

8. Ооплазматическая сегрегация при дроблении.
После проникновения сперматозоида начинаются процессы расслоения, отмешивания различных составных частей цитоплазмы яйцеклетки (ооплазмы) – ооплазматическая сегрегация.
Намечается радиальная (относительно анимально-вегетативной оси) симметрия.
У многих беспозвоночных разные бластомеры дают начало строго определенным зачаткам.
Видимо, это обусловлено ооплазматической сегрегацией при дроблении и взаимодействиями между бластомерами.
Движения цитоплазмы хорошо заметны в тех случаях, когда разные участки цитоплазмы содержат разноцветные гранулы (желток, темный пигмент и др.). Например, у гребневиков по периферии яйцеклетки располагается зеленая цитоплазма (до стадии 8 бластомеров). После деления становятся неравномерными: крупные бластомеры отпочковывают от себя микромеры, которые целиком состоят из зеленой эктоплазмы. Микро- и макромеры резко отличаются по своей будущей судьбе и не способны ее изменить.
При спиральном дроблении (например, кольчатые черви и моллюски) на вегетативном полюсе есть полярная плазма. После 3 деления она вся попадает в один бластомер (1D, вегетативное полушарие, спинная сторона). Позже она большей частью попадает в 2d и 4d, из которых развивается большая часть эктодермы и целомическая мезодерма. Те, при удалении полярной плазмы, возникают личинки, лишенные мезодермы и некоторых эктодермальных закладок.

9. Дробление. Типы и закономерности пространственного расположения бластомеров.
Дробление начинается после оплодотворения и активации. Дробление – ряд непрерывно следующих друг за другом митотических делений, но рост отсутствует. Образуется полость внутри – бластоцель.
Положение частей цитоплазмы не меняется, но они попадают в разные бластомеры. Ядерно-плазменное отношение в начале дробления достигает уровня, характерного для обычных соматических клеток.
Дробление (синхронность и асинхронность указывают на перестройку клеточных циклов и отражают синтетическую активность бластомеров):
- синхронное (ранние стадии, первые 4-7 раз, число бластомеров 2^n)
- асинхронное (сначала идет волнами; пробуждается транскрипционный аппарат зародыша, появляется фаза G1; иногда эту фазу называют бластуляцией)
Функции дробления:
- формирование многоклеточности
- увеличение ядерно-плазменного отношения (которое сильно уменьшилось во время большого роста)
Особенности клеточного цикла:
- синтез ДНК начинается уже в пронуклеусах (все факторы инициации синтеза и репликации (полимеразы, гистоны и др) уже есть в яйце с оогенеза), начинается еще в телофазах.
- Клеточные циклы укорочены за счет G1 и G2 периодов (опять же потому, что все нужные продукты есть в яйце).
- S период укорочен за счет полирепликонности клеток эукариот – каждый репликон может реплицироваться автономно (большая полирепликонность, чем у обычных эукариотических клеток).
Цитотомия - осуществляется с помощью двух механизмов:
- образование кольца из сократимых микрофиламентов (образуется в кортикальном слое клетки, в плоскости, перпендикулярной прямой, соединяющей клеточные центры; начинает деление цитоплазмы; ярче выражен у маложелтковых клеток)
- встраивание запасенных предшественников цитоплазмы (сильнее у многожелтковых клеток)
Регуляция дробления. Фактор созревания (пик концентрации в митозе, самая низкая – в S-фазе), который регулируется цитостатическим фактором (стабилизирует фактор созревания, задерживая клетки в митозе), а тот ингибируется ионами Са.
Пространственная организация дробления. Определяется:
- закономерным расположением интерфазных ядер
- закономерной ориентацией веретен деления
- движениями бластомеров
Закон Гертвига-Сакса:
- клеточное ядро стремится расположиться в центре чистой, свободной от желтка цитоплазмы.
- веретено клеточного деления стремится расположиться по направлению наибольшего протяжения свободной от желтка цитоплазмы.
Принято считать, что характер дробления зависит от типа яйцеклетки.
Дробление:
1. Полное (голобластическое)
1.1. Равномерное (для олиго- и изолецитальных клеток). Объемы бластомеров одинаковы. Целобластула – однослойная бластодерма с крупным бластоцелем в центре.
