

54. Средняя арифметическая величина и ее значение
Средняя арифметическая представляет собой как бы точку равновесия вариационного ряда, отклонения от которой в сторону увеличения или уменьшения признака взаимно уравновешиваются. Средняя арифметическая показывает, какую величину признака имели бы особи данной группы, если бы эта величина была у всех одинаковой.
Простейший метод вычисления средней арифметической величины для небольшой выборки (n30) – это простое суммирование, т.е. нахождение суммы вариант выборки и деление ее на объем выборки. Среднюю арифметическую обозначают Хср или М.
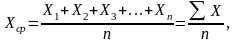
где X – величина варьирующего признака;
n – объем выборки;
– знак суммирования.
Д
 ля
больших выборок среднюю арифметическую
удобнее вычислить косвенным методом
по формуле:
ля
больших выборок среднюю арифметическую
удобнее вычислить косвенным методом
по формуле:

где А – условное среднее значение нулевого класса;
р – частоты;
а – условное отклонение;
n – объем выборки;
i – величина классового промежутка.
55. Показатели разнообразия
Генетическое разнообразие
Генетическое внутривидовое разнообразие определяется структурой аллелофонда и генофонда популяций.
Аллелофонд – это множество аллелей в популяции. Для количественного описания структуры аллелофонда используется понятие «частоты аллелей».
Генофонд – это множество генотипов в популяции. Для количественного описания структуры генофонда используется понятие «частоты генотипов».
Для описания генетического разнообразия используют следующие показатели:
– доля полиморфных генов;
– частоты аллелей для полиморфных генов;
– средняя гетерозиготность по полиморфным генам;
– частоты генотипов.
На основании этих показателей вычисляют разнообразные индексы разнообразия (например,Шеннона-Увера, Симпсона).
Первичные механизмы возникновения генетического разнообразия
Источники новых генотипов – это рекомбинации, возникающие при мейозе и половом размножении, а также в результате разнообразных парасексуальных процессов.
Основные источники новых аллелей в популяции – это мутационный процесс и иммиграцииносителей новых аллелей.
Дополнительные источники связаны с латеральным (горизонтальным) переносом генов от одного биологического вида к другому: или при межвидовой половой гибридизации, или при симбиогенезе, или при участии организмов-посредников.
Единичная мутация – это редкое событие. В стационарной популяции мутантный аллель можетсовершенно случайно не перейти в следующее поколение.
Механизмы повышения уровня генетического разнообразия
Мутационный процесс (давление мутаций) в больших популяциях
Одна и та же мутация с одной и той же частотой q возникает в каждом поколении (если принять, что численность популяции велика: миллионы особей).
В то же время мутантный аллель может быть утрачен под воздействием случайных факторов (в т.ч. и вследствие обратных мутаций). Если не учитывать обратных мутаций, то фактическая частота мутантного аллеля нелинейно возрастает. Зависимость частоты мутантного аллеля от порядкового номера поколения может быть примерно аппроксимирована логарифмической функцией. Расчеты показывают, что частота рецессивного селективно нейтрального мутантного аллеля (и вероятность его фенотипического проявления) возрастает примерно следующим образом:
Поколения |
0 |
20 |
260 |
450 |
750 |
1250 |
2100 |
3550 |
q (а), ×10 – 6 |
1 |
5 |
10 |
11 |
12 |
13 |
14 |
15 |
q2 (aa), ×10 – 12 |
1 |
25 |
99 |
121 |
144 |
169 |
196 |
225 |
Таким образом, в длительно существующей популяции (с высокой численностью) вероятность фенотипического проявления рецессивного мутантного аллеля возрастает в десятки и сотни раз за счет давления мутаций. В то же время нужно признать, что реальные популяции существуют ограниченное число поколений, поэтому давление мутаций не может принципиально изменить генетическую структуру популяций.
Дрейф генов (генетико-автоматические процессы)
Дрейф генов – это случайное изменение частоты селекционно нейтральных (илипсевдонейтральных) аллелей в малых изолированных популяциях. В малых популяциях велика роль отдельных особей, и случайная гибель одной особи может привести к значительному изменению аллелофонда.
Чем меньше популяция, тем больше вероятность случайного изменения частот аллелей. Чем меньше частота аллеля, тем больше вероятность его элиминации.
В сверхмалых популяциях (или популяциях неоднократно снижающих свою численность до критического уровня) по совершенно случайным причинам мутантный аллель может занять место нормального аллеля, т.е. происходит случайная фиксация мутантного аллеля. В результате уровень генетического разнообразия снижается.
Дрейф генов может наблюдаться и в результате эффекта генетической воронки (эффекта бутылочного горлышка): если популяция на некоторое время снижает, а затем повышает свою численность (эффект основателей новой популяции ex situ, восстановление популяции после катастрофического снижения численности in situ).
