковые выступы серого вещества. Таким образом, в спинном мозге различают три парных столба серого вещества: передний, боковой и задний, которые на поперечном разрезе спинного мозга носят название переднего, бокового и заднего рогов.
Передний рог имеет округлую или четырехугольную форму и содержит клетки, дающие начало передним (двигательным) корешкам спинного мозга.
Задний рог уже и длиннее и включает клетки, к которым подходят чувствительные волокна задних корешков.
Боковой рог образует небольшой треугольной формы выступ, состоящий из клеток, относящихся к вегетативной части нервной системы.
Белое вещество спинного мозга составляет передний, боковой и задний канатики и образовано преимущественно продольно идущими нервными волокнами, объединенными в пучки - проводящие пути. Среди них выделяют три основных вида:
- чувствительные (восходящие) волокна (афферентные, центростремительные), которые частично являются продолжением волокон задних корешков, частично отростками клеток спинного мозга и восходят кверху к головному мозгу.
- двигательные (нисходящие) волокна (эфферентные, центробежные), идущие из головного мозга в спинной на соединение с клетками, дающими начало передним двигательным корешкам;
- вставочные (ассоциативные) волокна, соединяющие участки спинного мозга на различных уровнях;
От спинного мозга, образуясь из передних и задних корешков, отходит 31 пара смешанных спинномозговых нервов: 8 пар шейных, 12 пар грудных, 5 пар поясничных, 5 пар крестцовых и 1 пара копчиковых.
Участок спинного мозга, соответствующий отхождению пары спинномозговых нервов, называют сегментом спинного мозга. В спинном мозге выделяют 31 сегмент.
Спинному мозгу присущи две функции: рефлекторная и проводниковая.
Как рефлекторный центр спинной мозг способен осуществлять сложные двигательные и вегетативные рефлексы. Афферентными - чувствительными - путями он связан с рецепторами, а эфферентными - со скелетной мускулатурой и всеми внутренними органами.
Длинными восходящими и нисходящими путями спинной мозг соединяет двусторонней связью периферию с головным мозгом. Афферентные импульсы по проводящим путям спинного мозга проводятся в головной мозг, неся ему информацию об изменениях во внешней и внутренней среде организма. По нисходящим путям импульсы от головного

Рис.8. Строение спинного мозга.
мозга передаются к эффекторным нейронам спинного мозга и вызывают или регулируют их деятельность.
Нервные центры спинного мозга непосредственно связаны с рецепторами и рабочими органами. Кроме спинного, мозга, такие центры имеются в продолговатом и среднем мозге. Эти центры, непосредственной связи с периферией не имеют. Они управляют ею посредством сегментарных центров. Двигательные нейроны спинного мозга иннервируют все мышцы туловища, конечностей, шеи, а также дыхательные мышцы - диафрагму и межреберные мышцы. Помимо двигательных центров скелетной мускулатуры, в спинном мозге находится ряд симпатических и парасимпатических вегетативных центров. В боковых рогах грудного и верхних сегментах поясничного отделов спинного мозга расположены спинальные центры симпатической нервной системы, иннервирующие сердце, сосуды, потовые железы, пищеварительный тракт, скелетные мышцы, т.е. все органы и ткани организма. Именно здесь лежат нейроны, непосредственно связанные с периферическими симпатическими ганглиями. В верхнем грудном сегменте, находится симпатический центр расширения зрачка, в пяти верхних грудных сегментах - симпатические сердечные центры. В крестцовом отделе спинного мозга заложены парасимпатические центры, иннервирующие органы малого таза
Проводниковая функция спинного мозга. Спинной мозг выполняет проводниковую функцию за счет восходящих и нисходящих путей, проходящих в белом веществе спинного мозга. Эти пути связывают отдельные сегменты спинного мозга друг с другом, а также с головным мозгом.
Головной мозг расположен в черепной коробке и покрыт тремя оболочками. Выстилающие череп и позвоночный канал соединительнотканные оболочки, которые окружают головной и спинной мозг. Наружная оболочка - твердая оболочка мозга - неэластичная, прочная и более толстая, чем средняя (паутинная) и внутренняя (мягкая) оболочки. Между двумя внутренними оболочками циркулирует спинномозговая жидкость.
Мозг взрослого человека в среднем весит 1300-1350гр, новорожденного ребенка – 400 гр.
Головной мозг состоит из :
1. продолговатого мозга;
2. заднего мозга (мозжечок и Варолиев мост);
3.среднего мозга;
4. промежуточного мозга (зрительные бугры и подбугровая область);
5. переднего (конечного)мозга - двух полушарий, соединенных между собой мозолистым телом;
Кратко остановимся на описании анатомии и физиологии перечисленных выше отделов головного мозга.
Продолговатый мозг расположен в полости черепа. Книзу продолговатый мозг переходит непосредственно в спинной мозг. На передней поверхности продолговатого мозга имеется продольная щель, по сторонам которой расположены два возвышения в форме валиков. На задней поверхности проходят продольная борозда и два задних канатика, которые являются продолжением задних столбов спинного мозга.
В своем строении продолговатый мозг сохраняет отдельные сегментарные признаки спинного, однако правильность в распределении серого и белого вещества нарушена. Серое вещество располагается в виде ядер – скопления тел нервных клеток.
От продолговатого мозга отходят с V по XII пары черепно-мозговых нервов. Волокна этих нервов образуют связи между продолговатым мозгом и самыми разнообразными органами человеческого тела. Через черепно-мозговые нервы к нему идут сигналы от рецепторов кожи лица, слизистых оболочек глаз, носовой и ротовой полостей, органа слуха и вестибулярного аппарата, рецепторов гортани, трахеи и легких, от интерорецепторов сердечно-сосудистой системы и пищеварительного тракта.
В сером веществе расположены центры: дыхания, глотания, сердечной деятельности, пищеварения, слюноотделения, защитных рефлексов (кашля, чихания, рвоты, моргания) – рефлекторная функция.