1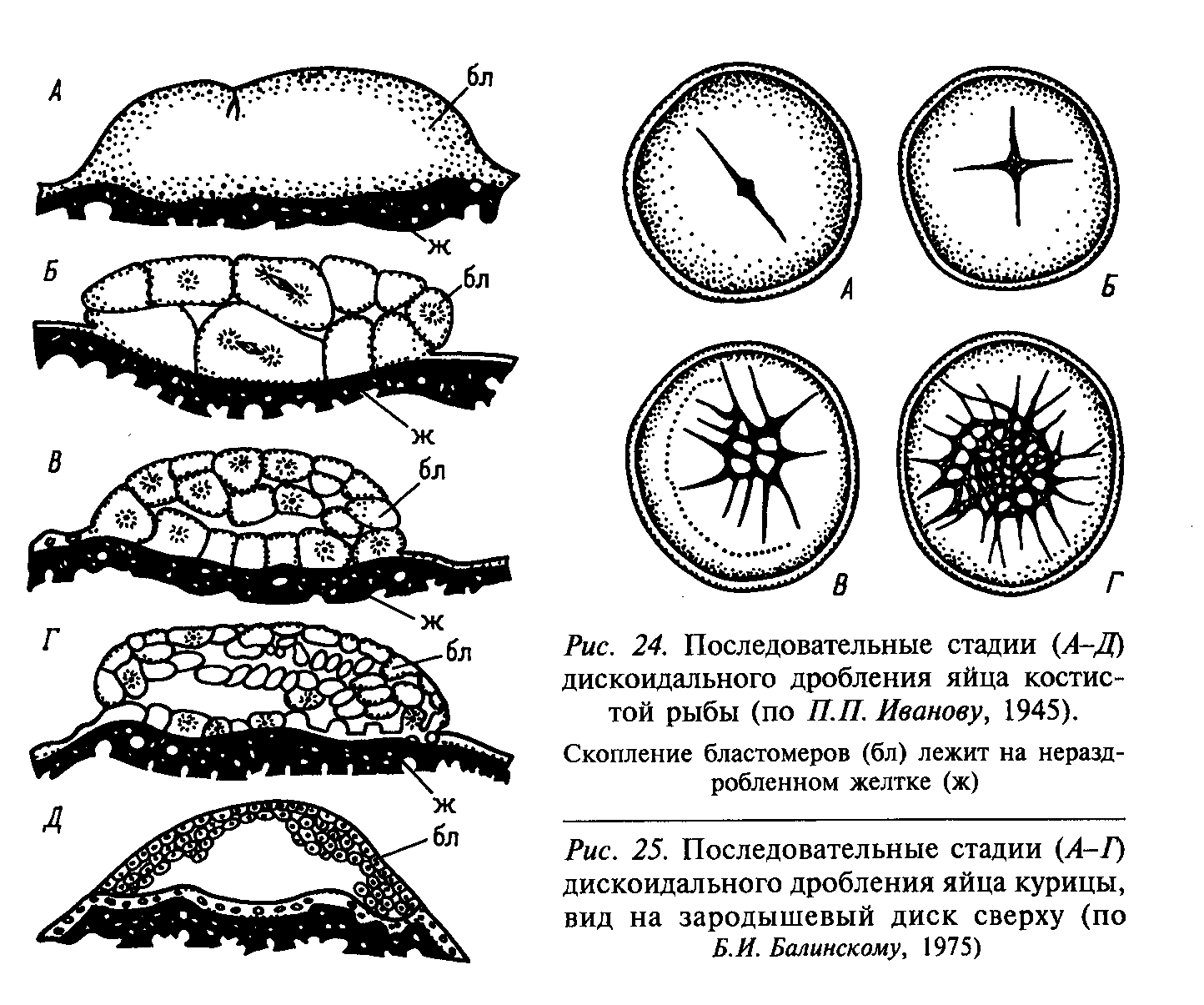 .2. Неравномерное
(для мезо- и тело). Объемы бластомеров
неодинаковы. Амфибластула – состоит
из микро- и макромеров, бластоцель
сдвинут к анимальному полюсу (к
микромерам).
.2. Неравномерное
(для мезо- и тело). Объемы бластомеров
неодинаковы. Амфибластула – состоит
из микро- и макромеров, бластоцель
сдвинут к анимальному полюсу (к
микромерам).
2. Неполное (меробластическое)
2.1. Дискоидальное (поли- и тело-). Борозды формируются вслед за делениями ядер, но разделяют только один из полюсов яйца. Дискобластула – диск бластомеров, лежащий на желтке.
