

3.2.3. Строение и состав липопротеинов
Поверхностная часть липопротеинов образована монослоем ориентированных фосфолипидов и белками (аполипопротеинами) (рис. 14).
Фосфолипиды
Амфифильные
аполипопротеины
Триацилглицеролы
Холестерин
Эфиры
холестерина
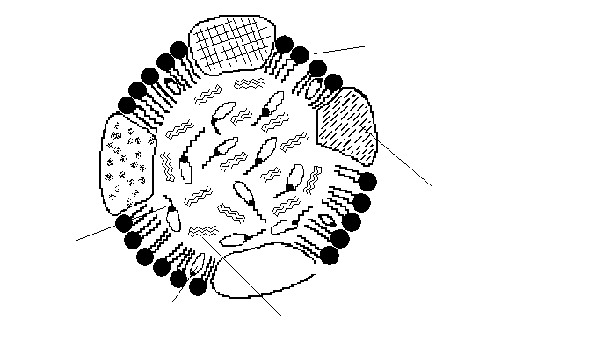
Рис. 14. Строение липопротеинов
Фосфолипиды гидрофильной частью молекулы образуют наружную поверхность, а гидрофобные концы «растворены» в липидной фазе внутри частицы. В составе липопротеинов обнаружено несколько разных белков. Для всех этих белков характерно также наличие гирофильной и гидрофобной частей. Гидрофильные радикалы аминокислот апобелка контактируют с плазмой крови (с молекулами воды), а гидрофобные радикалы – с липидами внутри липопротеиновой мицеллы.
Таблица 12
Представители аполипопротеинов
(обозначаются латинскими буквами А, В, С)
ЛПВП и хиломикроны |
Имеют два главных апобелка – А-I и А-II. А-I в составе ЛПВП является активатором фермента лецитин:холестеролацилтрансферазы |
ЛПНП и ЛПОНП |
Апобелок В-100. Синтезируется в печени |
Хиломикроны |
Апобелок В-48. Синтезируется в кишечнике (и возможно в печени) |
ЛПОНП и ЛПВП |
Апобелки С-I и C-II являются активатором липопротеинлипазы и, следовательно, участвуют в освобождении крови от триацилглицеролов |
ЛПОНП и ЛПВП |
Апобелок Е (богатый аргинином). В избытке имеется у людей, страдающих гиперлипопротеинемией типа III |
Гидрофобное ядро липопротеина содержит главным образом триацилглицерины, свободный холестерол и эфиры холестерола, а также жирорастворимые витамины и возможно присутствие чужеродных веществ гидрофобной природы.
Таблица 13.
Состав липопротеинов крови человека (%)
Фракция ЛП |
Белки |
Триацил-глицеролы |
Фосфо-липиды |
Холестерин |
|
Эфиры |
Свободный |
||||
ЛПВП |
45 |
5 |
25 |
20 |
5 |
ЛПНП |
25 |
7 |
21 |
40 |
7 |
ЛПОНП |
10 |
50 |
18 |
15 |
7 |
Хиломикроны |
2 |
85 |
7 |
4 |
2 |
Плотность и электрофоретическая подвижность липопротеинов прямо пропорциональны содержанию белков и обратно пропорциональны содержанию триацилглицеринов.
Примечание к таблице 13: приведенные величины имеют лишь ориентировочный характер, поскольку в процессе функционирования липопротеинов (обмен компонентами между фракциями ЛП в крови, а также между ЛП и тканями) их состав непрерывно изменяется.
3.2.4. Образование и функции липопротеинов
Липопротеины образуются в клетках слизистой оболочки кишечника (хиломикроны), в клетках печени – гепатоцитах (ЛПОНП и ЛПВП), в плазме крови (ЛПНП и ЛПВП) – при функционировании ЛП.
Жиры (фосфолипиды, триацилглицерины, холестерол), синтезированные в клетке эпителия кишечника или печени, проникают в шероховатый эндоплазматический ретикулум, образуя каплю. Фосфолипидный монослой замыкается вокруг капли, присоединяются апобелки, и формируется отдельная липопротеиновая частица. Затем липопротеины перемещаются в аппарат Гольджи, где образуются секреторные гранулы. Освобождение липопротеинов в кровь или лимфу происходит путем экзоцитоза. В крови происходит дозревание липопротеинов, то есть обмен поверхностными компонентами (апобелками) между разными фракциями липопротеинов крови и стенками кровеносных сосудов органов и тканей.
Выделенные из крови хиломикроны и ЛПОНП содержат апобелки С и Е, в то же время секретируемые (новообразованные) липопротеины содержат незначительное количество этих апобелков или же вообще их не содержат. По-видимому, апобелки С и Е из ЛПВП переносятся на хиломикроны и ЛПОНП после того как они поступают в кровоток.
Хиломикроны и ЛПОНП служат для транспорта триацилглицеринов по кровеносному руслу, а ЛПВП и ЛПНП – для транспорта холестерина.
Жиры, ресинтезирующиеся в клетках кишечника из продуктов переваривания, включаются главным образом в хиломикроны. Хиломикроны поступают в лимфатические капилляры кишечника, затем через лимфатические сосуды брыжейки в грудной проток и оттуда через яремную вену в общий кровоток.
Жиры, образующиеся в печени, упаковываются в ЛПОНП, которые поступают в кровь. Печень выделяет в кровь 20–50 г жиров в сутки (в составе ЛПОНП).
