

7. Потенциал действия и его фазы. Изменение проницаемости калиевых, натриевых и кальциевых каналов в процессе формирования потенциала действия.
Если на клетку нанести раздражение достаточной силы, клетка придет в новое, активное состояние.
При нанесении раздражения увеличивается проницаемость мембраны для натрия. Это происходит за счет энергии раздражителя и связано с активацией небольшого числа натриевых каналов. Возникает небольшое усиление входящего натриевого тока, интенсивность входящего натриевого тока пропорционально силе раздражителя. За счет этого процесса происходит уменьшение полярности мембраны по сравнению с исходным (с 70 мВ до 40-50 мВ). Эти изменения называются - пассивная деполяризация или частичная деполяризация, выраженность и скорость этого процесса зависит от силы раздражителя.
Если силы раздражителя недостаточно, чтобы сместить ПМ до некого критического уровня, то происходит возращение ПМ к исходному уровню, т.е. к уровню ПП. Возникшие изменения ПМ называются - локальный ответ.
Если силы раздражителя достаточно, чтобы сместить ПМ до критического уровня деполяризации, то произойдет формирования потенциала действия (ПД), что свидетельствует о возбуждении клетки переходе ее в деятельное состояние.
Критический уровень деполяризации/КУД/- это такая величина разности потенциалов (40-50 мВ), при которой активируется большое количество потенциалзависимых быстрых натриевых каналов, проницаемость мембраны для натрия становится максимальной и перестает быть зависимой от силы раздражителя.
Возникает лавинообразный входящий натриевый ток, который быстро (доли мс) смещает потенциал мембраны до 0 (активная деполяризация - потеря полярности), а затем его силы хватает чтобы изменить знак мембраны на противоположный - плюс 10-20 мВ. (Смена знака потенциала мембраны называется овершут или реверсия потенциала).
Входящий натриевый ток формирует восходящую часть пика (спайка) потенциала действия, наличие которого указывает на сформировавшееся возбуждение клетки. Амплитуда пика не зависит от силы раздражителя - закон «все или ничего», будем рассматривать в следующей лекции.
Наличие восходящей части пика ПД свидетельствует, что клетка перешла в новое функциональное состояние – состояние возбуждения, т.е. в деятельное состояние.
Вторая половина ПД (нисходящая) состоит из трех частей:
1. Нисходящая часть пика ПД (от острия пика до КУД), формируется быстро (за доли мс), 2. Положительный следовой потенциал (от КУД до ПП) формируется медленнее (несколько мс),
3. Отрицательный следовой потенциал(несколько мс).
1 и 2 части обеспечиваются процессом реполяризации, 3 часть - процессом гиперполяризации.
Процесс реполяризации - возвращение, восстановление полярности мембраны клеток, которое для них характерно в покое. Процесс реполяризации обусловлен:
1. активацией потенциалзависимых быстрых калиевых каналов, которая(активация) возникает при ПМ 0- плюс 5 мВ), что приводит к возникновению значительного по объему выходящего калиевого тока.
2. быстрой инактивацией потенциалзависимых натриевых каналов, которая возникает сразу после достижения высшего значения ПД (+10,+20мв). Это блокирует входящий натриевый ток.
3. значительной активацией калий-натриевого насоса (увеличение скорости оборота), которая обеспечивает удаление избытка натрия в клетке, возникшего в фазу деполяризации.
Эти три процесса обеспечивают возвращение ПМ до уровня ПП.
Следует заметить, что эти три процесса ионного транспорта инертны и не инактивируются мгновенно при достижении ПМ уровня ПП, что приводит к избыточному перемещению ионов и, как следствие, к избыточной поляризации мембраны (гиперполяризации), за счет которой (гиперполяризации) и формируется отрицательный следовой потенциал. Затем ПМ мембраны клетка приходит в исходное состояние.
Следует иметь ввиду, что при формировании ПД выходящий калиевый ток, осуществляемый через неуправляемые медленные калиевые каналы по объему ничтожно мал по сравнению с объемом перемещения натрия и калия через быстрые потенциалзависимые каналы, так как процесс формирования ПД происходит за мс.
ВАЖНО. Включение других потенциалзависимых каналов существенно влияет на форму и продолжительность потенциала действия. Так, активация долговременных потенциалзависимых кальциевых каналов (L-тип), их много в кардиомиоцитах, формирует фазу плато, клетка длительно деполяризована.
Потенциал действия всегда, сформировавшись, распространяется на соседние участки клетки, локальный ответ остается там, где он возник: он не способен распространяться.
Дополнительно:
Мембранным потенциалом покоя (МПП) или потенциалом покоя (ПП) называют разность потенциалов покоящейся клетки между внутренней и наружной сторонами мембраны. Внутренняя сторона мембраны клетки заряжена отрицательно по отношению к наружной. Принимая потенциал наружного раствора за нуль, МПП записывают со знаком «минус». Величина МПП зависит от вида ткани и варьирует от -9 до -100 мв. Следовательно, в состоянии покоя клеточная мембранаполяризована. Уменьшение величины МПП называют деполяризацией, увеличение — гиперполяризацией, восстановление исходного значения МПП —реполяризацией мембраны.
Основные положения мембранной теории происхождения МПП сводятся к следующему. В состоянии покоя клеточная мембрана хорошо проницаема для ионов К+ (в ряде клеток и для СГ), менее проницаема для Na+ и практически непроницаема для внутриклеточных белков и других органических ионов. Ионы К+диффундируют из клетки по концентрационному градиенту, а непроникающие анионы остаются в цитоплазме, обеспечивая появление разности потенциалов через мембрану.
Возникающая разность потенциалов препятствует выходу К+ из клетки и при некотором ее значении наступает равновесие между выходом К+ по концентрационному градиенту и входом этих катионов по возникшему электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, называется равновесным потенциалом.
Потенциал действия -это быстрое колебание мембранного потенциала ,сопровождающееся перезарядкой клеточной мембраны ,возникающее при возбуждении
Фазы потенциала действия
1. Локальный ответ (открываются медленные натриевые каналы –ионы натрия поступают в клетку и происходит медленная деполяризация мембраны).
2. Деполяризация (при достижении критического уровня медленная деполяризация сменяется быстрой, открываются быстрые натриевые каналы и ионы натрия устремляются в клетку и мембранный потенциал становиться «+». Т.е. происходит перезарядка мембраны).
3. Реполяризация (это фаза восстановления- натриевые каналы закрываются и открываются калиевые, ионы калия выходят из клетки).
4. Отрицательный следовой потенциал (ионы калия медленно выходят из клетки).
5. Положительный следовой потенциал (связан с активацией натриевого насоса, который удаляет ионы натрия из клетки).
Проведение возбуждения
Проведение возбуждения по нервному волокну осуществляется за счет локальных токов между возбужденным и покоящимися участками мембраны. Последовательность событий в этом случае представляется в следующем виде.
При нанесении точечного раздражения на нервное волокно в соответствующем участке мембраны возникает потенциал действия. Внутренняя сторона мембраны в данной точке оказывается заряженной положительно по отношению к соседней, покоящейся. Между точками волокна, имеющими различный потенциал, возникает ток (локальный ток), направленный от возбужденного (знак (+) на внутренней стороне мембраны) к невозбужденному (знак (-) на внутренней стороне мембраны) к участку волокна. Этот ток оказывает деполяризующее влияние на мембрану волокна в покоящемся участке и при достижении критического уровня деполяризации мембраны в данном участке возникает МПД (Мембранный потенциал действия). Этот процесс последовательно распространяется по всем участкам нервного волокна.