2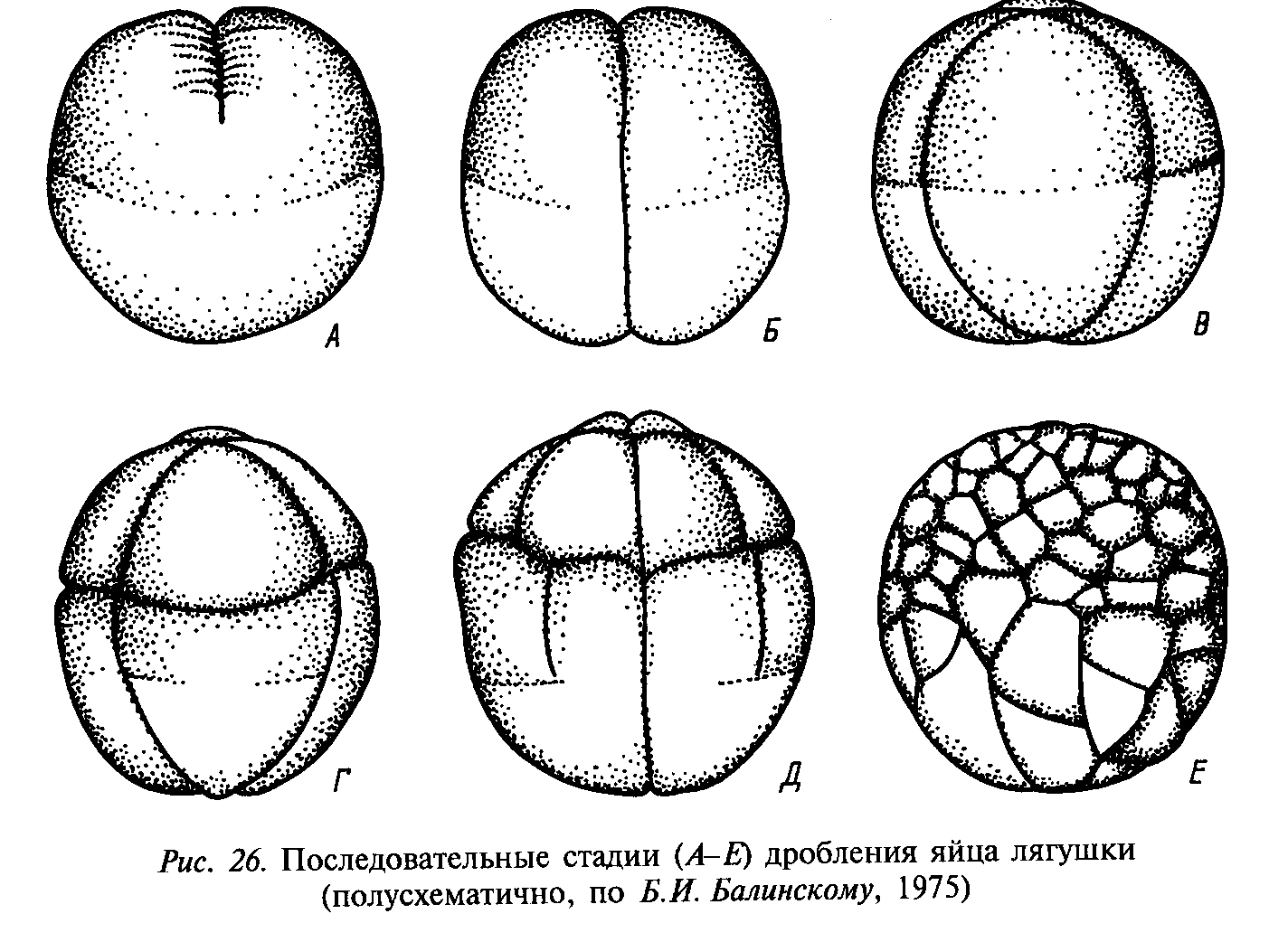 .2. Поверхностное
(поли- и центро-). Поверхностный слой
цитоплазмы разделяется перегородками,
перпендикулярными поверхности яйца,
центральная часть не делится. Перибластула
– не имеет бластоцеля – внутри желток.
.2. Поверхностное
(поли- и центро-). Поверхностный слой
цитоплазмы разделяется перегородками,
перпендикулярными поверхности яйца,
центральная часть не делится. Перибластула
– не имеет бластоцеля – внутри желток.