Помимо рефлекторной функции продолговатый мозг выполняет проводниковую функцию. Через него проходят проводящие пути: от спинного мозга – восходящие и к нему – нисходящие.
Таким образом, продолговатый мозг играет огромную роль в жизни организма; малейшее его повреждение представляет большую опасность и часто приводит к смерти, вследствие прекращения дыхания и деятельности сердца и это несмотря на его малые размеры: длина 2,5 – 3 см, вес – 7 г.
Задний мозг (Варолиев мост и мозжечок). Варолиев мост располагается над продолговатым мозгом в виде утолщения. Боковые отделы моста постепенно суживаются и уходят под мозжечок – это ножки моста, они соединяют мост с мозжечком. На передней поверхности Варолиева моста расположены пучки нервных волокон, которые направляются к головному мозгу и переходят в ножки мозга. В глубине Варолиева моста расположены ядра. В продолговатом мозгу и в мосту расположены также парасимпатические ядра, обуславливающие слюноотделение и жизненно важные функции (сердечно-сосудистый и дыхательный центры).
Мозжечок состоит из двух полушарий, соединенных между собой так называемым червем. Мозжечок связан со среднем мозгом, с Варолиевым мостом и продолговатым мозгом. Различают наружное серое вещество мозжечка - его кору и белое вещество, расположенное внутри. Кора мозжечка трехслойная.
Полушария и червь имеют многочисленные, глубокие, концентрические борозды, где располагается большая часть коры (85,5%). В белом веществе каждого полушария заложено по 3 ядра.
Основной функцией мозжечка является координация движений, определяющая их четкость, плавность и сохранение равновесия тела. Повреждение мозжечка у человека и высших животных приводит к расстройству мышечной координации. При отсутствии мозжечковой регуляции мышечные движения становятся неточными и неаккуратными, подобно тем, которые наблюдаются у сильно опьяневшего человека.
Чтобы представить объем работы мозжечка, достаточно сказать, что даже при таком простом движении, как поднесение руки к лицу, чтобы подавить зевок, ему приходится привести в соответствие работу 58 различных мышц, определяющих положение 32 костей.
Мозжечок немедленно вступает в деятельность после того, как из коры или другого участка нервной системы поступит сигнал о ее начале. Импульсы мозжечка последовательно включают и выключают десятки мышц, которые должны участвовать в работе. Мозжечок, получая общие указания о характере движений, как бы вырабатывает детальные инструкции и направляет их ко всем мышцам, участвующим в движении, благодаря чему действие становится четким и мы легко преодолеваем силы инерции.
Кроме того, мозжечок влияет на характер сосудистых рефлексов, на состав крови, на работу желудочно-кишечного тракта и другие вегетативные функции.
Средний мозг (ножки мозга и четверохолмие). Ножки мозга - это скопление нервных волокон, которые имеют вид двух толстых нервных пучков. В ножках мозга выделяют основание и покрышку. В ножках мозга проходят восходящие и нисходящие пути, расположены ядра III и IV пары черепно-мозговых нервов, красные ядра, черная субстанция и другие скопления нервных клеток.
Четверохолмие имеет вид пластинки с четырьмя возвышениями в форме небольших бугорков, два из них верхние и два нижние. Между верхними бугорками расположен эпифиз. В верхних бугорках имеется скопление нервных клеток, которые функционально относятся к подкорковым зрительным центрам; нервные клетки нижних бугорков являются подкорковыми слуховыми центрами. При их участии осуществляются зрительные и слуховые ориентировочные рефлексы.
Наконец, средний мозг имеет важное значение в регуляции мышечного тонуса и в осуществлении установочных рефлексов, обеспечивающих сохранение правильного положения тела в пространстве.
Промежуточный мозг (зрительные бугры и подбугровая область). Зрительные бугры (таламус) – это два больших образования, состоящие из скопления серого вещества образующего около 40 ядер. Функционально их можно разделить на специфические и неспецифические. Специфические ядра получают информацию от рецепторов, перерабатывают её и направляют в определенные области коры головного мозга. Неспецифические ядра не имеют прямой связи с рецепторами, они получают импульсы от рецепторов через большое количество синапсов. Импульсы от этих образований через подкорковые ядра поступают к нейронам, расположенным в различных областях коры головного мозга, вызывая повышение возбудимости этих участков.
Подбугровая область (гипоталамус) расположена к низу от зрительных бугров. Главными образованиями ее является серый бугор и сосочковые тела. В них лежат ядра, имеющие отношение к регулированию обмена веществ в организме. В гипоталамусе находятся высшие вегетативные центры. Он имеет отношение ко сну, температурной и водной регуляции, к углеводному и жировому обмену, к голоду и насыщению, к половой функции. Он имеет также отношение к эмоциональным состояния и сопутствующим им изменениям некоторых функций нашего тела, таким, например, как усиление сердцебиения, появление сухости слизистой рта, потоотделения и т.п.
Ретикулярная формация (от лат. reticulum - сеточка, formatio - образование), сетевидное образование, совокупность нервных структур, расположенных в центральных отделах стволовой части мозга (продолговатом и среднем мозге, промежуточном). Нейроны, составляющие ретикулярную формацию, разнообразны по величине, строению и длине аксонов; их волокна густо переплетаются. Термин "ретикулярная формация", введённый немецким учёным О. Дейтерсом, отражает лишь морфологические её особенности. Ретикулярная формация морфологически и функционально связана со спинным мозгом, мозжечком, лимбической системой и корой больших полушарий головного мозга. В ее области осуществляется взаимодействие поступающих в неё как восходящих - афферентных, так и нисходящих - эфферентных импульсов. Возможна также циркуляция импульсов по замкнутым нейронным цепям. Таким образом, существует постоянный уровень возбуждения нейронов ретикулярной формации, вследствие чего обеспечиваются тонус и определённая степень готовности к деятельности различных отделов центральной нервной системы. Степень возбуждения ретикулярной формации регулируется корой больших полушарий головного мозга.