Естественный отбор (natural selection)
Естественный отбор – это совокупность биологических процессов, обеспечивающих дифференциальное воспроизведение генотипов в популяциях.
Естественный отбор – это направленный фактор эволюционного процесса, движущая сила эволюции. Направление естественного отбора называется вектором отбора.
Исходной (ведущей) формой является движущий отбор, который приводит к изменениюгенетической и фенотипической структуры популяции.
Сущность движущего отбора заключается в накоплении и усилении генетически детерминированных отклонений от первоначального (нормального) варианта признака. (В дальнейшем первоначальный вариант признака может стать отклонением от нормы.)
В ходе движущего отбора повышается частота аллелей и генотипов, обладающих максимальной приспособленностью
Таким образом, движущий отбор проявляется в виде устойчивого и, в известной мере, направленного изменения частот аллелей (генотипов, фенотипов) в популяции.
Первоначально в ходе отбора уровень биоразнообразия повышается, затем достигает максимума, а на заключительных этапах отбора – снижается.
Сцепленное наследование
Движущая форма отбора действует по немногим (лимитирующим) признакам, по немногим генотипам и аллелям немногих генов. Однако, если селектируемый ген (например, полезный аллель А) сцеплен с селективно нейтральным или псевдонейтральным геном (например, аллелемN), то частота нейтрального аллеля N также будет изменяться.
Существует два типа сцепления: истинное сцепление и квазисцепление.
При истинном сцеплении селектируемый и нейтральный аллели (A и N) находятся (локализуются) в одной хромосоме. Тогда отбор в пользу гаплотипа AN будет способствовать повышению частоты аллеля N.
При квазисцеплении аллели А и N локализованы в разных хромосомах, но они связаны между собой общностью морфогенетических процессов (онтогенетическими корреляциями). Тогда, как и в предыдущем случае, частота селективно нейтральногоаллеля N будет изменяться.
Механизмы поддержания высокого уровня генетического разнообразия в природных популяциях
Безразличное равновесие в больших популяциях
В популяциях с высокой численностью случайно сложившаяся (например, за счет эффекта бутылочного горлышка) структура алелелофонда может сохраняться в течение длительного времени, даже если рассматриваемый признак является селекционно нейтральным.
Миграции
Миграции – это перемещения носителей генетической информации (особей, семян, спор) из одной популяции в другую. При наличии миграций эффект дрейфа генов и естественного отбора резко снижается. В результате:
а) предотвращается вырождение генетической структуры популяции (не происходит элиминации аллелей, генотипов, признаков);
б) в популяции могут обнаруживаться признаки (аллели, генотипы), снижающие её приспособленность.
Естественный отбор на разнообразие
Кроме движущего отбора, изменяющего генетическую структуру популяций, существует множество форм отбора, сохраняющих эту структуру.
Отбор в пользу гетерозигот. Во многих случаях гетерозиготы приспособленылучше, чем любая из гомозигот (явление гетерозиса, или сверхдоминирование). Тогда в популяции действует стабилизирующий отбор на разнообразие, сохраняющий существующий уровень биологического разнообразия.
Частотно-зависимый отбор действует в том случае, если приспособленность фенотипа (генотипа, аллеля) зависит от его частоты. В простейшем случае феномен частотно-зависимого отбора определяется внутривидовой (внутрипопуляционной)конкуренцией. В этом случае приспособленность гено/фенотипа обратно пропорциональна его частоте: чем реже встречается данный гено/фенотип, тем выше его приспособленность. При возрастании частоты гено/фенотипа его приспособленность снижается.
Эти формы отбора могут наблюдаться в чистом виде, но могут сочетаться и с другими формами отбора, например, отбор в пользу гетерозигот в сочетании с родственным (групповым отбором) или частотно-зависимый отбор в сочетании с половым отбором.
Родственный (групповой) отбор действует в том случае, если признак, нейтральный или даже вредный для данной особи (с данным фенотипом, генотипом, аллелем), оказывается полезным для группы (семьи). Эта форма отбора приводит к формированию альтруистических признаков.
Половой отбор – это форма естественного отбора, основанная на соперничестве особей одного пола (чаще мужского) за спаривание с особями другого пола. При этом приспособленность генотипа (фенотипа) оценивается не по его выживаемости, а по его участию в размножении. Эта форма отбора может приводить к появлению и сохранению признаков, снижающих выживаемость (конституциональную приспособленность) их носителей.
Сцепленное наследование
Если стабилизирующий отбор (или иная форма отбора, благоприятствующая сохранению генетической структуры популяции) действует по селектируемому гену А, то будет сохраняться не только определенная частота этого аллеля, но и определенные частоты сцепленных с ним нейтральных аллелей.