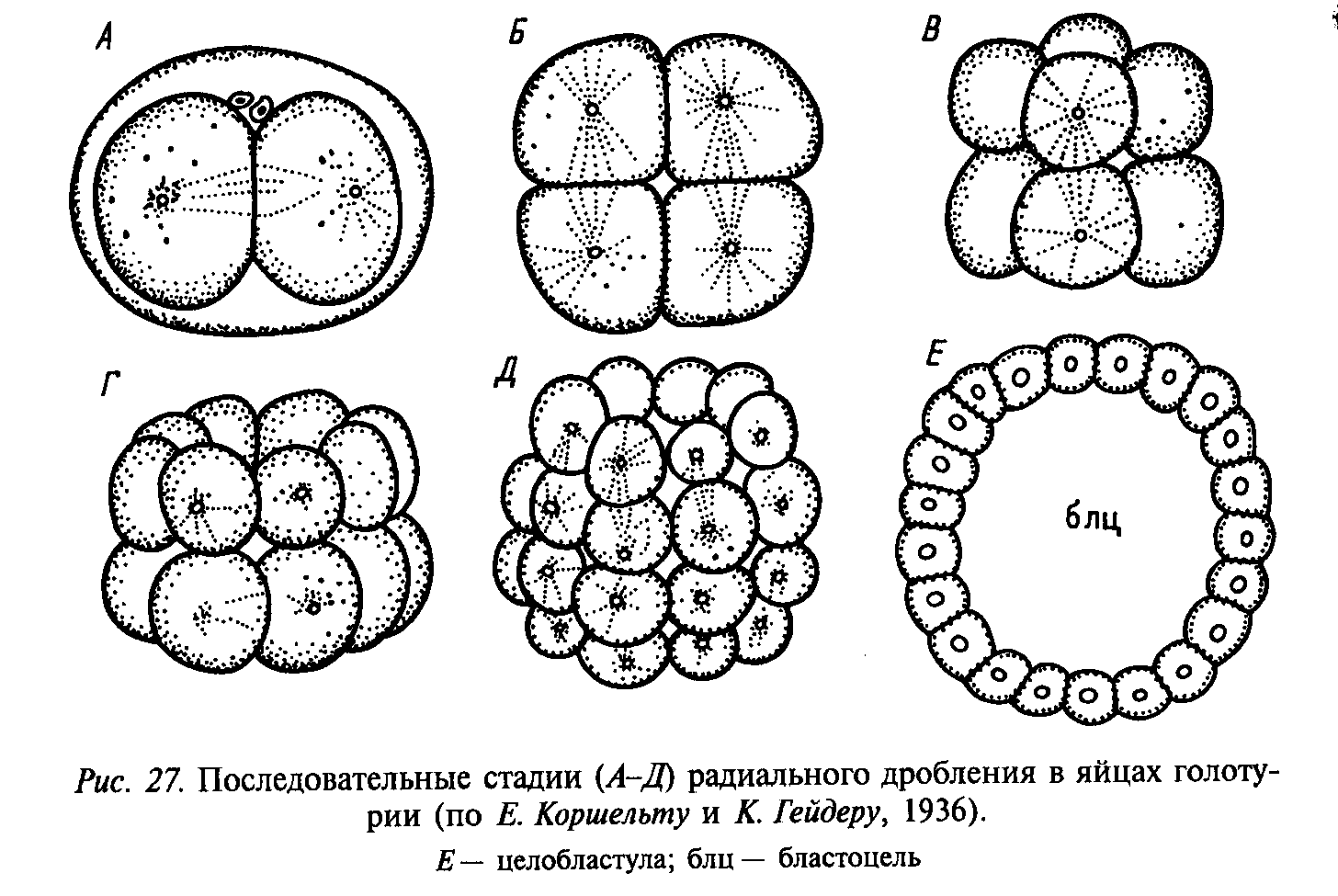
Расположение образующихся из зиготы бластомеров в пространстве друг относительно друга послужило основанием для определения типов дробления, исходя из расположения бластомеров. Выделяют радиальное (ланцетник), спиральное (моллюски), билатеральное, или двусторонне-симметричное (круглые черви), бисимметричное, или двусимметричное (гребневики), и анархичное (плоские черви) дробления.
При радиальном дроблении борозды деления (митотические веретена) ориентированы параллельно или перпендикулярно анимально-вегетативной оси яйцеклетки. Через такую бластулу проходит несколько плоскостей (осей) симметрии.
Спиральное дробление отличается нарушением такого соответствия (борозды деления располагаются наклонно к анимально-вегетативной оси), и дочерние бластомеры располагаются как бы по спирали. Образующаяся при спиральном дроблении бластула (стереобластула) не имеет ни полости, ни даже одной плоскости симметрии.
Билатеральное дробление характеризуется наличием в формирующейся бластуле одной оси (плоскости) симметрии. При бисимметричном дроблении формирующаяся бластула имеет две оси (плоскости) симметрии.
Анархичное деление резко выделяется от описанных выше неупорядоченным расположением бластомеров и отсутствием оси (плоскости) симметрии.