В некоторых клетках (нейронах, гладких мышцах) МПД имеет не натриевую природу, а обусловлен входом ионов Ca2+ по потенциал-зависимым кальциевым каналам. В кардиомиоцитах генерация МПД связана с входящими натриевым и натрий-кальциевым токами.
№11 Мембранный потенциал - это разность потенциалов между наружной и внутренней поверхностями элементарной пограничной мембраны клетки
Мембранный потенциал – сила электростатического взаимодействия, которая уравновешивает электрические токи, протекающие через мембрану изнутри кнаружи, снаружи – внутрь.
Современная теория Ходжкина-Хаксли-Катца:
1. В основе возникновения биопотенциала является обмен веществ и энергии между клеткой и окружающей средой
2. Местом возникновения биопотенциала является поверхностная мембрана клетки
3. Непосредственной причиной возникновения биопотенциала является разность концентрации ионов (ионная асимметрия вне и внутри клетки)
4. Поверхностная пограничная мембрана клетки обладает свойством полупроницаемости, которое может меняться при различных состояниях.
№12 см №10
№13??? Соотношение фаз возбудимости, ПД и сокращения.
•При нанесении одиночного порогового и сверхпорогового раздражения на мышцу, мышца сокращается, а затем расслабляется, то есть наблюдается одиночное сокращение мышц.
•Имеет место ряд последовательных явлений:
•после нанесения раздражения сокращение мышцы наступает не сразу, а через некоторый интервал 2,5 мсек. (латентный период).
•Реакция мышцы на раздражение начинается с генерации ПД продолжительностью 3-5 мсек. и в это же время
•начинается сокращение мышцы, приблизительно 50 мсек. (фаза сокращения),
•затем следует фаза расслабления, по продолжительности либо равна или может быть несколько больше фазного сокращения. Длительность зависит от морфофункциональных свойств.
•Так, у наиболее быстро сокращающихся волокон глазных мышц фаза напряжения равна 7-10 мсек., а у медленных волокон камбаловидной мышцы равна 50-100мсек
Возбудимость мышц в ходе одиночного сокращения меняется.
•Во время развития ПД, т.е. в начале сокращения мышцы наступает абсолютная рефрактерная фаза(состояние невозбудимости, длительностью 3-5 мсек.).
•За ней следует фаза относительной рефрактерности, а затем
•возникает фаза повышенной возбудимости(примерно 20 мсек.).
•Длительность этих периодов различна у животных разных видов и зависит от функционального состояния нервно-мышечного аппарата.
Дополнительно: Соотношение фаз потенциала действия и возбудимости
Когда ткань возбуждается – генерирует ПД, то временно (соответственно с длительностью ПД) в ней меняется возбудимость: вначале ткань становится совершенно невозбудимой (абсолютная рефрактерность) – любой по силе стимул не способен вызвать в ней новый приступ возбуждения. Эта фаза обычно наблюдается во время пика ПД. Затем происходит постепенное восстановление возбудимости до исходного состояния (фаза относительной рефрактерности) – в этот момент раздражитель может вызвать возбуждение (генерацию нового ПД), но для этого он должен быть намного больше порогового (исходного). Затем (в фазу следовой негативности) возбудимость повышается (супервозбудимость, или фаза экзальтации). В этот момент подпороговые раздражители могут вызывать возбуждение. Наконец, в тканях, в которых ярко проявляется следовая гиперполяризация, наблюдается еще одна фаза – субнормальной возбудимости (сниженной возбудимости).
№14 Законы раздражения отражаютопределенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани.
Законы проведения возбуждения в нервах.
Анатомической и физиологической целостности нервного волокна.
Двухстороннего проведения возбуждения.
Изолированного проведения возбуждения.
К законам раздраженияотносятся:
•закон силы,
•закон "все или ничего",
•закон аккомодации (Дюбуа-Реймона) ,
•закон силы-времени (силы-длительности),
•закон полярного действия постоянного тока,
•закон физиологического электротона.
Закон силы:
•чем больше сила раздражителя, тем больше величина ответной реакции. В соответствии с этим законом функционируют сложные структуры, например, скелетная мышца. Амплитуда ее сокращений от минимальных (пороговых) величин постепенно увеличивается с увеличением силы раздражителя до субмаксимальных и максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. Поэтому на пороговые раздражители отвечают только те мышечные волокна, которые имеют самую высокую возбудимость, амплитуда мышечного сокращения при этом минимальна. С увеличением силы раздражителя в реакцию вовлекается все большее и большее количество мышечных волокон и амплитуда сокращения мышцы все время увеличивается. Когда в реакцию вовлечены все мышечные волокна, составляющие данную мышцу, дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения.
Закон "все или ничего"
•подпороговые раздражители не вызывают ответной реакции ("ничего"), на пороговые раздражители возникает максимальная ответная реакция ("все"). По закону "все или ничего" сокращаются сердечная мышца и одиночное мышечное волокно. Закон "все или ничего" не абсолютен. Во-первых, на раздражители подпороговой силы не возникает видимой ответной реакции, но в ткани происходят изменения мембранного потенциала покоя в виде возникновения местного возбуждения (локального ответа). Во-вторых, сердечная мышца, растянутая кровью, при наполнении ею камер сердца, реагирует по закону "все или ничего", но амплитуда ее сокращения будет больше по сравнению с сокращением сердечной мышцы, не растянутой кровью.
Закон раздражения Дюбуа-Реймона (аккомодации):
•раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени.
•При действии медленно нарастающегораздражителя возбуждение не возникает, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации. Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации.
При снижении скорости нарастания силы раздражителядо некоторого минимального значения потенциал действия вообще не возникает. Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого -окончанию потенциала действия.
•При медленном нарастании токана первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще. Способность к аккомодации различных структур неодинакова. Наиболее .высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
•При быстром нарастании стимулаповышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости.
Аккомодация возбудимых тканей
•Раздражители характеризуются не только силой и длительностью действия, но и скоростью роста во времени силы воздействия на объект, т. е. градиентом.
•Уменьшение крутизны нарастания силы раздражителя ведет к повышению порога возбуждения, вследствие чего, ответ биосистемы при некоторой минимальной крутизне вообще исчезает. Это явление названо аккомодацией.
•Зависимость между крутизной нарастания силы раздражения и величиной возбуждения определена в законе градиента:реакция живой системы зависит от градиента раздражения: чем выше крутизна нарастания раздражителя во времени, тем больше до известных пределов величина функционального ответа.
