35. Эмбриональные регуляции различных групп животных. Закон Дриша и его современные трактовки.
Эмбриональные регуляции – восстановление нормальной, геометрически правильной и полной структуры организма, несмотря на удаление, добавление или перемешивание части материала зародыша.
Открыты были при отделении друг от друга двух бластмеров морского ежа (морская вода без Са, который укрепляет межклеточные контакты). Разделенные бластомеры дробились абсолютно так же, давая полубластулы, которые замыкались в шар. Таким образом, некоторая часть зародыша может дать целостный организм (это и есть эмбриональная регуляция = дришевская регуляция).
Дришевские регуляции возможны лишь при наличии по крайней мере мультипотентных клеток зародыша. Могут рассматриваться как достаточный экспериментальный критерий мультипотентности.
Проспективное значение каждого элемента системы есть функция его положения в целом.
Опыты: слияние двух яйцеклеток и получение нормальной особи; сдавливание дробящейся яйцеклетки морского ежа, так что перемешивались бластомеры, но возникали совершенно нормальные зародыши, так как судьба бластомеров менялась в соответствии с их новым положением. Были поставлены опыты, в которых бластомеры были диссоциированы и перемешаны. В большом числе случаев формировался полноценный организм, но развитие шло различными путями (эквифинальность), например, через формирование морулы.
Интерпретации закона:
Позиционная информация (Wolpert) – на определенной стадии развития, еще до видимой дифференцировки, каждая клетка по отдельности получает информацию о своем положении в зародыше, а за тем с помощью генетического аппарата интерпретирует эту информацию, дифференцируясь независимо от окружения. ПИ может задаваться неким веществом морфогеном или соотношением концентрации нескольких морфогенов. Они распределены неоднородно, в простейшем случае – по градиенту. Источник морфогена и сток (нидняя точка градиента).Много недостатков у такой концепции.
Морфогенетическое поле – система согласованного взаимодействия всех элементов зародыша, обеспечивающая целостный порядок. Части зародыша ощущают свое положение в зависимости от окружения, но на них могу влиять не только непосредственные соседи, но и более дальние (собственно, поэтому «поле»). Особых точек может и не быть, но могут быть градиенты поля. Теория впервые сформирована А.Г. Гурвичем. МП может быть связано с целым набором физ-хим факторов, механическим напряжением
Мозаичность развития (mosaic development) — тип индивидуального развития зародыша, при котором форма и расположение его отдельных частей детерминированы еще до оплодотворения; наличие М.р. подтверждается тем, что после удаления некоторой части эмбриона на ранней стадии развития эта «потеря» в дальнейшем не восполняется. Недришевские регуляции. При рассыпании зародышей амфибий на отдельные клетки, те вновь собирались в экто- энто- и мезодерму.
36. Роль внеклеточного матрикса и механических напряжений в регуляции клеточной дифференцировки.
Внешние факторы дифференцировки можно разделить на химические и механические. Из механических факторов наиболее хорошо изучен фактор натяжения.
Химические факторы:
- связывание лиганда, пришедшего извне, с рецептором клетки: дистантные взаимодействия (когда лиганд пришел с расстояния, многократно превышающего поперечник клетки), контактные (когда лиганд произведен соседней клеткой) и короткодистантные (когда лиганд продуцировала клетка, расстояние до которой сопостовимо с несколькими поперечниками клетки).
Короткодистантные взаимодкйствия особенно важны т.к. именно на них основаны действия ньюкуповской индукции (индукция мезодермы на стадии средней-поздней бластулы) и шпемановской индукции (индукция нейральных закладок из эмбриональной эктодермы хордомезодермой).
При контактных взаимодействиях лиганд может быть встроен в мембрану своей клетки или перемещаться вдоль нее. Это обоюдные взаимодействия. Кроме того, лиганд может быть иммобилизован на компонентах внеклеточного матрикса, что, например, играет важную роль в дифференцировке клеток мезодермального происхождения.
Механическое напряжение: механическое напряжение играет важную роль в ориентации морфогенетических клеточных движений. Кроме того, в дальнейшем развитии механическое напряжение играет важную роль и в дифференцировке клеток. Например, дифференцировка клеток глазного зачатка определяется растяжением-сжатием: если эпителий растянут, то клетки дифференцируются в пигментный эпителий (при развитии эта сторона становится более выпуклой), а если растяжения нет, то образовывается сетчатка. При нормальном развитии это поддерживается силами внутриглазного давления. В экспериментах удавалось получать из зачатка сплошной пигментный эпителий (при распластывании ткани) и сплошную сетчатку (при условиях отсутствия растяжения). При проколе глазной камеры уже дифференцированный пигментный эпителий преобразовывался в сетчатку из-за отсутствия натяжения.
Механическое напряжения изменяет биохимию клетки - влияет на скорость синтеза нуклеиновых кислот и белков. Некоторые типы клеток дифференцируются только в условиях натяжения. Например, фибробласты, остеобласты, хондроциты не дифференцируются в суспензиии, но дифференцируются только в посеве на растянутый субстрат. А эритроидные клетки синтезируют глобиновую мРНК и гемоглобин только в суспензии, но никак не растянутыми. Клеткам эндотелия для активации генов и нормального синтеза белков необходимо пульсирующее натяжение.
У дрозофилы по крайней мере один ген, экспрессирующийся в области эмбриональной глотки, активируется давлением клеток задних отделов кишечной трубки. Если этого давления нет, ген не активируется. Если создать это давление искусственно, он активируется.
Механическое давление способствует связыванию в клеточных ядрах белка бета-катенина, который регулирует ряд ключевых процессов развития. Например, определяет дорсальную сторону зародыша амфибии и активирует гены, кодирующие эмбриональные индукции.
Ряд авторов полагает, что ткани зародышей и взрослых буквально пронизаны сетью фибрилл, находящихся в напряженном состоянии. В межклеточных пространствах эта сеть состоит из элементов внеклеточного матрикса (в основном, коллагена различных типов и фибронектина, который особо важен в данрой роли у зародышей).