Нисходящие влияния. В ретикулярной формации различают области, которые оказывают тормозящие и облегчающие влияния на двигательные реакции спинного мозга. Зависимость между раздражением различных областей ствола мозга и спинномозговыми рефлексами впервые отметил в 1862 И. М. Сеченов. В 1944-46 американский нейрофизиолог Х. Мэгоун с сотрудниками показали, что раздражение различных участков ретикулярной формации продолговатого мозга оказывает облегчающее или тормозящее влияние на двигательные реакции спинного мозга. Электрическое раздражение медиальной части ретикулярной формации продолговатого мозга у наркотизированных и децеребрированных кошек и обезьян сопровождается полным прекращением движений, вызываемых как рефлекторно, так и стимуляцией двигательных участков коры мозга.
Раздражение латеральной области ретикулярной формации продолговатого мозга по периферии области, оказывающей тормозящие влияния, сопровождается облегчающим действием на моторную активность спинного мозга. Область ретикулярной формации, оказывающая облегчающие влияния на спинной мозг, не ограничивается продолговатым мозгом, а распространяется кпереди, захватывая область варолиева моста и среднего мозга.
Ретикулярная формация может воздействовать на различные образования спинного мозга, например на альфа-мотонейроны, иннервирующие основные волокна мышц, участвующих в произвольных движениях.
Восходящие влияния. Различные отделы ретикулярной формации (от промежуточного до продолговатого мозга) оказывают возбуждающие влияния на кору мозга, т. е. вовлекают в процесс возбуждения все области коры больших полушарий.
В 1949 итальянский физиолог Дж. Моруцци и Мэгоун, исследуя биоэлектрическую активность мозга, установили, что раздражение ретикулярной формации ствола мозга изменяет медленные синхронные высоковольтные колебания, характерные для сна, на низкоамплитудную высокочастотную активность, характерную для бодрствования. Изменение электрической активности коры мозга сопровождается у животных внешними проявлениями пробуждения.
Ретикулярная формация тесно связана анатомически с классическими проводящими путями, и возбуждение её осуществляется с помощью экстеро- и интероцептивных афферентных (чувствительных) систем. На этом основании ряд авторов относит ее к неспецифической афферентной системе мозга.
Однако применение различных фармакологических веществ при изучении функции ретикулярной формации, открытие избирательного действия химических препаратов на реакции, осуществляемые с ее участием, позволили П. К. Анохину сформулировать положение о специфичности восходящих влияний ретикулярной формации на кору мозга. Активирующие влияния всегда имеют определённое биологическое значение и характеризуются избирательной чувствительностью к различным фармакологическим веществам (Анохин, 1959, 1968). Введённые в организм наркотические средства вызывают торможение нейронов ретикулярной формации, блокируя тем самым её восходящие активирующие влияния на кору мозга.
Кора больших полушарий, испытывающая тонические активирующие влияния со стороны ретикулярной формации, может активно изменять функциональное состояние ретикулярных образований (изменять скорость проведения возбуждения в ней, влиять на функционирование отдельных нейронов), т. е. контролировать, по выражению И. П. Павлова, "слепую силу" подкорки.
Открытие свойств ретикулярной формации, её взаимоотношений с другими подкорковыми структурами и областями коры мозга позволило уточнить нейрофизиологические механизмы боли, сна, бодрствования, активного внимания, формирования целостных условно-рефлекторных реакций, развития различных мотивационных и эмоциональных состояний организма.
Подкорковые ядра (базальные) расположены над промежуточным мозгом, во внутренней части больших полушарий. Представляют собой высший отдел мозгового ствола и объединяют деятельность нижележащих образований, регулируя мышечный тонус и обеспечивая должное положение тела во время физической работы.
Наибольшее значение имеет хвостатое ядро, скорлупа и бледное ядро.
Бледное ядро обеспечивает вспомогательные движения во время двигательных актов и оказывает тормозящее влияние на красное ядро среднего мозга.
При гиперфункции бледного ядра чрезмерно усиливаются вспомогательные движения и резко снижается тонус мышц. При этом наблюдаются размашистые неточные движения, гримасы на лице и т.п. Такие нарушения наблюдаются при таком заболевании как хорея.
При гипофункции бледного ядра исчезают вспомогательные движения, так как снимаются тормозящие влияния на красное ядро. При этом наблюдается маскообразное лицо, движения становятся скованными и негармоничными из-за отсутствия вспомогательных движений. Подобные нарушения наблюдаются при Паркинсоновой болезни.
Хвостатое ядро и скорлупа оказывают тормозящее влияние на бледное ядро. При гиперфункции хвостатого ядра и скорлупы будет заторможено красное ядро и, следовательно, расторможено бледное ядро. В результате будут нарушения типа Паркинсоновой болезни. При гипофункции хвостатого ядра и скорлупы бледное ядро выйдет из-под тормозящего влияния и будет сильно тормозить красное ядро. В результате будут нарушения, как при хорее.
Большие полушария переднего мозга являются самой большой по величине, самой сложной по строению и самой важной по функции частью головного мозга. Так, 80% массы головного мозга приходится на большие полушария. В них содержится около 15 млрд. нейронов, т.е. столько же, сколько во всех остальных отделах ЦНС вместе взятых.
Большие полушария головного мозга состоят из серого и белого вещества. Серое вещество полушарий расположено снаружи в виде тонкого слоя и называется корой головного мозга. Толщина его примерно 2 – 4 мм. Основную массу полушарий составляет белое вещество, расположенное под корой. В глубине его имеются скопления серого вещества в форме отдельных узлов.
Белое вещество морфофункционально подразделяется на три группы волокон. Первая группа — ассоциативные волокна, связывающие участки коры в пределах одного полушария. Вторая группа — комиссуральные волокна, соединяющие симметричные участки полушарий. Наиболее представительная часть комиссуральных волокон — мозолистое тело, основные части которого хорошо видны на сагиттальном разрезе. Третья группа — это проекционные волокна — связывающие кору с нижележащими отделами и наоборот (проекция в кору чувствительных и двигательных центров). Ближе к коре эти волокна представлены 'лучистым венцом; концентрируясь между таламусом и базальными ядрами, они образуют внутреннюю капсулу.