Закон силы-длительности:
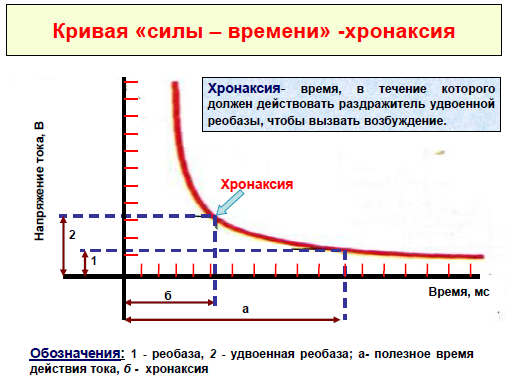
•раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать для возникновения возбуждения. Зависимость силы-длительности имеет гиперболический характер. Из этого следует, что ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Причиной такой зависимости является мембранная емкость. Очень "короткие" токи просто не успевают разрядить эту емкость до критического уровня деполяризации. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой.
Хронаксия
•Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности.
•Кривая силы –длительности, или силы –времени, изученная при исследовании различных нервов и мышц Гарвегом (1892), Вейсом (1901) и Лапиком (1909) свидетельствует о том, что ток ниже некоторой минимальной силы или напряжения не вызывает возбуждения, как бы длительно он ни действовал.
•Минимальная сила постоянного тока, способная вызвать возбуждение (порог раздражения), названа реобазой. Наименьшее время, в течение которого должен действовать раздражающий стимул, величиной в одну реобазу, называется полезным временем.При очень коротких стимулах кривая силы-времени становится параллельной оси ординат. Т.е. при таких кратковременных раздражениях возбуждение не возникает, как бы ни была велика сила раздражителя.
•Хронаксия –это время, в течение которого должен действовать ток удвоенной реобазы, чтобы вызвать возбуждение.
Использование хронаксиметрии полезно в неврологической практике(возможность установить наличие органического поражения двигательного нерва, поскольку величина реобазы и хронаксии нервных волокон значительно меньше соответствующих величин мышечных волокон), поэтому при пороговых силах тока возбуждение прежде всего возникает в нервных волокнах и от них передается на мышцу. Следовательно, при измерении хронаксии мышцы фактически получают значения хронаксии, иннервирующих ее нервных волокон. Если нерв поврежден или перерожден,тогда раздражающий стимул выявляет хронаксию собственно мышечных волокон, которая имеет большую продолжительность.
Виды хронаксиметрии
•В зависимости от возбудимости тканей, подвергаемых исследовании: двигательная, чувствительная, зрительная, вестибулярная.
•Двигательная хронаксия.
С помощью умеренной силы тока определяют моторную точку, после чего устанавливают пороговую реакцию, затем удваивают реобазу, постепенно увеличивая длительность импульсов до получения такой же пороговой реакции.
•Чувствительная хронаксия.
В данном случае вместо определения моторного эффекта определяют при какой силе тока ребенок почувствует легкий удар; ползание мурашек или тепло на коже при раздражении тактильных, болевых или термических рецепторов.
•Границы тактильной хронаксии: 0,06-0,07 Мсек;
•Болевой: 0,30-3,5 Мсек;
•Термической: 0,60-7 Мсек.
•Вестибулярная хронаксия
Активный электрод располагается на pr. mastoideus. Определяют силу, при которой возникает головокружение N12-24 Мсек, вызывающую наклонение головы.
Диагностическое значение
•Нормальные величины двигательной, чувствительной и вестибулярной хронаксии говорят о хорошем состоянии нервно-мышечного аппарата, ЦНС, вестибулярного аппарата.
•Наибольшие значения имеют двигательная и чувствительная хронаксия при поражении периферической нервной системы.
•В первые 10 дней после травмы обычно имеются только количественные изменения. При тяжелых травмах реобаза может резко удлиняться. Через 10 дней, в зависимости от тяжести травмы и отдаленности исследуемой мышцы, постепенно хронаксия удлиняется в различной степени. Наряду с изменениями хронаксии, наступающими в пораженном нервно-мышечном аппарате, возникают таковые и в строго симметричных участках.
•Согласно данным Bourgignon, при дифтерии страдает вестибулярная хронаксия; при сывороточных интоксикациях удлиняется, прежде всего, хронаксия мышц, имеющих короткую хронаксию.
•При диабете затрагивается как двигательная, так и чувствительная хронаксия. Прежде всего, удлиняется двигательная хронаксия m. ext. hal. longusи чувствительные корешки S1и S2.
При наличии болевых синдромов в мышечно-суставном аппарате детей и взрослых двигательная и чувствительная хронаксия укорачивается, при уменьшении болей –постепенно нормализуется.
•При церебральных детских параличах и парезах, у людей с параличами вследствие поражения центральных двигательных нейронов развивается асимметрия; а мышцы, имеющие отдаленные места прикрепления, постепенно показывают более высокую реобазу и слегка удлиненную хронаксию; а мышцы, имеющие приближенные места прикрепления –обратные соотношения. Хронаксия изменяется и при миопатиях. При миотонии удлиняется хронаксия.
Закон полярного действия постоянного тока:
•при замыкании тока возбуждение возникает под катодом, а при размыкании -под анодом. Прохождение постоянного электрического тока через нервное или мышечное волокно вызывает изменение мембранного потенциала покоя.
•Так, в области приложения к возбудимой ткани катода положительный потенциал на наружной стороне мембраны уменьшается, возникает деполяризация, которая быстро достигает критического уровня и вызывает возбуждение.
•В области же приложения анода положительный потенциал на наружной стороне мембраны возрастает, происходит гиперполяризация мембраны и возбуждение не возникает. Но при этом под анодом критический уровень деполяризации смещается к уровню потенциала покоя. Поэтому при размыкании цепи тока гиперполяризация на мембране исчезает и потенциал покоя, возвращаясь к исходной величине, достигает смещенного критического уровнями возникает возбуждение.
Особенности действия постоянного тока на ткани
•При раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания –только под анодом. Открыт Пфлюгером в 1859 г.
•Было установлено, что если на нерв воздействовать слабым постоянным током, то его возбудимость под катодом повышается, а под анодом снижается.
•Впоследствии Б.Ф. Вериго (1883) показал, что как повышение возбудимости под катодом, так и снижение под анодом характерно только для первоначального действия постоянного тока.
•При длительном действии под катодом развивается катодическая депрессия, и возбудимость снижается, становясь меньше исходной. А под анодом может возникнуть явление анодической экзальтации с повышением возбудимости.
•Изменение возбудимости клеток или ткани под действием постоянного электрического тока называется физиологическим электротоном.Соответственно различают катэлектрон и анэлектрон (изменение возбудимости под катодом и анодом).
Закон физиологического электротона:
•действие постоянного тока на ткань сопровождается изменением ее возбудимости. При прохождении постоянного тока через нерв или мышцу порог раздражения под катодом и соседних с ним участках понижается вследствие деполяризации мембраны -возбудимость повышается. В области приложения анода происходит повышение порога раздражения, т. е. снижение возбудимости вследствие гиперполяризации мембраны. Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение возбудимости). Повышение возбудимости под катодом называется катэлектротоном, а снижение возбудимости под анодом -анэлектротоном.
•При дальнейшем действии постоянного тока первоначальное повышение возбудимости под катодом сменяется ее понижением, развивается так называемая катодическая депрессия. Первоначальное же снижение возбудимости под анодом сменяется ее повышением -анодная экзальтация. При этом в области приложения катода происходит инактивация натриевых каналов, а в области действия анода происходит снижение калиевой проницаемости и ослабление исходной инактивации натриевой проницаемости.