Напряжение, возикающие во внеклеточном матриксе передаются через белки интегрины на особые участки клеточной мембраны, преимущественно фокальные контакты. Около них сосредоточены важные ферментные комплексы и элементы цитоскелета промежуточные и микрофиламенты). Они запускают разнообразные внутриклеточные сигнальные пути, где в качестве вторичного посредника нередко выступает Са2+.
Фибриллярные структуры непрерывны вплоть до ядра, где элементы цитоскелета связываются с волокнами ядерного матрикса, а те, в свою очередь, связаны с ДНК интерфазных хромосом. При короткой интерфазе хромосомы сохраняют т.н. ориентацию Рабля (центромерами к противоположным стенкам дочерних клеток), которую они приобрели в телофазе митоза. А если интерфаза длинная, то ориентация Рабля нарушается, хотя хромосомы прикреплены к ядерной оболочке в определенных местах. Характер и места прикрепления хроматина на разных стадиях развития эмбриона может отличаться.
Существуют теории о том, что в межклеточном веществе сигналы передаются по твердым субстратам, сделанных из определенных белковых молекул. А из этого делается вывод о наличии волн конформационных перестроек, прокатывающихся по белковым конструкциям при передаче сигнала.
МЕЗОДЕРМА (все вместе)
Мезодерма
Закладка мезодермы
Типы закладки мезодермы:
- телобластический (наиболее часто у спирально дробящихся форм). Две крупные клетки потомки бластомера 4d расположены в полости бластоцеля в области губ, имеют полярную плазму и дают начало всей целомической мезодерме личинки. Эти бластомеры называются мезобластами или мезотелиобластами. В результате отделения от них более мелких клеток возникает пара мезодермальных полосок, которые позже подразделяются на сомиты, внутри которых впоследствии образуются целомические полости (шизоцельный/кавитационный способ). Мезодерма никак не связана с энтодермой.
- энтероцельный
(у ланцетника, иглокожих). Материал
будущей мезодермы вворачивается вместе
с энтодермой в составе гастрального
впячивания (первичный кишечник или
архентерон). Граница между закладками
экто- и мезодермы неразличима. Мезодерма
выделяется путем выпячивания стенок
архентерона и их отшнуровывания или,
реже, деламинации или иммиграции клеток
из них. После отеделения мезодермы
остается чисто энтодермальный материал,
архентерон превращается в полость
вторичной (дефинитивной) кишки.
энтероцельный
(у ланцетника, иглокожих). Материал
будущей мезодермы вворачивается вместе
с энтодермой в составе гастрального
впячивания (первичный кишечник или
архентерон). Граница между закладками
экто- и мезодермы неразличима. Мезодерма
выделяется путем выпячивания стенок
архентерона и их отшнуровывания или,
реже, деламинации или иммиграции клеток
из них. После отеделения мезодермы
остается чисто энтодермальный материал,
архентерон превращается в полость
вторичной (дефинитивной) кишки.
Производные мезодермы
Осевая мезодерма и боковая пластинка.
Осевая мезодерма подразделяется на сомиты и метамеризуется, причем закладка сомитов в разных классах хордовых неодинаков. У ланцетника закладываются жнетероцельно и сразу имеют целомическую полость, в то время как у большинства позвоночных сперва закладывается в виде сплошных скоплений мезодермальных клеток, и лишь потом появляются полости шизоцельным путем.
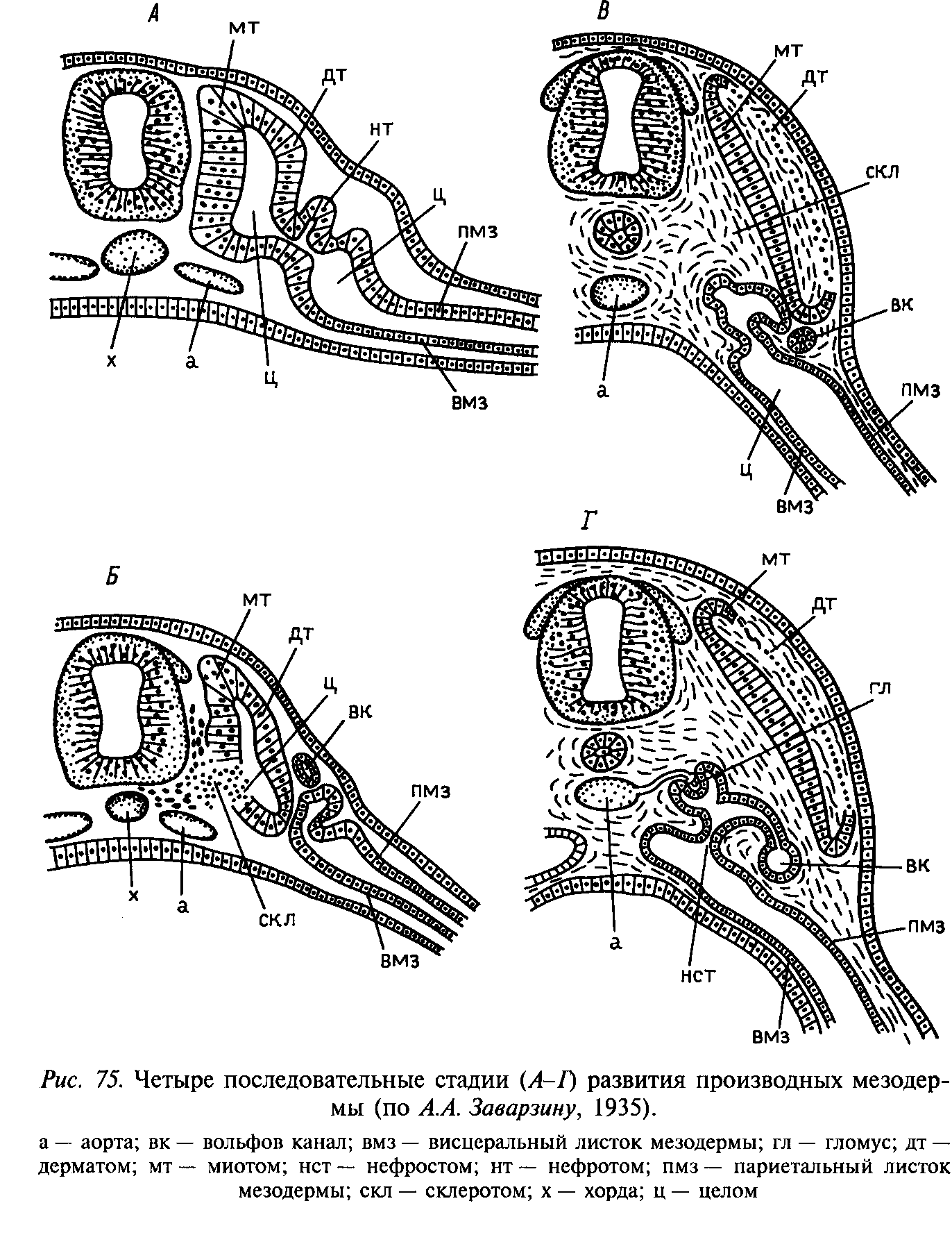
В ходе дальнейшего развития сомита из его клеток формируются три основные закладки:
- дерматом – наружная, обращенная к эктодерме. Формирует кожный листок, из него впоследствии дифференцируется соединительная часть кожи.