Наряду с перечисленными образованиями в толще полушарий выделяют структуры, которые состоят из серого и белого веществ и относятся к обонятельному мозгу (свод, прозрачная перегородка, гиппокамп и др.).
На поверхности мозговой коры хорошо выражены многочисленные складки, которые отделяются друг от друга то более глубокими, то менее глубокими бороздами. Участок коры, расположенный между двумя бороздами, называется извилиной. Общая поверхность больших полушарий равна 1700 – 18—см2 , но 2/3 ее находится в глубине борозд (около 1200 см2 ).
Полушария головного мозга разделяются на следующие доли: лобные, теменные, височные и затылочные. Расположение долей примерно соответствует костям черепа. Границей между долями являются наиболее постоянные и выраженные борозды.
На выпуклой верхнелатеральной поверхности полушарий хорошо видны две постоянные глубокие борозды, которые можно использовать в качестве ориентиров при отграничении долей. Это идущая во фронтальной плоскости центральная(Роландова) борозда, разграничивающая лобную и теменную доли, и идущая под углом к ней боковая (Сильвиева), или латеральная борозда, отделяющая названные доли от височной. Затылочная доля с выпуклой стороны отграничена от соседних нечетко, с медиальной поверхности граница представлена постоянной, хорошо видной теменно-затылочной бороздой. Наконец, в глубине боковой борозды можно видеть пятую долю — островок.
В лобной доле предцентральная борозда, идущая параллельно центральной борозде, отделяет вместе с последней предцентральную извилину, где замыкаются условные двигательные рефлексы (ядро двигательного анализатора). От V слоя коры этой извилины (гигантские пирамидные клетки) начинается пирамидный путь, связывающий правую двигательную область коры с левой половиной тела и наоборот.
Две борозды, идущие перпендикулярно предцентральной, отделяют верхнюю, среднюю и нижнюю лобные извилины. В задних отделах средней лобной извилины расположены центр письма (двигательный анализатор письменной речи) и центр сочетанного поворота головы и глаз в одну сторону. В задних отделах нижней лобной извилины локализован моторный центр речи (двигательный анализатор артикуляции речи). Этот центр, как и другие речевые центры (слуховой анализатор устной речи и пр.), имея двустороннюю закладку в эмбриогенезе, развивается лишь с одной стороны: у правшей слева, а у левшей справа.
В теменной доле постцентральная борозда отделяет вместе с центральной бороздой постцентральную извилину — центр осязания, болевой и температурной чувствительности. Близкий к указанному центру по локализации и функции центр стереогнозии (узнавание предметов на ощупь) расположен в верхней теменной дольке, отграниченной от нижней теменной дольки внутритеменной бороздой. Последняя следует перпендикулярно к постцентральной борозде. В пределах нижней теменной дольки видна надкраевая извилина, в которую упирается латеральная борозда (ориентир извилины). Это центр праксии (синтез целенаправленных навыков трудового, спортивного характера и пр.) Ниже надкраевой видна угловая извилина — центр чтения (зрительный анализатор письменной речи). Оба последних центра расположены у правшей слева.
Височная доля имеет пять извилин, идущих параллельно латеральной борозде и друг другу. Три извилины, разделенные верхней и нижней височными бороздами, видны на верхнелатеральной поверхности, а две — на нижней и медиальной поверхностях полушарий. В задней части верхней височной извилины (у правшей слева) находится слуховой анализатор речи (сенсорный центр речи). В среднем отделе верхней височной извилины, на поверхности, обращенной к островку, расположено ядро слухового анализатора. Наиболее медиально в височной доле находится парагиппокампальная извилина (рядом с ней в толще височной доли залегает гиппокамп). Передний отдел извилины изогнут и поэтому назван крючком. Здесь находится центр обоняния и вкуса.
На медиальной поверхности затылочной доли четко выделяется глубокая шпорная борозда, выше и ниже которой находятся соответственно клин и язычная извилина, являющиеся центром зрительного анализатора.
Ряд структур головного мозга: поясная извилина (над мозолистым телом), парагиппокампальная извилина, гиппокамп, свод и др., расположенные в совокупности в виде кольца, объединены в функциональное понятие «лимбическая система» (лимб — кольцо). Эта система регулирует работу внутренностей, эндокринных органов, а также обеспечивает эмоциональные реакции.
Полостью конечного мозга являются боковые желудочки. Они имеют по четыре отдела, расположенные в каждой доле полушария: передний рог (в лобной доле), задний рог (в затылочной доле), нижний рог (в височной доле), центральная часть (в теменной доле). Боковые желудочки сообщаются с III желудочком через межжелудочковое отверстие.
Кора большого мозга имеет толщину 1,3—4,4 мм и площадь 2200 см2, Клетки коры расположены в 6 слоев, отличающихся друг от друга строением и функцией: I и II слои (считая снаружи) —ассоциативные, III и IV — афферентные и V и VI — эфферентные. В функциональном отношении, по И. П. Павлову, кора представляет собой центральные отделы различных анализаторов, обеспечивающих анализ и синтез поступающих в нервную систему через органы чувств раздражений.
Нейроноархитектоника коры
Как уже отмечалось, кора большого мозга подразделяется в эволюционном отношении на древнюю кору, paleocortex, старую кору, archicortex, и новую кору, neocortex. Древняя и старая кора располагаются на медиальной и базальной поверхности полушария, составляя 2,8% всей поверхности коры. Их окружают межуточные корковые формации, выделенные Н. И. Филимоновым под названием перипалеокортекса и периархикортскса (мезопалеокортекс и мезоархикортекс по Международной гистологической номенклатуре). На долю последних приходится 1,6 % поверхности полушария. Остальные 95,6% поверхности занимает новая кора.
На основании данных эмбрионального развития выделяют гомогенетическую кору, или изокортекс, который на 6-м месяце внутриутробного периода подразделяется на 6 слоев, и гетерогенетическую кору, или аллокортекс, имеющий меньшее количество слоев. В последующие периоды развития количество слоев в коре может изменяться. Кора, сохраняющая шестислойное строение, называется гомотипической; если количество слоев увеличивается или уменьшается, говорят о гетеротипической коре.