При пищеварении содержание липопротеинов в крови повышается, и кровь приобретает белесоватый цвет. Максимум концентрации липопротеинов наблюдается через 4–5 часов после приема пищи.
Липопротеинлипаза и ее значение в утилизации триацилглицеролов тканями
Хиломикроны и ЛПОНП распределяют по органам за сутки 70–150 г экзогенных и эндогенных жиров. В эндотелии кровеносных сосудов разных органов есть фермент липопротеинлипаза гидролизующий жиры из состава липопротеинов. Липопротеинлипаза локализована на стенках капилляров, к которым данный фермент прикрепляется протеогликановыми цепями гепаратансульфата. Липопротеинлипаза обнаружена в экстрактах из сердца, жировой ткани, селезенки, легких, мозгового слоя почек, аорты, диафрагмы и лактирующей молочной железы. Липопротеинлипаза практически отсутствует в свободном виде в составе крови, однако, после инъекции гепарина, гепарансульфатная связь перестает удерживать липопротеинлипазу и фермент поступает в кровяное русло, где катализирует гидролиз триацилглицеролов.
Как фосфолипиды, так и апобелок С-II находящиеся в составе хиломикронов и ЛПОНП, являются одновременно кофакторами липопротеинлипазы.
Апобелок С-II имеет специфический участок связывания фосфолипидов, которым он присоединяется к липопротеину.
Таким образом, хиломикроны и ЛПОНП обеспечивают липопротеинлипазу как субстратом (триацилглицеролом), так и кофакторами.
При переходе от сытого состояния к голодному концентрация триацилглицеролов в плазме крови уменьшается; при этом липопротеинлипаза сердца остается насыщенной субстратом, а насыщенность фермента жировой ткани снижается. Это приводит к перераспределению потребления субстрата (триацилглицерола) в пользу ткани сердца. Подобное перераспределение наблюдается также при лактации (при образовании молока в молочных железах).
Изменение состава липопротеиновых фракций крови при участии липопротеинлипазы. Пути удаления ЛП из крови
Хиломикроны и ЛПОНП под действием липопротеинлипазы теряют около 90% триацилглицеролов, а также апобелок С возвращается на ЛПВП (апобелок Е остается). Диаметр остатка хиломикрона уменьшается в 2 раза, а содержание в них холестерола и эфиров холестерола увеличивается (за счет убыли триацилглицеролов). ЛПОНП подвергаются подобным превращениям и образуются остатки ЛП средней плотности.
Липопротеинлипаза имеет центр связывания липопротеинов и каталитический центр для гидролиза триацилглицеролов. Продукты гидролиза поступают в клетки тканей, где могут окисляться или участвовать в других метаболических превращениях.
Хиломикроны и ЛПОНП, постепенно освобождаясь от триацилглицеролов, превращаются в ЛПНП, а также, вероятно, в ЛПВП. Время полужизни хиломикронов и ЛПОНП в крови около 5 часов.
ЛПНП и ЛПВП поглощаются путем эндоцитоза клетками печени, кишечника, жировой ткани, почек, надпочечников и разрушаются в лизосомах.
Деградация ЛПНП до 50 % осуществляется в печени и 50% во внепеченочных тканях. На поверхности этих клеток есть рецепторы (В-100) для связывания ЛПНП и ЛПВП2. Если рецепторы не функционируют, то развивается семейная гиперхолестеролемия. Существует прямая корреляция между вероятностью возникновения коронарного атеросклероза и концентрацией ЛПНП в крови.
Транспорт холестерина в крови (молоке). Пути удаления холестерина из кровотока и организма
Особая роль гепатоцитов заключается в том, что в них синтезируется холестерин не только для собственных нужд, но и «на экспорт».
Липопротеины содержат холестерин и эфиры холестерина. Свободный холестерин входит в состав поверхностного фосфолипидного монослоя между гидрофобными концами жирных кислот. Эфиры холестерина находятся в липидном ядре липопротеина.
Между циркулирующими в крови липопротеинами происходит обмен холестерином, особенно активный между ЛПНП и ЛПВП. При контактах липопротеиновых частиц холестерин диффундирует из одной частицы в другую. Обмен имеет двусторонний характер, но в целом преобладает поток холестерина в ЛПВП. Это связано с тем, что ЛПВП активно этерифицируют холестерин с участием лецитин–холестерол–ацилтрансферазы (ЛХАТ).
ЛХАТ катализирует перенос ацильного остатка жирной кислоты из β–положения лецитина на холестерин.
О
Лецитин
Лизолецитин
Холестерин
Олеилохолестерин
Образующиеся в поверхностном слое ЛПВП эфиры холестерина погружаются внутрь частицы. Вследствие этого концентрация свободного холестерина в поверхностном слое уменьшается и освобождается место для поступления холестерина из других липопротеинов. Двусторонний обмен холестерином путем диффузии происходит также при контакте липопротеинов с клетками.
ЛПВП извлекают холестерин из клеточных мембран, а ЛПНП, наоборот, снабжают клетки холестерином.
ЛПВП, нагруженные холестерином, удаляются из кровотока путем эндоцитоза клетками печени, а ЛПНП поглощаются клетками многих органов. ЛПВП предотвращают накопление избытка холестерина в клетках.
В стационарном состоянии суммарное количество холестерина, поступающего в кишечник с пищей, и холестерина, синтезируемого в тканях, равно количеству экскретируемых холестерина и желчных кислот в кишечник в составе желчи.