- склеротом – внутренняя часть сомита, примыкает к хорде (у низших позвоночных) или к хорде и нервной трубке (у высших). Зачаток осевого скелета, впоследствии распадающийся на отдельные клетки.
- миотом – располагается между дерматомом и склеротомом. Зачаток всей поперечно-полосатой мускулатуры.
У низших позвоночных основная часть сомитов – миотомы. У высших позвоночных наиболее активна зона, дающая как дерматом, так и миотом (он возникает путем подворачивания под зачаток дерматома).
Осевая мезодерма метамеризуется к в туловищном, так и головном отделах, но в последствии головные сомиты у позвоночных кроме ланцетника они распадаются, из их клеток формируются парахордалии, то есть по своей потенции они соответствуют склеротомам. Спереди от них возникают парные хрящевые закладки – трабекулы.
Часы сегментации. У зародышей костистых рыб, птиц и млекопитающих был обнаружен периодический процесс в несегментированной осевой мезодерме.
На примере куриного зародыша: спереди от гензеновского узелка в еще не сегментированной осевой мезодерме каждые 90 минут возникает область активации генов (один из них – c-hairy 1), начинается синтез мРНК. Область активации распространяется вперед, до границы уже сегментированной мезодермы, сзади ее догоняет область дезактивации, таким образом в конце от активной области остается лишь узкая полоса. Ровно в этот момент от несегментированной мезодермы отделяется новый сомит, и активная полоска остается в его составе.
Исследования показали, что клетки включают и выключают данную группу генов независимо от индукции, таким образом, часы действительно автономны.
Наличие часов сегментации не является необходимым условием для возникновения сомитов.

Развитие органов выделения.
У анамний два сменяющих друг друга органа выделения – головная почка (предпочка, пронефрос) и туловищная (первичная, мезонефрос).
У амниот вслед за ними развивается метанефрос (тазовая почка).
Все три типа образуются из мезодермы ножек сомитов: - пронефрос из передних сомитов (наиболее четкая метамеризация; стенки канальцев непосредственно из ножек, поэтому канальцы открываются прямо в целом, имея на конце воронки-нефростомы, а другими концами сливаются в вольфов канал – первичный мочеточник; рост вольфова канала назад индуцирует образование мезонефрических канальцев)
- мезонефрос из почти всех туловищных сомитов (ножки к этому моменту отшнуровываются от сомитов и преобразуются в мезенхимную ткань; из них формируются метамерные канальцы с ответвлениями и изгибами; у высших позвоночных канальцы слепо заканчиваются в мезенхиме, а наружные концы открываются в первичный мочеточник; у высших позвоночных от них остаются только небольшие придатки половых желез)
- метанефрос из скопления нефрогенной мезенхимы, произошедшей из ножек сомитов (уже никаких следов метамерии; не связан с целомом ни на одной стадии развития; от первичного мочеточника в сторону нефрогенной мезенхимы идет остросток с расширенным концом, который превращается во вторичный мочеточник с почечной лоханкой)
Боковая пластинка.
Боковая пластинка располагается вентральнее ножек сомитов, рано разделяется на париетальный и висцеральный листки, соединенные брыжейками. Между ними находится целом, и оба листка формируют его выстилку – париетальный внешнюю, висцеральный внутреннуюю.
Производные висцерального листка (половые железы, сердце, кровеносные сосуды и клетки крови). Для полноценного развития нуждаются в контакте с энтодермой.
- Половые железы и протоки.
Развиваются из висцерального листка боковой пластинки на уровне ножек сомитов – герминативного эпителия (половые клетки возникают из гоноцитов и мигрируют позже). Герминативный эпителий – соматическая ткань, формирующая стенки половой железы. Сама железа вначале представляет собой впячивание – половую складку, которая постепенно заполняется мезенхимой, из которой развивается мозговая часть железы.
Поначалу железа имеет одинаковое строение у обоих полов, но под воздействием проникших первичных половых клеток и гормонального баланса дифференцируется в семенник (преимущественно мозговой слой) или яичник (преимущественно корковая часть).
- У самцов семенные канальцы впадают в вольфовы каналы, которые становятся семяпроводами. У амниот это единственная функция вольфовых каналов (так как у тазовой почки вторичный мочеточник), у анамний они же являются мочеточниками.
- У самок имеются мюллеровы каналы (закладываются параллельно вольфовым, но у самцов дегенерируют), превращаются в яйцеводы.
У многих анамний развиваются верхние отделы мюллеровых каналов и открываются в целом воронкой нефростома, превратившейся в воронку яйцевода, так что яйцо сперва выходит в целом, а затем захватывается воронкой.