Рассмотрим строение гомогенетической, гомотипической шестислойной коры. От поверхности в глубину она имеет следующие слои:
Молекулярный слой, который состоит в основном из нервных волокон, содержит немногочисленные горизонтальные нейроциты.
Наружный зернистый слой, образованный густо расположенными зернистыми нейроцитами с короткими, сильно ветвящимися отростками; в нем имеются также малые пирамидальные нейроны.
Пирамидный слой, характеризующийся наличием пирамидальных нейроцитов с длинными, уходящими в глубину аксонами. Размеры клеток увеличиваются от поверхности в глубину слоя.
Внутренний зернистый слой, содержащий малые зернистые и звездчатые нейроциты.
Ганглионарный слой, образованный пирамидальными пейроцитами самых больших размеров; их аксоны выходят из пределов коры.
Полиморфный слой, имеющий в своем составе нервные клетки различной формы; наиболее типичными являются треугольные и веретенообразные нейроциты.
Гетеротиаическая кора отличается от гомотипической тем, что внутренний зернистый или ганглиозный слои в ней слабо выражены или отсутствуют. С другой стороны, пирамидный, внутренний зернистый или ганглионарный слои могут подразделяться на подслои.
Функциональное значение слоев коры определяется их клеточным составом и нейропальными связями. В молекулярном слое оканчиваются волокна из других частей коры и противоположного полушария. Здесь происходит переключение импульсов на верхушечные дендриты пирамидальных нейронов, поднимающиеся в молекулярный слой из глубоких частей коры (рис. 19).
Нейроциты наружного зернистого и пирамидного слоев являются в основном ассоциативными; за счет их отростков осуществляются горизонтальные и вертикальные внутрикоковые связи. Эти два слоя представляют наиболее позднее эволюционное приобретение и особенно сильно развиты в коре человека. В онтогенезе они созревают позже других слоев. Внутренний зернистый слой представляет главный воспринимающий слой коры. В нем оканчивается большинство специфических проекционных афферентных волокон из таламуса и коленчатых тел. Часть афферентных волокон образует синапсы с базальными дендритами пирамидальных нейронов 3-го слоя.
Ганглионарный слой является местом начала эфферентных проекционных путей коры.
Полиморфный слой функционально неоднороден. Имеются данные о том, что его нейроциты дают начало комиссуральным волокнам. Нейроны этого слоя образуют также вертикальные связи с вышерасположенными слоями коры.
Благодаря сильному развитию внутрикорковых связей нейроны коры большого мозга образуют замкнутые крути различной степени сложности, расположенные как параллельно, так и перпендикулярно к поверхности коры. Каждый из этих кругов имеет собственные афферентные и эфферентные волокна и представляет отдельную функциональную единицу (И. С. Беритов).
Нейроноархитектоника коры неодинакова в различных ее участках. Бродман разделил кору на 52 архитектонических поля, каждое из которых обозначается определенной цифрой. Нумерация полей коры, предложенная Бродманом, сохранена и в цитоархитектоничеокой карте Института мозга, однако здесь ряд полей делится на подполя, которые обозначаются буквами латинского алфавита.
Миелоархитектоника коры
В коре выделяют радиальные и тангенциальные нервные волокна. Первые вступают в кору из белого вещества или, наоборот, идут из коры в белое вещество полушария. Вторые располагаются параллельно поверхности коры и образуют в ней сплетения, называемые полосками. Различают полоски молекулярного слоя, наружного и внутреннего зернистого слоев и ганглионарного слоя. Количество полосок в различных частях коры неодинаково; в зависимости от этого описываются однополосковый, двухполосковый и многополосковый типы коры. Детальное исследование миелоархитектоники коры провели немецкие исследователи супруги Сесиль и Оскар Фогты, которые разделили полушарие более чем на 100 миелоархитектонических полей.
Представительство функций в коре большого мозга
В центральной нервной системе с особенной полнотой проявляется диалектическое единство строения и функции. В свое время И. П. Павлов выдвигал в отношении нервной системы принцип структурности, согласно которому все мельчайшие детали конструкции мозга имеют динамическое значение. Исходя из этого, мы должны признать, что особенности архитектоники коры, различия в структуре областей и полей коры связаны с их функциональными отправлениями. Так мы подходим к вопросу о локализации, или представительстве, функций в коре большого мозга.
Учение о функциях мозга имеет многовековую историю. Значение мозга как органа ощущения и мысли понимали еще древнегреческие врачи Алкмеон и Гиппократ. В течение длительного времени свойство мышления приписывали «психической пневме» и локализовали психические функции в желудочках мозга. Леонардо да Винчи на одном из своих анатомических рисунков изобразил связь желудочков с opганом зрения посредством зрительного нерва, который рассматривался в то время как полая трубка, передающая зрительные впечатления в «передний» желудочек. Отсюда, как считал Леонардо да Винчи, ощущения передаются «общему чувству», расположенному «точно в середине головы». В XVII веке французский философ и ученый Р. Декарт считал, что в эпифизе мозга находится «седалище души», основываясь на том, что эпифиз лежит в центре полости черепа.
Первая попытка локализовать психические функции в определенных частях мозга;; была предпринята австрийским врачом Ф. И, Галлем, создавшим в конце XVIII века ученее -френологию. Согласно этому учению у человека имеется 37 душевных способностей, каждая из которых связана с особым участком полушария большого мозга. Величина и, форма полушарий отражают развитие тех или иных способ; костей л, .в свою очередь, отражаются на форме черепа. По: этому на основании осмотра и измерения головы можно судить о характере и способностях человека. Публичные выступления Галля носили материалистический и атеистический характер и были запрещены австрийскими властями, что вынудило ученого покинуть Вену. Учение Галля было вульгарно-материалистическим, и вскоре стала очевидной несостоятельность многих его положений, однако в нем содержалась правильная мысль о взаимосвязи между строением и функцией мозга.