Постоянный электрический токизменяет
•электрический заряд плазматической мембраны,
•ее ионную проницаемость и
•функциональные свойства (возбудимость, проводимость и т.д.)
Помимо физического электрона (сдвиги электрического заряда и ионной проницаемости мембраны) выделяют:
•Физиологический электротон–изменения функциональных свойств мембраны при прохождении через нее постоянного электрического тока.
•Физиологический электротон проявляется по-разному (под анодом (+) заряд тока и катодом (-) заряд).
Различают
•А) катэлектротон –повышение функциональных свойств мембраны под катодом;
•Б) анэлектротон –понижение функциональных свойств мембраны под анодом (в медицине используется для электросна).
•Периэлектротон
•-волнообразные изменения электрического и функционального состояния плазматической мембраны, создаваемые ПД (или электрическими раздражителем ткани) на участках мембраны, удаленных от места ее возбуждения.
•Изучен недостаточно. Предполагается, что периэлектротон является механизмом безимпульсной, неспецифической передачи информации. Проблема представляет большой интерес для науки и практики.
Физиологическое действие периодического тока
•Организм в целом, а также различные его ткани представляют собой сложный электролитный раствор. Под влиянием постоянного электрического поля, возникающего между двумя электродами, ионы, заключенные внутри ткани, приходят в движение:
•Положительно заряженные –к катоду;
•отрицательно заряженные –к аноду.
В клетках изменяется
•ионное соотношение –происходит разделение ионов,
•скопление их у полупроницаемых мембран по силовым линиям электрического поля,
•возникает так называемая поляризация.
•В клетке происходит разрыхление оболочки, и
•увеличивается ее проницаемость.
•Изменяются биофизические свойства, что способствует повышению возбудимости клетки и тканей.
•Энергичнее протекает обмен веществ.
•Неповрежденная кожа обладает большим сопротивлением для постоянного тока. Начальным местом воздействия гальванического тока на организм является кожа и заложенные в ней нервные рецепторы.
•Действие гальванизации вызывает не только местную реакцию на коже.
Основным механизмом действия гальванического токаявляется рефлекс, возникающий вследствие раздражений током заложенных в коже и в более глубоких тканях нервных рецепторов. Раздражение передается в ЦНС, в том числе через вегетативный отдел, на различные органы и системы, и проявляются изменением их кровенаполнения, обменом веществ, трофики.
•Установлено, что ответные реакции на раздражение нервных рецепторов могут протекать в пределах как тех сегментов, кожа которых подвергается воздействию гальваническим током, так и более отдаленных с участием подкорковых и корковых центров.
•Физиологическое действие электрофорезаразличных лекарственных ионов складывается из действия самого гальванического тока и фармакологического действия вводимых с его помощью ионов.
•В коже под электродом, прокладка которого смочена лекарственными веществами, образуется скопление ионов. Из такого «депо» лекарственные ионы очень медленно и равномерно поступают в общий кровоток и достаточно медленно выводятся из организма.
•Под влиянием самого тока повышается чувствительность организма к вводимым лекарственным веществам.
№15 см №14 Дополнительно: Действие постоянного тока…
Постоянный ток на ткань оказывает два вида действия: 1. Возбуждающее действие
2. Электротоническое действие.
Возбуждающее действие сформулировано в трех законах Пфлюгера:
1. При действии постоянного тока на ткань возбуждение возникает только в момент замыкания цепи или в момент размыкания цепи, или при резком изменении силы тока.
2. Возбуждение возникает при замыкании под катодом, а при размыкании - под анодом.
3. Порог катодзамыкательного действия меньше, чем порог анодразмыкательного действия.
Разберем эти законы:
1. Возбуждение возникает при замыкании и размыкании или при сильном действии тока, потому что именно эти процессы создают необходимые условия для возникновения деполяризации мембран под электродами.
2. Под катодом, замыкая цепь, мы по существу вносим мощный отрицательный заряд на наружную поверхность мембраны. Это приводит к развитию процесса деполяризации мембраны под катодом.
Поэтому именно под катодом возникает процесс возбуждения при замыкании.
Рассмотрим клетку под анодом. При замыкании цепи происходит внесение мощного положительного заряда на поверхность мембраны, что приводит к гиперполяризации мембраны. Поэтому под анодом никакого возбуждения нет. Под действием тока развивается аккомодация. КУД смещается вслед за потенциалом мембраны, но в меньшей степени. Возбудимость снижается. Нет условий для возбуждения
Разомкнем цепь - потенциал мембраны быстро вернется к исходному уровню.
КУД быстро меняться не может, он будет возвращаться постепенно и быстро меняющийся потенциал мембраны достигнет КУД - возникнет возбуждение. В этом главная причина того, что возбуждение возникает в момент размыкания.
В момент размыкания под катодом КУД медленно возвращается к исходному уровню, а потенциал мембраны это делает быстро.
1. Под катодом при длительном действии постоянного тока на ткань возникнет явление - катодическая депрессия.
2. Под анодом в момент замыкания возникнет анодный блок.
Главным признаком катодической депрессии и анодного блока является снижение возбудимости и проводимости до нулевого уровня. Однако, биологическая ткань при этом остается живой.
Электротоническое действие постоянного тока на ткань.
Под электротоническим действием понимают такое действие постоянного тока на ткань, которое приводит к изменению физических и физиологических свойств ткани. В связи с эти различают два вида электротона:
1. Физический электротон.
2. Физиологический электротон.
Под физическим электротоном понимают изменение физических свойств мембраны, возникающее под действием постоянного тока - изменение проницаемости мембраны, критического уровня деполяризации.
Под физиологическим электротоном понимают изменение физиологических свойств ткани. А именно - возбудимости, проводимости под действием электротока.
Кроме того, электротон разделяют на анэлектротон и катэлектротон.
Анэлектротон - изменения физических и физиологических свойств тканей под действием анода.
Каэлектротон - изменения физических и физиологических свойств тканей под действием катода.
Изменится проницаемость мембраны и это будет выражаться в гиперполяризации мембраны и под действием анода будет постепенно снижаться КУД.
Кроме того, под анодом при действии постоянного электрического тока развивается физиологический компонент электротона. Значит под действием анода изменяется возбудимость. Как изменяется возбудимость под действием анода? Включили электроток - КУД смещается вниз, мембрана гиперполяризовалась, резко сместился уровень потенциала покоя.
Разница меджду КУДом и потенциалом покоя увеличивается в начале действия электрического тока под анодом. Значит возбудимость под анодом в начале будет снижаться. Потенциал мембраны будет медленно смещаться вниз, а КУД - достаточно сильно. Это приведет к восстановлению возбудимости до исходного уровня, а при длительном действии постоянного тока под анодом возбудимость вырастет, так как разница между новым уровнем КУДа и потенциалом мембраны будет меньше, чем в покое.
№16Физиологические свойства мышц
1. Возбудимость – это способность мышцы отвечать на раздражение возбуждением;
2. Проводимость – это способность проводить возбуждение вдоль всего мышечного волокна;
3. Сократимость – это способность изменять длину или напряжение при возбуждении;
4. Эластичность – это способность мышцы после сокращения принимать первоначальную форму;
Дополнительно: Физиологические свойства и функции поперечно-полосатых (скелетных) мышц…
Поперечно-полосатая мускулатура составляет основу скелетной мускулатуры. Она обладает двумя важнейшими функциями: 1.Функция движения.