- Развитие сердца.
У птиц парный зачаток сердца возникает на вторые сутки инкубации в виде двух симметрично расположенных утолщений висцерального листка мезодермы, который тесно связан с энтодермой. Зачатки соединяются лишь после сворачивания энтобласта в трубку головной кишки. Из объединившейся висцеральной мезодермы возникает миокард. Эндокард формируется при слиянии трубчатых зачатков, образованных мигрировавшими мезенхимными клетками.
Единая сердечная трубка переходит в желточные вены, несущие кровь от стенки желточного мешка. Она лежит в перикардиальной полости, являющейся частью целома.
- Кровеносные сосуды. Развиваются из мезенхимы, закладываются в виде кровяных островков, отдельные трубочки сливаются в рыхлую сеть. Наружные клетки островков (ангиобласт) вступают в контакт друг с другом и формируют эндотелий, внутренние клетки (гемобласт) формируют клетки крови. Парные желточные вены впадают в зачаток сердца сзади и несут кровь от внезародышевых органов у амниот.
Ствол аорты делится на два артериальных ствола. Сначала формируется 6 дуг аорты по числу жаберных дуг, у высших позвоночных впоследствии большая часть редуцируется. Изначально формируется избыток сосудов, из которых потом остаются те, направление которых соответствует анатомическим особенностям тела взрослого животного.
Производные париетального листка – конечности.
- Конечности. Развиваются из выселившихся из париетального листка мезодермы мезенхимных клеток и покровной эктодермы.
У зародышей амфибий ранние зачатки конечностей имеют вид обособленных бугорков.
У зародышей амниот вначале формируются длинные складки (вольфовы гребни), которые позже редуцируются в средней части, а из переднего и заднего участка развиваются передние и задние конечности.
На ранних стадиях роста эктодермальный эпителий пассивно растягивается делящейся мезенхимой, но затем начинает активно участвовать в росте конечностей. У амниот эктодерма верхушки конечности утолщается, образуя апикальный гребешок.
Апикальная часть по мере роста расширяется и уплощается, зачаток скручивается вдоль своей продольной оси, на апикальной поверхности появляются зачатки пальцев (обособение пальцев связано с апоптозом клеток между ними).
Параллельно идет образование внутреннего скелета путем образования хрящей из мезенхимы. Выделяется стилоподия (зачаток проксимального хряща), их которой разовьются плечевые/бедренные кости; затем зигоподия (локтевой и лучевой хрящи; большой и малый берцовые) и аутоподия (хрящи кисти или стопы). Хрящи поясов конечностей формируются по времени между стило- и аутоподиями.
В конечность прорастают кровеносные сосуды и миобласты.
Интенсивные эпителио-мезодермальные взаимодействия. На начальной стадии мезодерма влияет на эктодермальный эпителий (он утолщается и начинает активно расти).
Нормальное развитие пальцев определяет обратное влияние на мезодерму со стороны эпителия верхушки почки конечности (апикального гребешка). При его удалении фаланги не дифференцируются. (При пересадке апикального гребешка на презумптивную мезодерму проксимальной части конечности развивается дистальный отдел; при пересадке проксимальной мезодермы задней конечности под гребешок передней, она даст дистальную часть задней конечности.)
Зона поляризующей активности – морфогенетически активная зона зачатка конечности на заднем крае, у основания. Если ее пересадить на передний край, то произойдет ее зеркальное удвоение – появится второй задний край с соответствующими пальцами. ЗПА характеризуется повышенной концентрацией морфогенетически активной ретиноевой кислоты (витамина А), градиент концентрации которой определяет передне-заднюю полярность.
В передних конечностях экспрессируется ген Tbx5, а в задних - Tbx4, продукты которых – факторы транскрипции, т.е. от них зависит активность других генов.
Важную роль играют белки группы факторов роста фибробластов. Для возникновения зоны поляризующей активности необходим ФРФ-10, а процессы апоптоза в пальцах контролируются белками ВМР.
Птицы.
Яйца птиц относятся к меробластическому типу, дробление дискоидальное. У верхнего полюса бластодиска формируется узкий клеточный слой – серп Коллара, который начинает расти в подзародышевую полость, в которую ранее выселяются отдельные клетки и образуют первичный гипобласт. Затем первичный гипобласт объединяется с серпом Коллара во вторичный гипобласт (носитель передне-задней полярности зародыша). Гипобласт дает начало внезародышевой энтодерме, которая обрастает желток и участвует в формировании желточного мешка.
Наружный слой бластодиска после выселения из него клеток называется эпибластом.
Ц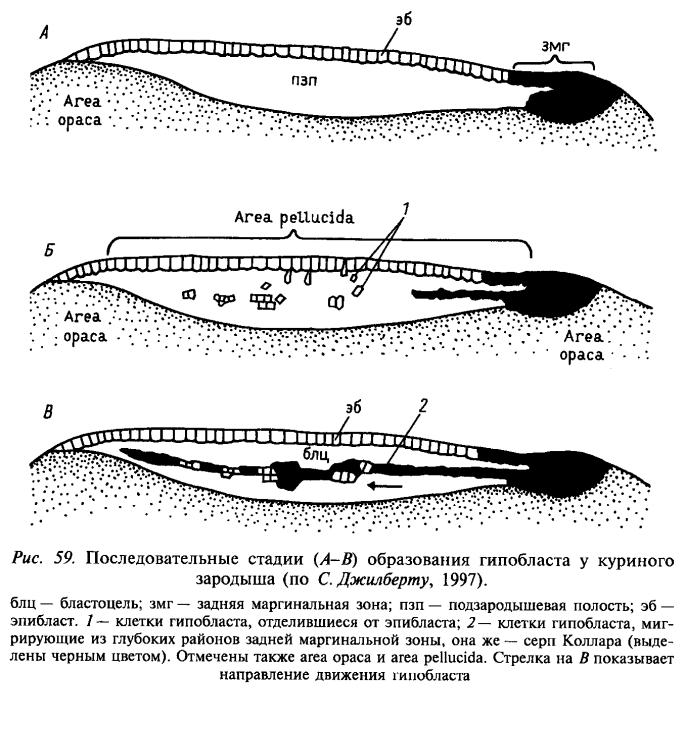 ентральная
зона бластодермы после разделения
выглядит прозрачной и носит название
area
pellucida.