Начало строго научному подходу к вопросу о локализации функций в коре положил французский ученый П. Брока, открывший в 1861 году двигательный центр речи в заднем отделе нижней лобной извилины левого полушария. В. А. Бец показал, что открытые им гигантские пирамидальные нейроны имеют отношение к двигательным центрам коры. В дальнейшем была определена локализация в коре различных видов чувствительности, выделены двигательные и ассоциативные зоны и стала преобладать тенденция к строгому пространственному делению коры в функциональном отношении.
Наряду с этим некоторые исследователи придерживались взгляда о функциональной равноценности, эквипотенциальности всех частей полушарий. Такой взгляд выдвигали в XIX веке французский физиолог Флуранс, производивший опыты на голубях, в XX веке американский психолог Лешли, ставивший эксперименты на крысах.
Как отмечает А. Р. Лурия, каждое из этих двух направлений внесло вклад в развитие учения о функциях мозга. Локализационистские взгляды способствовали разработке тонкой анатомии мозга и представлений о дифференцированной работе его частей. Сторонники эквипотенциализма развивали положения о высокой пластичности мозговой ткани и способности к восстановлению функций, нарушенных в результате ее повреждения.
Большую роль в изучении локализации функций в коре большого мозга сыграли работы И. П. Павлова и сто школы. И. П. Павловым была заложена основа динамического подхода к локализации функций. Согласно учению И. П. Павлова нервная система представляет комплекс анализаторов внешней и внутренней среды организма. В коре большого мозга находятся центральные концы анализаторов, которые осуществляют анализ и синтез сигналов, поступающих к ним по афферентным нервным путям.. На основании экспериментов на собаках И. П. Павлов установил, что корковые концы анализаторов не имеют резко очерченных границ и что их периферические части заходят друг за друга, образуя обширные зоны перекрытия.
За последние десятилетия в изучении функций коры широко применялись электрофизиологические методики, а также наблюдения на больных людях при нейрохирургических операциях. В настоящее время признается, что различные • корковые поля являются функционально многозначными, то есть участвуют в нескольких функциях, выступая в каждой из них в сочетаниях с другими мозговыми структурами. Поэтому любая функция коры обычно является результатом деятельности ряда взаимосвязанных ее» частей. Архитектоника коры составляет анатомическую основу понимания ее сложной, системной деятельности.
В структурно-функциональном отношении кора большого мозга делится на передний и задний отделы. Граница между ними проходит по центральной борозде. Таким образом, задний отдел полушария образован теменной, затылочной и височной долями, а передний отдел представлен лобной долей. Задний отдел коры связан с восприятием и переработкой афферентных сигналов, то есть представляет комплекс корковых концов анализаторов. Передний отдел коры имеет отношение к организации действий.
Принято подразделять цитоархитектонические поля коры на первичные, вторичные и третичные (рис. 1).

Рис. 9. Локализация первичных и вторичных корковых полей. Первичные чувствительные поля заштрихованы, первичное двигательное поле черное. Вторичные чувствительные поля выделены мелким, вторичные двигательные поля---крупным пунктиром. Третичные поля коры оставлены белыми. Нумерация корковых полей по Бродману.
Первичные поля представляют четко локализованные участки, которые соответствуют центральным (ядерным) частям анализаторов. В эти поля по специфическим афферентным путям приходит основная масса сигналов от органов чувств. В архитектоническом отношении первичные поля характеризуются сильным развитием внутреннего зернистого слоя, в котором, как мы знаем, оканчивается большинство афферентных волокон. Первичные поля связаны с реле-ядрами таламуса и коленчатых тел. Они имеют экранную структуру и, как правило, пространственную проекцию периферии. Повреждение первичных воспринимающих нолей приводит к нарушению непосредственного восприятия и топкой дифференцировки раздражений.
Представительство кожной и сознательной проприоцептивной чувствительности занимает первичные корковые поля 1, 2 и 3, расположенные в постцентральной извилине. В каждом полушарии имеется обратная проекция противоположной половины тела. В верхней части извилины находится проекция нижней конечности, ниже располагается проекция живота, груди, верхней конечности. В нижней трети извилины локализуются раздражения, приходящие от головы и шеи, причем проекция частей головы и шеи является уже прямой. Первичное зрительное поле (17) расположено на медиальной поверхности полушария вдоль шпорной борозды; над бороздой проецируются верхние, а под бороздой нижние квадранты сетчаток. В первичном зрительном поле хорошо развит внутренний зернистый слой коры, который подразделяется на 3 части посредством волокнистых белых полосок (area striata).
Первичные слуховые поля (41 и 42) локализуются в поперечных височных извилинах и заходят в верхнюю височную извилину.
Первичная обонятельная область захватывает древнюю кору, покрывающую обонятельный треугольник, переднее продырявленное пространство, прозрачную перегородку и подмозолистое поле.
Первичное вкусовое поле располагается, по-видимому, в нижнем участке постцентральной извилины, в глубине латеральной борозды. По другим данным, оно находится в крючке.
Точно не установлено нахождение поля, принимающего раздражения от вестибулярного аппарата.
Вторичные поля коры примыкают к первичным, их можно рассматривать как периферические части корковых концов анализаторов. Вторичные поля связаны с ассоциативными ядрами таламуса, в них преобладают нейроны с короткими отростками, за счет которых происходит взаимодействие сигналов, поступающих по различным афферентным системам и из первичных полей. При поражении вторичных полей сохраняются элементарные ощущения, однако нарушается способность к более сложным восприятиям.
Вторичные поля не имеют строго очерченных границ, и в них отсутствует пространственная проекция периферии. Вторичное поле общей чувствительности располагается в верхней теменной дольке (поля 5 и 7). Вторичные зрительные поля занимают медиальную и большую часть латеральной по-верхности затылочной доли (поля 18 и 19). Вторичное слуховое поле 22 находится в верхней и средней височных извилинах. Вторичные обонятельные и вкусовые поля локализуются в парагиппокампальной извилине и крючке (поля 28 и 34).