2.Функция поддержания позы (позно-тоническая функция).
Поперечно-полосатая мускулатура обладает тремя главными физиологическими свойствами, а именно - возбудимостью, проводимостью и сократимостью.
Возбудимость скелетных мышц ниже, чем у нервов, и больше (выше), чем у клеток паренхиматозных органов. Возбудимость скелетных мышц значительно выше, чем у гладкой мускулатуры.
Проводимость. Скорость проведения возбуждения в мышцах, ниже, чем в нервах и больше, чем у паренхиматозных тканей. У скелетных мышц проводимость больше, чем у гладких.
Сократимость - это способность мышцы уменьшать свою длину или/и увеличивать свое напряжение. Сокращение - это процесс. Процесс сокращения может выражаться в изменении длины (укорочение мышцы), изменении напряжения мышцы, в изменении того и другого показателя.
Все мышечные сокращения могут быть:
1. изотонические сокращения - это такие сокращения, когда напряжение (тонус) мышц не изменяется ("изо" - равные), а меняется только длина сокращения (мышечное волокно укорачивается).
2. изометрические - при неизменной длине меняется только напряжение мышц.
3. ауксотонические - смешанные сокращения (это сокращения, в которых присутствует и один и другой компонент).
Фазы мышечного сокращения:
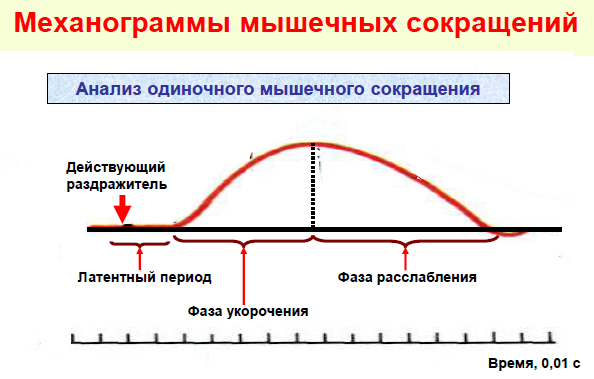
1. Латентный период - это время от нанесения раздражения до появления видимого ответа. Время латентного периода тратится на:
а) возникновение возбуждения в мышце;
б) распространение возбуждения по мышце;
в) электромеханическое сопряжение (на процесс взаимосвязи возбуждения с сокращением); г) преодоление вязко-эластических свойств мышц.
2. Фаза сокращения выражается в укорочении мышцы или в изменении напряжения, либо и в том и в другом.
3. Фаза расслабления - возвратное удлинение мышцы, или уменьшение возникшего напряжения или то и другое вместе.
4. Фаза остаточных колебаний
С позиций фаз все сокращения делятся на: фазные, нефазные
Фазные сокращения - это те сокращения, в которых четко выделяют все три фазы. Нефазные сокращения - это такие сокращения, в которых какая- либо из фаз смазана, отсутствует, растянута на неопределенное время.
Фазные сокращения.
К ним относятся: одиночное мышечное сокращение, тетанус
Одиночное мышечное сокращение:
1. латентный период 2. фаза сокращения 3. фаза расслабления
На скелетных мышцах одиночное мышечное сокращение может возникнуть только в экспериментальных условиях (в искусственно созданных условиях). В естественных условиях скелетные мышцы никогда не ответят одиночным мышечным сокращением. Потому что к ним импульсы в естественных условиях приходят группами. Однако одиночное мышечное сокращение лежит в основе всех других видов мышечных сокращений.
Тетанус - это длительное суммированное фазное сокращение.
1.всегда суммированные одиночные мышечные сокращения.
2.всегда фазное сокращение (можно выделить все три фазы).
Механизм формирования тетануса. В основе формироваия тетанического сокращения лежит процесс суммации. По мнению Гельмгольца, крупнейшего немецкого физиолога, в основе тетануса лежит суммация мышечных сокращений. По мнению Введенского, первично наблюдается суммация возбуждения, а вторично - суммация мышечных сокращений. Если в момент расслабления мышцы, когда она находится в фазе относительной рефрактерности, нанести повторное раздражение, то виден эффект суммации - одно мышечное сокращение наслоится на другое. Если нанести через какой-то интервал времени еще одно раздражение, то снова виден эффект суммации. И каждый раз новая суммация на серию импульсов будет начинаться с нового уровня.
Зубчатый тетанус возникает тогда, когда импульс приходит в фазу расслабления. Но бывает, что приходящий импульс застает мышцу на пике сокращения и тогда возникает полная суммация амплитуды. При такой частоте возникает сокращение - гладкий тетанус (возрастание амплитуды). Для одного и того же объекта в одном и том же физиологическом состоянии большая частота будет давать гладкий тетанус, меньшая - зубчатый, совсем редкая частота - одиночное мышечное сокращение. Если уменьшить частоту, то в какой-то момент гладкий тетанус перейдет в зубчатый. Для каждой конкретной мышцы своя частота получения гладкого и зубчатого тетануса. От функционального состояния мышцы (от ее лабильности) зависит, какой будет тетанус - гладкий или зубчатый. Если вы на определенной частоте получили зубчатый тетанус и продолжаете раздражать долго, то получится ли гладкий тетанус? И если продолжать долго раздражать, то перейдет ли гладкий тетанус в зубчатый?
Нефазные мышечные сокращения
1. Тонус - это длительное, суммированное, постоянно существующее у мышцы напряжение мышечных волокон. Т.е. тонус у живой мышцы существует всегда. В нем нет начала и нет конца. Поэтому тонус относят к нефазным мышечным сокращениям. Это признак того, что мышечный объект живой. Выраженность его может меняться. В нем нельзя выделить фазы.
2. Контрактура - это длительное, суммированное мышечное сокращение с растянутым периодом расслабления.
№17 Режимы сокращения мышц
Для скелетной мышцы характерны два основных режима сокращения - изометрический и изотонический. Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, мышца пытается поднять большой груз) - она не укорачивается. Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способную поднять данный груз, а потом мышца укорачивается - меняет свою длину, сохраняя напряжение, равное весу поднимаемого груза. Так как изотоническое сокращение не является " чисто" изотоническим (элементы изометрического сокращения имеют место в самом начале сокращения мышцы), а изометрическое сокращение тоже не является " чисто" изотоническим (элементы смещения все-таки есть, несомненно), то предложено употреблять термин " ауксотоническое сокращение" - смешанное по характеру.
Виды сокращений
У скелетной мышцы выделяют одиночное сокращение и суммированное сокращение, или тетанус. Одиночное сокращение - это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы. После короткого скрытого периода (латентный период) начинается процесс сокращения. При регистрации сократительной активности в изометрических условиях (два конца неподвижно закреплены) в первую фазу происходит нарастание напряжения (силы), а во вторую - ее падение до исходной величины. Соответственно эти фазы называют фазой напряжения и фазой расслабления. При регистрации сократительной активности в изотоническом режиме (например, в условиях обычной миографической записи) эти фазы будут называться соответственно фазой укорочения и фазой удлинения. В среднем сократительный цикл длится около 200 мс (мышцы лягушки) или 30-80 мс (у теплокровных). Если на мышцу действует серия прямых раздражении (минуя нерв) или непрямых раздражении (через нерв), но с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания 2-й фазы, то мышца будет на каждый из этих раздражителей отвечать одиночным сокращением.