Перибласт (краевое кольцо бластодермы)
выглядит темным и называется area
opaca.
На заднем конце зародыша между этими
двуми зонами возникает первичная
полоска,
после чего приобретает углубление и
начинает называться первичной бороздкой.
На переднем конце возникает первичная
ямка,
или гензеновский
узелок
(из области узелка начинают иммигрировать
под эпибласт клетки дефинитивной
кишечной энтодермы - энтобласта).
ентральная
зона бластодермы после разделения
выглядит прозрачной и носит название
area
pellucida.
Перибласт (краевое кольцо бластодермы)
выглядит темным и называется area
opaca.
На заднем конце зародыша между этими
двуми зонами возникает первичная
полоска,
после чего приобретает углубление и
начинает называться первичной бороздкой.
На переднем конце возникает первичная
ямка,
или гензеновский
узелок
(из области узелка начинают иммигрировать
под эпибласт клетки дефинитивной
кишечной энтодермы - энтобласта).
Формирование энтобласта – начало второй стадии гаструляции. Клетки энтобласта оттесняют гипобласт на периферию зародышевого щитка. Вслед за энтобластом через гензеновский узелок и стенки первичной бороздки мигрируют клетки будущего мезобласта (среднего зародышевого листка). Эпителио-иезенхимный переход при миграции.
Центральная часть мезобласта, мигрировавшая через гензеновский узелок дает начало головной мезодерме и хордальному выросту (и затем хорде).
Материал, мигрировавший через боковые стенки первичной бороздки, дает начало внезародышевой мезодерме (кровяные островки и сосуды).
Материал, мигрирующий последним (ближе к оси зародыша) даже мезодерму боковой пластинки, а самый поздний – сомиты.
Бороздка постепенно укорачивается, узелок смещается назад, а затем превращается в анальное отверстие.
Вторая фаза гаструляции гомологична гаструляции амфибий. Первичная бороздка гомологична бластопору, ее края – губам бластопора, гензеновский узелок – дорсальной губе бластопора (подтврждается экспериментально; первичные организаторы гаструляции). Материал энтобласта гомологичен прехордальной пластинке амфибий.
Образование туловищных складок.
В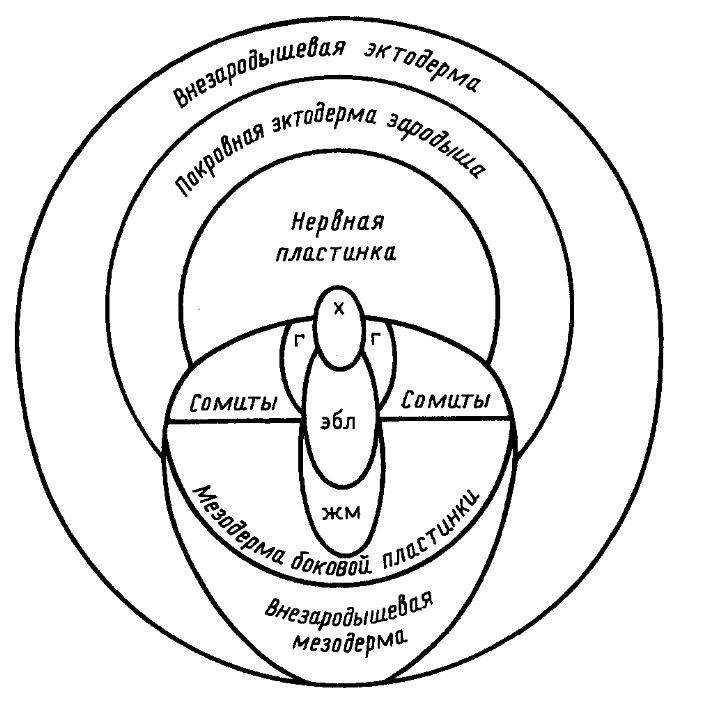 конце первых суток инкубации передний
конец зародыша начинает приподниматься
над поверхностью бластодиска и отделяться
от него узкой впадиной – головной
складкой. Края этой складки распространяются
назад и отделяют зародыш от внезародышевой
части, формируя туловищные складки.
конце первых суток инкубации передний
конец зародыша начинает приподниматься
над поверхностью бластодиска и отделяться
от него узкой впадиной – головной
складкой. Края этой складки распространяются
назад и отделяют зародыш от внезародышевой
части, формируя туловищные складки.
Одновременно от желтка начинает отделяться передняя часть энтобласта и образует выпячивание – головную кишку, а вход в нее сзади называется воротами головной кишки.
В начале вторых суток возникает парный зачаток сердца (утолщения висцерального листка мезодермы, тесно связанной с энтодермой). При сворачивании энтобласта в трубку головной кишки зачатки сердца объединяются, в миокард. В конце вторых суток эндокард сливается из двух трубчатых образований из мигрировавших по энтобласту мезенхимных клеток. Сердечная трубка соединяется с желточными венами.
После 30-часовой инкубации из осевой мезодермы дифференцируются десять пар мезодермальных сомитов, замыкается передняя часть нервной трубки и подразделяется на три мозговых пузыря.
Развитие зародышевых оболочек.