Представительство в коре внутренностей и сосудов изучено недостаточно, по-видимому оно имеет более диффузный характер, чем представительство покрова тела и мускулатуры. Важная роль в регуляции функций внутренних органов придается лимбической области коры, в которую входят поясная и парагиппокампальная извилины, гиппокамп, перегородка и подмозолистое поле. Лимбическая кора вместе с миндалевидным телом и некоторыми другими подкорковыми образованиями составляют лимбическую систему, которая представляет субстрат эмоций и реакций, связанных я основными биологическими, влечениями (голод, жажда,страх,- ярость, половое возбуждение).
Третичные поля коры отличаются, наиболее тонкой нейронной структурой с преобладанием ассоциативных элементов: Третичные поля занимают в заднем отделе полушария всю нижнюю и часть верхней теменной долек и затылочно-височно-тменную область. Они связаны с задним ядром (подушкой) таламуса. В третичных полях осуществляются наиболее сложные формы взаимодействия анализаторов, лежащие в основе познавательного процесса (гнозия), формируются программы целенаправленных действий (праксия).
Кора височной доли имеет отношение к хранению и воспроизведению впечатлений. Известный канадский учёный; нейрохирург Уилдер Пенфилд установил, что при электрическом раздражении некоторых участков височной коры возникают своеобразные реакции в форме «вспышек пережитого» или ощущения «уже виденного». При этом переживается как действительность какой-то отрезок прошлого или больному кажется, что операция уже производилась раньше и он знает, что собирается делать хирург. Пенфилд предполагает, что в коре височных долей создается нейронная запись потока сознания, она хранится неопределенно долгое время, но человек не может произвольно воспроизводить эту «запись», и, она активируется лишь при искусственной симуляции или болезненных состояниях.
Передний отдел полушария также подразделится на: первичные, вторичные и третичные корковые поля. Первичное двигательное поле располагается в предцентральной извилине (поле 4). Здесь отсутствует внутренний зернистый слой (агранулярная кора) и особенно сильно развит ганглионарный слой, в котором находятся гигантские пирамидальные нейроны. Аксоны этих клеток образуют пирамидный путь. На гигантские пирамидальные нейроны непосредственно переключаются импсульсы, приходящие сюда по афферентным волокнам из мозжечка через центральное медиальное ядро таламуса. В первичном двигательном поле представлены все мышечные группы тела. Как и в первичном чувствительном поле, здесь имеется обратная проекция частей тела. Верхние участки предцентральной извилины связаны с мускулатурой нижних конечностей, далее книзу находится проекция мускулатуры туловища, верхней конечности/головы и шеи. Первичное двигательное поле регулирует индивидуальные сокращения мышц, от которых зависит выполнение произвольных движений.
Вторичным двигательным полем является кора лобных извилин кпереди от предцентральной извилины (поля 6 и 8). Они характеризуются сильным развитием пирамидного слоя, в котором преобладают крупные пирамидальные нейроциты. Они, как и гигантские клетки первичного двигательного поля, получают импульсы из мозжечка. Вторичное двигательное поле является корковым центром экстрапирамидной системы; эфферентные волокна идут из него в полосатое тело.
Сигналы, посылаемые вторичным двигательным полем, необходимы для осуществления сложных стереотипных двигательных актов.
Поля 4 и 6 имеют многочисленные связи с задним отделом полушария. Роль двигательной коры состоит в том, что в ней сопоставляются программы движений с реальным ходом выполнения этих программ и вносятся коррекции по ходу выполняемых действий. Обратная связь между двигательным аппаратом и корой осуществляется через мозжечок, который воспринимает афферентные импульсы с периферии и после соответствующей переработки сигналов направляет их в кору.
Третичные поля занимают большую часть поверхности лобной доли и четверть всей коры большого мозга. В них хорошо выражен внутренний зернистый слой, в который приходят импульсы из медиальных ядер таламуса. Третичные поля лобной коры имеют отношение к высшим формам целенаправленной деятельности и социальному поведению человека. При их поражении не возникает каких-либо чувствительных или двигательных расстройств, но человек становится пассивным, безучастным к происходящему, теряет способность к оценке событий и предвидению.
Важнейшую особенность человека составляет членораздельная речь. И. П. Павлов относил речь ко второй сигнальной системе, с помощью которой осуществляется непрямое отражение действительности. Речевые функции имеют широкое представительство в коре большого мозга. И. П. Павлов полагал, что двигательная речь должна быть связана с двигательной областью коры, письменная речь — со зрительными областями, а слышимая речь — с височными долями. У. Пенфилд и Л. Роберте, основываясь на данных, полученных при электрическом раздражении и удалении у больных различных участков коры, выделили в коре три речевых поля (рис. 2).
Заднее речевое поле располагается в затылочно-височно-теменной области, захватывая все три височные, надкраевую и угловую извилины. Это поле связано преимущественно с восприятием и пониманием речи и в функциональном отношении является ведущим. При его поражении всегда наступает афазия — расстройство речи.
Переднее речевое поле находится в задней части нижней лобной извилины и соответствует двигательному центру речи Брока.
Верхнее, дополнительное речевое поле локализуется у верхнего края полушария и заходит на его медиальную поверхность кпереди от парацентральной дольки. Поражение этого участка не всегда вызывает речевые расстройства.
Речевые поля, как и другие части коры, связаны с ядрами таламуса. Заднее поле связано с задним ядром, верхнее поле— с латеральным ядром, переднее поле — с медиальными ядрами таламуса.

Рис. 10. Речевые поля коры по Пенфилду и Робертсу.
Все речевые поля объединяются посредством нервных связей, образуя единую функциональную систему.
Замечательной особенностью речевых центров коры является их асимметрия. У большинства людей они располагаются в левом полушарии, которое является доминантным г. отношении речи. Принято считать, что доминантность левого полушария связана с праворукостью и что у левшей речевые функции локализуются в правом полушарии. В противоположном субдоминантном полушарии соответствующие участки коры выполняют иные функции: с ними связано представление о пространственных отношениях и схеме тела.