Суммированные сокращения возникают в том случае, если на мышцу наносятся 2 и более раздражения, причем всякое последующее раздражение (после предыдущего) наносится либо во время 2-й фазы (расслабления или удлинения), либо во время 1-й фазы (укорочения или напряжения).
В случае, когда всякое второе раздражение попадает в период фазы расслабления (удлинения), возникает частичная суммация - сокращение еще полностью не закончилось, а уже возникло новое. Если подается много раздражителей с подобным интервалом, то возникает явление зубчатого тетануса. Если раздражители наносятся с меньшим интервалом и каждое последующее раздражение попадает в фазу укорочения, то возникает так называемый гладкий тетанус.
Одиночное сокращение и его фазы.
•Различают 2 основных вида мышечных сокращений:
•одиночные и
•тетанические.
•Механический ответ мышечного волокна или отдельной мышцы на однократные их раздражения нервным импульсом или кратким толчком тока, называется одиночным сокращением.
•При этом в миоплазме мышцы происходит кратковременный подъем концентарации Ca++ вн., сопровождаемый кратковременной работой -тягой мостиков, сменяющейся снова покоем. В изометрическом режиме одиночное напряжение начинается через 2 мсек. после пика ПДм, причем напряжению предшествует краткое и незначительное, так называемое латентное расслабление, создаваемое выходом Ca++ из саркоплазматического ретикулума, что приводит к потере ретикулумом тургора.
•В естественных условиях мышечные волокна двигательной единицы и скелетная мышца в целом работают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон.
№18 Тетанус и его виды
•Суммация сокращений и тетанус возникают в том случае, если на мышцу наносится не одно, а несколько раздражений с определенным интервалом времени.
•Рассмотрим пример:
•Если интервал между раздражениями будет больше длительности одиночного сокращения (больше 100 мсек.), то зарегиструются 2 одиночных сокращения. Уменьшая интервал между раздражениями от 100 до 50 мсек
•( частота 10-20 Гц), т.е. второе раздражение будет наноситься в фазу расслабления и амплитуда сокращения мышцы на второе раздражение будет больше, т.к. второе сокращение возникает тогда, когда мышца еще не успела полностью расслабиться, т.е. происходит суммация сокращений. При этом на миограмме регистрируется две вершины.
•Если интервал между раздражениями меньше 50 мсек
•(частота более 20 ГЦ), то второе раздражение воздействует на мышцу в конечный период фазы сокращения и произойдет полное слияние двух сокращений. Однако это будет наблюдаться до тех пор, пока интервал между раздражениями превышает длительность возникающего перед сокращением ПД, т.к. во время его развития резко падает возбудимость (фаза абсолютной рефрактерности.) и на второе раздражение, следующее с интервалом менее 5 мсек.
•(частота более 200 Гц) -не реагирует.
Зубчатый тетанус
•Развивается на ряд последовательных раздражений, интервал между которыми больше, чем длительность фазы сокращения мышцы.
•Гладкий тетанус
•Возникает тогда, когда интервал между раздражениями меньше длительности фазы сокращения, но больше чем продолжительность потенциала действия.
•У разных мышц длительность фаз одиночного сокращения неодинакова, а, следовательно, частота стимуляции или естественного возбуждения мышцы для получения тетанического сокращения различны.
•Тетанус характеризуется слиянием одиночных мышечных сокращений в непрерывное укорочение мышцы, которое по амплитуде превышает уровень, достигаемый при одиночном сокращении.
•В случае гладкого тетануса после нескольких первых импульсов последующие ответы мышечных волокон не изменяют достигнутого напряжения, а лишь поддерживают его. В подобном режиме двигательные единицы мышц человека работают при развитии максимальных изометрических усилий. При гладком тетанусе развиваемое двигательными единицами напряжение в 2-4 раза больше, чем при одиночных сокращениях.
•В режиме тетанического сокращения мышца способна работать лишь короткое время. Это объясняется тем, что из-за отсутствия периода расслабления она не может восстановить свой энергетический потенциал и работает как бы « в долг».
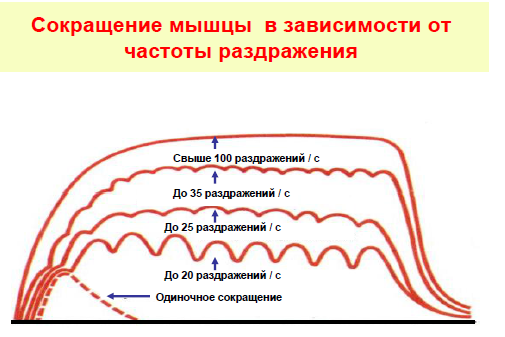
В естественных условиях мышечные волокна,двигательные единицы и скелетная мышца в целомработают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон.
•Так, режим одиночного сокращения медленных волокон камбаловидной мышцы человека обеспечивается при частоте импульсации мотонейрона менее 12 импульсов в секунду, а быстрых волокон глазодвигательных мышц –при скорости импульсации мотонейрона менее 50 импульсов в секунду.
•Гладкий тетанус для быстрых и медленных мышц достигается при разных частотах импульсации мотонейронов. Зависит это от времени одиночного сокращения. Так, гладкий тетанус для быстрой глазодвигательной мышцы проявляется при частоте свыше 150-200 импульсов в секунду, а у медленной камбаловидной мышцы –при частоте около 30 импульсов в секунду.
Оптимум и пессимум ( Введенский)
•Высокие частоты раздражения, вызывающие уменьшение сокращения мышцы, Введенский назвал пессимальными частотами раздражения, а ответ мышцы на это раздражение -пессимальным сокращениям. Уменьшение частоты стимуляции тотчас приводит к восстановлению высокого уровня тетанического сокращения.
•Чтобы разбираемая ситуация имела конкретные числовые выражения, что облегчит понимание, условно определим интервалы временной продолжительности
•Рассмотрим фазы: аболютная рефрактерность=5 мсек.
•относительная рефрактерность от 5-10 мсек.
•супернормальная. возбудимость от 10 до 40 мсек
•Отсюда можно рассчитать частоту ритма раздражения, при котором импульсы раздражающего тока будут совпадать с той или иной фазой возбудимости.
•Расчет показывает, что
•при раздражении с частотой ритма 200 импульсов/сек многие импульсы тока будут действовать на ткань, когда она находится в состоянии абсолютной рефрактерности и не способна отвечать на них и, следовательно, они будут неэффективными, а ритм раздражения подвергается трансформации в более медленный ритм возбуждения. Если подобное возникает при исследовании сократительной способности мышц, то в этих условиях степень сокращения мышц будет меньше, чем при раздражении тока меньшей частоты
При увеличении интервала между раздражениями от 5 до 10 мсек.( от 200 до 100 импульсов в секунду), каждое следующее раздражение будет наноситься в фазу относительной рефрактерности. В этом частотном интервале выявляется наивысшая частота, которая способна воспроизводиться исследуемой структурой без искажения ритма, что соответствует лабильности данной структуры.