З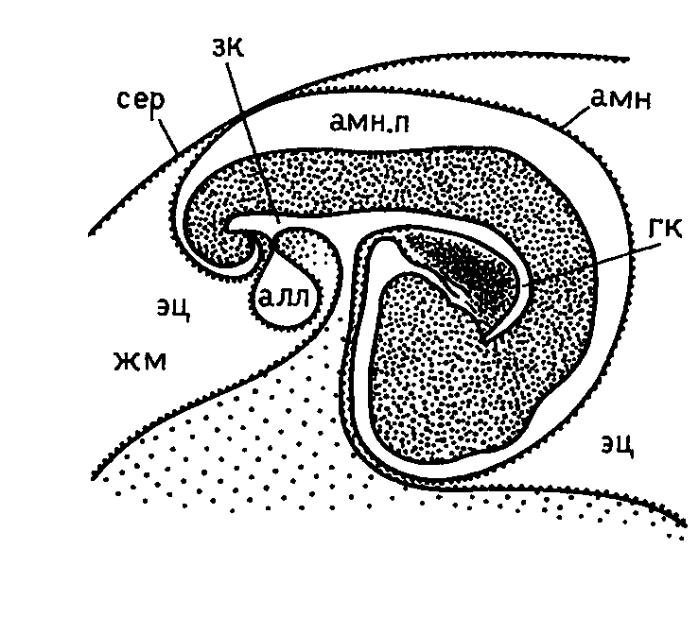 ародышевые
оболочки начинают формироваться в конце
вторых суток инкубации. Они формируются
как складки внезародышевой эктодермы
и прилегающего к ней париетального
листка мезодермы, после чего эти складки
удлиняются и смыкаются над телом
зародыша, швы между ними исчезают,
складки объединяются и формируют две
оболочки (каждая состоит из эктодермы
и примыкающей мезодермы). Внутренняя
оболочка – амнотическая (отграничивает
амниотическую полость), наружная -
серозная (отграничивает внезародышевый
целом – экзоцелом, который с обеих
сторон выстелен внезародышевой
мезодермой).
ародышевые
оболочки начинают формироваться в конце
вторых суток инкубации. Они формируются
как складки внезародышевой эктодермы
и прилегающего к ней париетального
листка мезодермы, после чего эти складки
удлиняются и смыкаются над телом
зародыша, швы между ними исчезают,
складки объединяются и формируют две
оболочки (каждая состоит из эктодермы
и примыкающей мезодермы). Внутренняя
оболочка – амнотическая (отграничивает
амниотическую полость), наружная -
серозная (отграничивает внезародышевый
целом – экзоцелом, который с обеих
сторон выстелен внезародышевой
мезодермой).
Ворота головной кишки по мере развития все сильнее смещаются назад, отделяя от желтка новые участки энтобласта. На вторые сутки на заднем конце формируется задняя кишка, так что к четвертым суткам инкубации ворота передней и задней кишки почти смыкаются, оставляя узкий просвет между кишечником и желточным мешком – желточный стебелек (при вылуплении он с остатками желточного мешка втягивается в кишечник).
К концу третьих суток появляется аллантоис, образованный энтодермой и прилежащим листок висцеральной мезодермы, представляет собой вырост задней кишки зародыша. Он работает как зародышевый орган выделения и орган дыхания (в мезодермальной оболочке разививается мощная сеть сосудов, а аллантоис разрастается, занимая всю полость внезародышевого целома и прирастает к серозе; формируется богатая сосудами оболочка – хориоаллантоис).
При вылуплении большая часть отбрасывается, а незначительная втягивается и формирует мочевой пузырь.
Внезародышевые органы выполняют разные функции и различаются по тому, какие зародышевые листки участвуют в их формировании.
Амнион и хорион образуются из слоя клеток, образованно эктодермой и париетальным листком латеральной мезодермы, называемым соматоплеврой. По своему происхождению амнион и хорион являются соматоплевральными внезародышевыми оболочками.
Стенка желточного мешка и аллантоис образуются в результате разрастания слоя клеток, образованного энтодермой и висцеральным листком латеральной мезодермы – спланхноплеврой. По своему происхождению желточный мешок и аллантоис являются спланхноплевральными внезародышевыми органами.
Желточный мешок выполняет функцию органа питания. Висцеральный листок мезодермы, входящий в состав стенки желточного мешка, развивает мощную систему кровесносных сосудов и капилляров, по которой питательные вещества из желточного мешка переносятся в тело зародыша. Желточный мешок соединяется со средней кишкой зародыша тонкой трубкой – желточным стебельком, внутри которого находится желточный проток. Однако питательные вещества через желточный проток не поступают. В утилизации желтка участвуют клетки стенки желточного мешка. Сначала энтодермальные клетки расщепляют белки желтка яйца до растворимых аминокислот, которые потом поступают в кровеносные сосуды мезодермы и с кровотоком переносятся в тело зародыша. Кроме того, желточный мешок является органом внезародышевого кроветворения. В мезодерме желточного мешка закладываются кровяные островки, которые дают первую генерацию клеток крови зародыша и являются источник стволовых кроветворных клеток. Последние мигрируют по внезародышевым кровеносным сосудам и заселяют кроветворные органы зародыша.
Амнион – внезародышевый орган, закладывается вместе с хорионом в виде складки на стенке зародышевого мешка в его головном отделе (головная амниотическая складка) в середине вторых суток инкубации. Он состоит из внезародышевой эктодермы и париетального листка внезародышевой мезодермы. Складка из этих листков с течение третьих суток инкубации нарастает на тело зародыша до уровня поворота и вдоль его тела. На третьи сутки инкубации формируется хвостовая амниотическая складка, она начинает подниматься над хвостовой частью тела зародыша, достигая примерно уровня растущей хвостовой кишки. К началу четвертых суток инкубации амниотические складки все больше нарастают над телом зародыша и покрывают его, образую сначала заметной величины отверстие, а затем смыкаются, оставляя узкое воронкообразное отверстие – сероамниотический проток.
Таким образом, над телом зародыша создается полость, которая через сероамниотический проток заполняется жидкой фракцией белка из белковой (третичной) оболочки яйца. С этого времени зародыш оказывается погруженным в жидкую среду, соответствующую водной среде обитания низших позвоночных животных. Жидкость амниона примерно на 14-ые сутки инкубации становится дополнительным к желтку источником питания цыпленка непосредственно через формирующийся рот и желудочно-кишечный тракт.