Нервные пути, относящиеся к коре большого мозга.
Все нервные пути подразделяются на три группы: ассоциативные, комиссуральные и проекционные
Ассоциативные нервные пути соединяют части коры в одном полушарии большого мозга. К ним относятся дугообразные волокна, связывающие соседние извилины и поля коры, и ряд длинных путей, которые имеют преимущественно продольное направление. Среди них выделяют верхний и нижний продольные пучки, крючкообразный пучок и пояс.
Комиссуральные нервные пути осуществляют связь между обеими полушариями. Они проходят в белом веществе полушарий в основном поперечно. Большинство комиссуральных волокон идет в мозолистом теле, которое соединяет области попой коры. Древняя и старая кора соединяются посредством путей, составляющих переднюю спайку; передняя ее часть связывает первичную обонятельную кору двух полушарий, задний участок содержит волокна, связывающие кору височных долей. Спайка свода соединяет гиппокампальные формации правого и левого полушарий.
Проекционные нервные пути связывают кору с нижерасположенными этажами мозга. Они пронизывают полушария в вертикальном направлении. Большинство проекционных путей проходит через внутреннюю капсулу.
Различают короткие и длинные проекционные системы.
Короткие проекционные пути:
1. Лучистый венец образуют таламо-корковые и корково-таламические пучки, соединяющие большинство полей новой коры с ядрами таламуса.
2. Зрительная лучистость состоит из волокон зрительного пути, идущих от ядер латерального коленчатого тела в кору затылочной доли;
Слуховая лучистость представляет систему волокон, проходящих от ядер медиального коленчатого тела в кору височной доли.
Корково-стриальные волокна идут от двигательных корковых полей к ядрам полосатого тела.
Корково-гипоталамические пучки соединяют кору лобной и височной долей и островка с ядрами гипоталамуса.
6. Свод содержит волокна, которые соединяют гиппокамп с ядрами сосцевидных тел.
Длинные проекционные пути:
Пирамидные пути состоят из корково-спинномозговых и корково-ядерных волокон.
Корково-красноядерный путь соединяет двигательную зону коры с красным ядром.
Корково-мостовые волокна идут от лобной, теменной, височной и затылочной долей к ядрам моста; соответственно различают лобно-мостовой, теменно-мостовой, височно-мостовой и затылочно-мостовой пути.
Корково-ретикулярные волокна проходят от коры лобной доли в ретикулярную формацию мозгового ствола.
Внутрикорковые и внекорковые связи обеспечивают структурно-функциональную целостность коры и ее взаимодействие со всеми другими отделами центральной нервной системы, что представляет важнейшее условие для осуществления сложных и многообразных функций.
Установлено, что почти вся кора больших полушарий человека состоит из шести слоев. Каждый слой имеет свои особенности строения. Эти слои следующие:
1. молекулурный (поверхностный);
2. наружный зернистый;
3. пирамидный;
4. внутренний зернистый;
5. ганглиозный;
6. полиморфный.
В различных участках строение коры обладает особенностями, что проявляется главным образом различной степенью выраженности того или иного слоя, количеством и густотой клеток. Одинаковые по клеточному строению участки стали обозначать одинаковыми цифрами, а разные – разными. Наибольшее распространение получила классификация по Бродману. По этой классификации в коре больших полушарий различают 52 основных поля. Так, передняя центральная извилина обозначается как поле 4. Впереди нее находятся поля 6, 8, 9,10,11. В задней ценральной извиле находятся поля 1,2,3. Оказалось, что различные поля имеют не только различное клеточное строение, но нейроны этих полей выполняют и различные функции.
В коре больших полушарий принято выделять ряд отдельных зон, выполняющих определенную функцию. Это сенсорные, ассоциативные, моторные зоны.
Сенсорные зоны представляют собой те участки коры, в которых приходят нервные импульсы от рецепторов. Выделяют следующие сенсорные зоны:
В затылочной зоне (поле 17) – зрительная сенсорная зона. В эту зону приходят импульсы от фоторецепторов сетчатки глаза. При его повреждении наступает полная слепота.
В височной доле (поле 41) находится слуховая сенсорная зона, или корковый центр слуха. В эту зону приходят нервные импульсы от рецепторов кортиева органа. При его повреждении наступает полная глухота.
Ассоциативные зоны представляют собой те участки коры, в которые приходят нервные импульсы сразу от нескольких видов рецепторов (кожных, зрительных, слуховых и др.). В ассоциативных зонах происходит слияние, синтез нервных импульсов от многих качественно отличных рецепторов. Ассоциативные зоны находятся рядом с сенсорными, иногда окружая их кольцеобразно. Выделяют следующие ассоциативные зоны:
Зрительная (поля 18, 19) – расположена кольцеобразно вокруг зрительной сенсорной зоны. При повреждении 18 и 19 полей зрение сохраняется, но человек не может узнать предметы (поле 18) или не может ориентироваться в новой обстановке, фиксируя взгляд на предметах (поле 19).
Слуховая ассоциативная зона (поля 22, 42) расположена около слуховой сенсорной зоны (поле 41). При повреждении 22-го поля наступает музыкальная глухота, т.е. человек не может узнать ранее хорошо знакомую ему мелодию, отличить одну ноту от другой. При повреждении 42-го поля наступает словесная глухота, т.е. человек слышит слова, но не может их узнать.
Все ассоциативные зоны связаны между собой длинными и короткими волокнами, и следовательно, функционируют как единая система, в которой протекает анализ и синтез информации, поступающей от всех рецепторов нашего организма.
Моторная или двигательная зона (поле 4) локализована в передней центральной извилине. В верхней части передней центральной извитлины локализованы нейроны, иннервирующие мышцы нижних конечностей, в средней части – мышцы туловища, а в нижней части – мышцы верхних конечностей, шеи и головы. При повреждении той или другой части передней центральной извилины наступает центральный паралич соответствующих мышц. Из этой зоны начинается пирамидный путь, обеспечивающий наши произвольные движения.