•Последующее увеличение интервала между раздражениями от 10 до 40 мсек.( частоты от 100 до 25 импульсов в секунду) создает условия, когда каждое раздражение приходится на фазу супернормальной возбудимости, что благоприятствует возникновению возбуждения, и при этом можно ожидать максимального сокращения мышцы. Частота, которая вызывает максимальный сократительный эффект, была названа Введенским оптимальной частотой раздражений, а сокращение -оптимальным.
•Изменяя силу раздражений при их фиксированной частоте, Введенский показал, что оптимум и пессимум сокращения зависят и от силы раздражения.
№19 Парабиоз
•Экспериментальные факты, составляющие основу учения о парабиозе, Н.В. Введенский (1901) изложил в своем классическом труде «Возбуждение, торможение и наркоз».
•При изучении парабиоза, так же как и при исследовании лабильности, опыты проводились на нервно-мышечном препарате.
•Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется.
Нервно-мышечный препарат помещался во влажную камеру, а на его нерв накладывались три пары электродов для нанесения раздражений и отведения биопотенциалов. Кроме этого, в опытах регистрировались сокращение мышцы и потенциала нерва между интактным и альтерированным участками. Если же участок между раздражающими электродами и мышцей подвергнуть действию наркотических веществ и продолжать раздражать нерв, то ответ на раздражение через некоторое время внезапно исчезает. Н.Е. Введенский, исследуя в подобных условиях действие наркотиков и прослушивая с помощью телефона биотоки нерва ниже наркотизированного участка, заметил, что ритм раздражения начинает трансформироваться за некоторое время до того, как полностью исчезнет ответ мышцы на раздражение. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы:
•уравнительную,
•парадоксальную и
•тормозную,
которые характеризуются разной степенью возбудимости и проводимости при нанесении на нерв слабых (редких), умеренных и сильных (частых) раздражений.
Если наркотическое вещество продолжает действовать после развития тормозной фазы, то в нерве могут произойти необратимые изменения, и он погибает.
•Если же действие наркотика прекратить, то нерв медленно восстанавливает свою исходную возбудимость и проводимость, а процесс восстановления проходит через развитие парадоксальной фазы
В состоянии парабиоза происходит снижение возбудимости и лабильности.
•Учение Н.Е.Введенского о парабиозе носит универсальный характер, т.к. закономерности реагирования, выявленные при исследовании нервно-мышечного препарата, присущи целому организму. Парабиоз есть форма приспособительных реакций живых образований на разнообразные воздействия и учение о парабиозе широко используется для объяснения различных механизмов реагирования не только клеток, тканей, органов, но и целого организма.
Дополнительно: Парабиоз - означает "около жизни". Он возникает при действии на нервы парабиотических раздражителей (аммиак, кислота, жирорастворители, КCl и т.д.), этот раздражитель меняет лабильность, снижает ее. Причем снижает ее фазно, постепенно.
Фазы парабиоза:
1. Сначала наблюдается уравнительная фаза парабиоза. Обычно сильный раздражитель дает сильный ответ, а меньший - меньший. Здесь наблюдаются одинаково слабые ответы на различные по силе раздражители( Демонстрация графика).
2. Вторая фаза - парадоксальная фаза парабиоза. Сильный раздражитель дает слабый ответ, слабый - сильный ответ.
3. Третья фаза - тормозная фаза парабиоза. И на слабый и на сильный раздражитель ответа нет. Это связано с изменением лабильности.
Первая и вторая фаза - обратимые, т.е. при прекращении действия парабиотического агента ткань восстанавливается до нормального состояния, до исходного уровня.
Третья фаза - не обратимая, тормозная фаза через короткий промежуток времени переходит в гибель ткани.
Механизмы возникновения парабиотических фаз
1. Развитие парабиоза обусловлено тем, что под действием повреждающего фактора происходит снижение лабильности, функциональной подвижности. Это лежит в основе ответов, которые называют фазы парабиоза.
2. В нормальном состоянии ткань подчиняется закону силы раздражения. Чем больше сила раздражения, тем больше ответ. Существует раздражитель, который вызывает максимальный ответ. И эту величину обозначают как оптимум частоты и силы раздражения.
Если эту частоту или силу раздражителя превысить, то ответная реакция снижается. Это явление - пессимум частоты или силы раздражения.
3. Величина оптимума совпадает с величиной лабильности. Т.к. лабильность - это максимальная способность ткани, максимально большой ответ ткани. Если лабильность меняется, то величины, на которых вместо оптимума развивается пессимум, сдвигаются. Если изменить лабильность ткани, то та частота, которая вызывала оптимум ответа, теперь будет вызывать пессимум.
Биологическое значение парабиоза
Открытие Введенским парабиоза на нервно-мышечном препарате в лабораторных условиях имело колоссальные последствия для медицины:
1. Показал, что явление смерти не мгновенно, существует переходный период между жизнью и смертью.
2. Этот переход осуществляется пофазно.
3. Первая и вторая фазы обратимы, а третья не обратимая.
Эти открытия привели в медицине к понятиям - клиническая смерть, биологическая смерть.
Клиническая смерть - это обратимое состояние.
Биологическая смерть - необратимое состояние.
Как только сформировалось понятие "клиническая смерть", то появилась новая наука - реаниматология ("ре" - возвратный предлог, "анима" - жизнь).
№20 Сила и работа мышц. Динамометрия. Эргография. Закон средних нагрузок
•Сила, развиваемая мышцей или пучком мышечных волокон, является суммой сил отдельных волокон. Чем толще мышца и больше ее «физиологическая S»поперечного сечения (физиологическая сумма Sпоперечного сечения отдельных волокон), тем сильнее мышца. Так, например, при мышечной гипертрофии сила и толщина мышцы возрастают в одинаковой степени.
•Мышечная сила зависит не только от активирующего влияния со стороны ЦНС, но также определяется внешними механическими условиями, при которых работает мышца.
•Итак, сила мышцы оценивается весом груза, который она при максимальном возбуждении способна удерживать не меняя своей длины.
Сила мышцы зависит от:
•суммы сил мышечных волокон ( их сократительной способности)
•количества мышечных волокон в мышце и количества функциональных единиц, одновременно возбуждающихся при развитии напряжения
•исходной длины мышцы (предварительно растянутая мышца
•развивает
•большую силу)
•характера регулятивных явлений
•условий взаимодействия мышцы с костями скелета
•В организме человека скелетные мышцы передают силу костям скелета через посредство эластических, обладающих растяжимостью структур-сухожилий. Во время развития силы возникает тенденция к укорочению мышцы и, следовательно, к растяжению и развитию напряжения эластических структур, прикрепляющих мышцы к костям.
•Мышечное сокращение, когда длина мышцы уменьшается с увеличением ее силы, наз. ауксотоническим сокращением. Максимальная ауксотоническая сила гораздо меньше, чем сила сокращения, развиваемого мышцей при постоянной длине, то есть при изометрическом сокращении.
Сила изометрического сокращения и длина мышцы
•В состоянии расслабления мышца, удерживаемая при «длине покоя» путем фиксации обоих ее концов, не развивает силу, которая передавалась бы на держатель. Однако, если один конец потянуть чтобы волокна растянулись в мышце развивается пассивное напряжение.
•Таким образом, покоящаяся мышца эластична, в отличие от резиновой полоски ее напряжение не возрастает линейно при напряжении.