Аллантоис формируется как дивертикул задней кишки зародыша, разрастающийся в полость экзоцелома и вытесняющий его. У птиц аллантоис представляет собой большой мешок, в котором накапливаются и хранятся до вылупления цыпленка токсические продукты метаболизма зародыша. В основном это продукты азотистого обмена, накапливающиеся в виде солей мочевой кислоты.
Хорион (сероза) образует внешнюю внезародышевую оболочку эмбриона и выполняет защитную функцию. По мере разрастания аллантоиса висцеральная мезодерма, покрывающая его снаружи, прирастает к париетальной мезодерме хориона – формируется хориоаллантоис. В хориаллантоисной оболочке из материала висцеральной мезодермы формируется большое число кровеносных сосудов, формирующих густую капиллярную сеть. Она осуществляет газообмен, то есть выполняет функцию дыхания зародыша. Хориоаллантоис обеспечивает также растворение кальция скорлупы, который необходим для остеогенеза, и его транспорт к зародышу.
НЕЙРУЛЯЦИЯ И ЭКТОДЕРМА
Движения гаструляции без существенного перерыва переходят в движения нейруляции – закладки ЦНС. Нейруляция – характерный для всех позвоночных процесс, определяющий главные структурные особенности организма. Зародыш в период нейруляции называется нейрулой.
Нейруляция включает в себя процесс скручивания эктодермы в нервную трубку, а также конвергентное смещение материала эктодермы и мезодермы к средней линии спинной стороны зародыша и растяжение дорсальной эктодермы в передне-заднем направлении.
Нейральная эктодерма уплощается и превращается в нервную пластинку, более широкую в головной части зародыша (это происходит благодаря кооперативной поляризации клеток дорсальной эктодермы: клетки сильно вытягиваются, а их апикальные части сокращаются). Ее формирование начинается от дорсо-медиальной линии зародыша и распространяется латерально, клетки оказываются скошены в сторону данной линии и образуют клеточный веер. Образующаяся пластинка имеет плоскую форму, и ее дальнейшее скручивание вызывается спрямлением скошенных клеток.
Клеточный механизм подобной трансформации основан на тесном взаимодействии цитоскелета и мембран: натяжение клеток связано со сборкой микротрубочек, сокращение апикальных стенок – с сокращением актиновых микрофиламентов и резорбцией мембран, спрямление – сокращением кортекса.
Аналогичным образом формируются органы из эпителиальных пластов (хрусталик, органы слуха и обоняния, желез пищеварительной системы).
Края пластинки приподнимаются, формируя нервные валики. Затем поверхность пластинки начинает сокращаться преимущественно за счет погружения наружных клеток в ее же внутренние слои и начинает складываться по средней линии. Возникает углубление – нервный желоб. После этого края пластинки смыкаются, формируя нервную трубку, в которой имеется полость – невроцель. Передняя, широкая часть превращается в головной мозг, а ее невроцель – в полость мозгового пузыря. Туловищная часть формирует спинной мозг со спинномозговым каналом.
После смыкания нервного желобка в трубку нервные валики формируют нервный гребень (не входит в состав ЦНС, дает много производных).
[У амфибий] Из мезодермы выделяется хорда, спереди от которой находится прехордальная пластинка, выстилающая глотку и ротовую полость. Это происходит благодаря интеркаляции клеток хордомезодермы, движущихся навстречу друг другу. Хорда образуется на линии встречи клеточных потоков на дорсальной стороне. Клетки интердигитируют, и хорда удлиняется. В головном и шейном отделе движение идет в обратном, дорсо-медиальном направлении, так что массив клеток оказывается на брюшной стороне и формирует закладку сердца.
Метамеризация осевой мезодермы, сомиты, закладка органов, все дела.
Эктодерма.
Формирует ротовое впячивание (образуется поздно, затем прорывается). Дорсальная часть ротового впячивания (карман Ратке) соприкасается с дном промежуточного мозга и отшнуровывается, образуя переднюю и промежуточную доли гипофиза. Эктодерма участвует в формировании зубов, дифференцировка происходит при тесном взаимодействии с мезенхимой с самых ранних стадий. Мезенхима зубных сосочков индуцирует синтез эмалевых белков в эпителии.
Кожа развивается из эктодермы и мезодермы. Эктодерма превращается в двухслойный, а затем многослойный эпителий – кожный эпидермис, внутренний слой которого всю жизнь выполняет роль камбиального элемента. Дерма образуется соединительнотканными клетками дерматомов. Роговые придатки – чешуи, пластинки, волосы, перья, рога. Дифференцировка кожи зависит от индукционного воздействия дермы на эпидермальную часть (плюс, дерма определяет порядок возникновения придатков и окончательное расположение). Начальные этапы формирования кожных придатков можно индуцировать дермой, взятой от зародышей других классов амниот.
ЦНС. Расширенный передний отдел замкнувшейся трубки называется первичным мозговым пузырем (первичным мозгом – arhencephalon), который открывается наружу невропором, а задний отдел через нервно-кишечный канал связан с задним отделом гастроцеля. Впоследствии они зарастают. Задняя граница первичного головного мозга подстилается производной прехордальной пластинки и отмечена вентральной мозговой складкой, спереди от которой образуется воронка мозга. Воронка и вентральная складка формируют теменной изгиб.
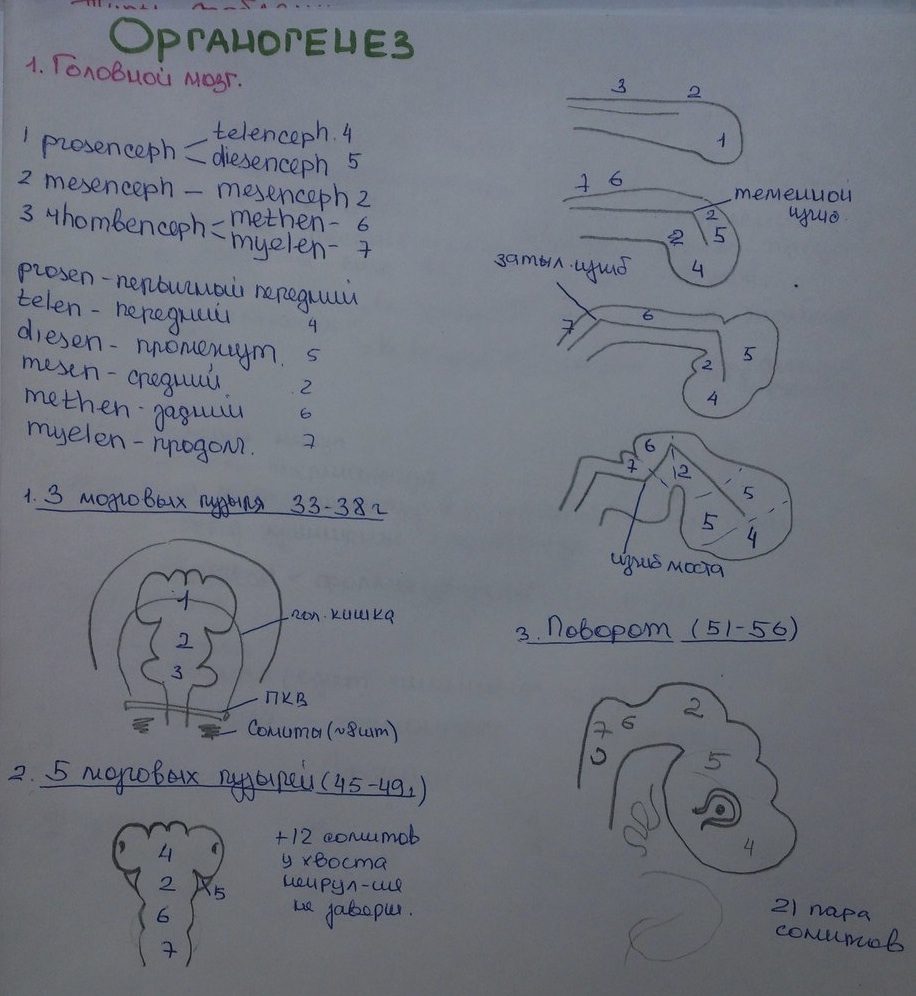
Первичный мозг дифференцируется на три мозговых пузыря: передний (prosencephalon), средний (mesencephalon) и задний (rhombencephalon), переходящий в спинной мозг. В переднем пузыре образуются выступы, дающие начало глазным зачаткам.
Передний мозговой пузырь подразделяется на передний (telencephalon) и промежуточный (diencephalon) мозг, из боковых стенок которого развиваются глазные зачатки. Средний мозговой пузырь в дальнейшем не расчленяется, а первичный задний пузырь дает задний мозг (metencephalon) и продолговатый (myelencephalon), переходящий в спинной.
У высших позвоночных вскоре после формирования пяти пузырей мозг образует резкие изгибы – затылочный и мостовой (там впоследствии возникает варолиев мост).
В области переднего мозга разрастаются стенки, образуя зачатки больших полушарий, неравномерное разрастание приводит к образованию борозд. У низших позвоночных развиваются только обонятельные доли.
В области промежуточного мозга выпячиваются глазные пузыри, а дно промежуточного мозга образует выпячивание – воронку мозга, из ее нижнец части возникает нейральная задняя часть гипофиза. Из стенки за воронкой образуется гипоталамус.
Стенка нервной трубки – ложномногослойный эпителий, то есть, все нейробласты прикреплены к внутренней поверхности нервной трубки. На более поздних стадиях развития нейробласты отрываются от нее и выходят наружу, образуя рыхлую клеточную массу – мантийный слой, где нейробласты приобретают отростки и превращаются в нейроны. Следующее поколение мигрирующих нейробластов дифференцируются в нейроглию, а оставшиеся клетки превращаются в эпендимную выстилку. Серое и белое вещество, ганглии.
Развитие глаза. По мере развития выпячиваний промежуточного мозга глазные пузыри все более отшнуровываются от зачатка промежуточного мозга, оставаясь соединенными с ним при помощи глазного стебелька. Пузыри дорастают до покровной эпидермы, и в месте соприкосновения эктодерма утолщается, образуя зачаток хрусталика – хрусталиковую плакоду. Часть глазного пузыря, контактирующая с плакодой, начинает впячиваться, в результате чего образуется двухслойный глазной бокал (индуцируется окружением, в том числе зачатком хрусталика). По мере углубления впячивания края глазного бокала начинают расти по направлению друг к другу, и некоторое время между ними остается глазная зародышевая щель. Внутренний слой бокала дает начало сетчатке, наружный – пигментный эпителий (индуцируется наличием или отсутствием контакта с мезенхимой). Край глазного бокала становится радужкой и цилиарным телом.
Клетки сетчатки проходят дифференцировку – они интенсивно делятся; первыми дифференцируются глиальные элементы сетчатки, ядра которых занимают наиболее центральное положение в зачатке – мюллеровы клетки, их отростки выходят по обе стороны и формируют пограничные мембраны. Затем дифференцируются ганглиозные клетки, которые образуют зрительный нерв. Вслед за ними дифференцируются клетки внутреннего ядерного слоя, формируют внутренний сетчатый слой. Последними дифференцируются клетки наружного ядерного слоя и образуют наружный сетчатый слой, их наружные отростки формируют палочки и колбочки.
Хрусталиковая плакода тоже впячивается, а затем полностью отшнуровывается, образуя хрусталиковый пузырек. Внутренний слой дает первичные волокна, наружный сохраняет пролиферативную активность в течение всей жизни. Синтезируются альфа- и бета-кристаллины. Развитие хрусталика индуцируется глазной чашей.
Покровный эпителий над хрусталиком истончается, теряет пигмент и становится эпителием роговицы. Индуцируется глазом
Клетки эмбриональной мезенхимы формируют сосудистую оболочку глаза и склеру.