•Отложив измеряемую силу в зависимости от длины в прямоугольной системе координат, получим график длина-напряжениедля покоящейся мышцы, т.е. кривую напряжения покоя.
Кривая нарастает тем круче, чем больше степень растяжения мышцы.
•Следовательно, модуль эластичности покоящейся мышцы возрастает с растяжением.Эластичность свойственна главным образом растяжимым структурам, которые располагаются параллельно по отношению к сократительным миофибриллам, к ним относятся
•сарколемма, окружающая мышечное волокно,
•продольная система саркоплазматического ретикулума,
•элементы соединительной ткани между волокнами.
•В отличие от этих структур миофибриллы в расслабленном состоянии практически не оказывают сопротивления растяжению, актиновые и миозиновые нити не связаны поперечными мостиками и могут легко передвигаться друг относительно друга.
•Степень предварительного растяжения определяет не только величину пассивного эластического напряжения покоящейся мышцы, но и величину дополнительной силы, которую может развивать мышца в случае ее активации при данной исходной длине. Изометрический прирост силы во время сокращения суммируется с пассивным напряжением покоящейся мышцы. Пассивные эластические силы растянутых продольных трубочек и сакролеммы суммируются с активными сократительными силами миофибрилл.
•Сократительная способность мышцы характеризуется ее абсолютной силой(сила, приходящаяся на 1 см 2 поперечного сечения мышечных волокон).
•Показатель рассчитывается так:
•Силу мышцы
•Sее физиологического поперечника (т.е. сумму Sвсех мышечных волокон мышцы).
•У перистых мышц физиологический поперечник больше, чем у IIволокнистых, и поэтому их сила больше.
•Абсолютная сила мышц (в кг на 1 см) у человека:
•Икроножная-6, 24;
•Разгибательная шеи –9.0;
•Жевательная-10,0
•Трехглавая-16,8
•Одиночное мышечное волокно способно развить усилие в 20-30 тонн.
•ЦНС изменяет силу сокращения мышцы путем изменения количества одновременно участвующих в сокращении функциональных единиц, а также частоты поступающих к ним импульсов. Симпатическая НС увеличивает силу сокращений.
Работа мышцы
•Измеряется произведением поднятого груза на величину укорочения мышцы.
•Между грузом, который поднимает мышца и выполняемой ею работой существует зависимость.
•Внешняя работа мышцы равна 0, если мышца сокращается без нагрузки.
•По мере увеличения груза работа вначале увеличивается, а затем постепенно уменьшается.
•При очень большом грузе, который мышца неспособна поднять, работа равна 0.
•Внешняя механическая работа мышцы (А) возможна только в изотоническом режиме работы с грузом при условии, что вес груза меньше общей силы мышцы. В этом случае сократительный аппарат сначала растягивает и напрягает эластический элемент –сухожилие, а затем поднимает подвешенный к нему груз.
•Внешняя механическая работа мышцы максимальна при средних нагрузках. Это называется законом средних нагрузок.
•Наибольшую работу мышца совершает при некоторых средних нагрузках. В данном случае 200-250 г.
•Мощность мышцы, измеряемая величиной работы в единицу времени, также достигает максимальной величины при средних нагрузках.
•Работа мышцы, при которой происходит перемещение, волокна развивают напряжение, но почти не укорачиваются (мышца при этом сокращается в изометрическом режиме) называется статической
Статическая работа более утомительна, чем динамическая
•В процессе двигательной деятельности динамические и статические мышцы сокращения взаимодействуют.
•Например, при более динамическом сокращении мышцы могут эффективно выполнять свою функцию, если статическое напряжение других мышц обеспечивает сохранение покоя.
•Динамометрия–метод измерения мышечной силы. Эргография –метод определения мышечной работоспособности.
•Регистрирует работу группы мышц, совместно выполняющих данные движения. Так, эргограф Моссо записывает движения нагруженного пальца и общего разгибателя всех пальцев руки. Форма кривой утомления и величина произведенной работы широко варьируют у разных людей и зависят от разнообразных факторов внешней среды.
№21 Распространение возбуждения по безмякотным нервным волокнам
•Исследованиями установлено, что в безмякотных волокнах местные токи могут проходить практически через любой участок мембраны, т.к. шванновские клетки не оказывают заметного влияния на электрические свойства их мембран.
•При распространении возб-я местные токи вызывают последовательную непрерывную деполяризацию до критического уровня с последующей генерацией ПД на всѐм протяжении волокна. Такое проведение возбуждения называется непрерывным, при этом местные токи, проходя через значительную Sповерхности мембраны очень быстро затухают. Constдлины «а» и Vраспространения возб-я по сравнению с миелиновыми невелики.
•Скорость распространения возб-я по безмякотным волокнам тем выше, чем больше dволокна.
Распространение возб-я по мякотным нервным волокнам
•Мякотные нервные волокна имеют длину от 1 до 22 мкм и возб-е распространяется со скоростью от 3 до 120 м/с. Осевой цилиндр мякотных волокон имеет миелиновую оболочку, состоящую из шванновских клеток, которая прерывается в перехватах Ранвье. Длина участков волокна между перехватами равна от 0.5 до 2 мм и зависит от длины волокна, возраста и вида организма. У нервных волокон, имеющих больший диаметр, расстояние между перехватами Ранвье больше.
•Миелин обладает свойствами изолятора, поэтому участки волокна, покрытые миелином оказывают сопротивление в 10 и 100 раз большее, чем в перехватах Ранвье.
•Благодаря особенностям своей структуры, миелинизированные нервные волокна проводят ПД очень быстро. Создаются предпосылки для возникновения качественно нового сальтаторного типапроведения возб-я, т.е. при сдвиге МП ток, по существу, не идет через мембрану межперехватных участках, и ПД от одного перехвата Ранвье к соседним перехватам распространяется через межперехватные участки электротонически и почти без декремента.
•Время проведения через межперехватные участки практически = 0 –возбуждение перескакивает от одного перехвата к следующему. Задержка проведения происходит только в перехватах, где электротонический потенциал должен достичь порога и вызвать возбуждение.
Мембрана перехвата специализированадля генерации возбуждения: плотность Na+ каналов здесь ≈ в 100 раз выше, чем в немиелинизированных нервных волокнах =» при сальтаторном распространении возбуждения ПД как бы перескакивают через участки волокна, покрытые миелином, от одного перехвата Ранвье к другому.
•Vзатуханияместных токов у мякотных волокон выражена значительно слабее, чем у безмякотных, т.к. они проходят не через всю поверхность мембраны волокна, а только через участки, ограниченные перехватами Ранвье. Т.е. у мякотных волокон местные токи распространяются на большие расстояния, чем у безмякотных.
•Constдлины и vраспространениявозбуждения у мякотных волокон пропорциональны dволокна, а не √d –как у безмякотных.
•Мякотные волокна имеют сравнительно и более высокий фактор надежности. ПД способны перескакивать не только от перехвата к перехвату, но и через 2-3 и даже 5 перехватов. У них более быстро нарастает и амплитуда ПД.
•Т.о. мякотные волокна и присущее им сальтаторное проведение возбуждения обеспечивает
•не только высокую vраспространения возбуждения, а =» и быстроту реакции, но и
•огромное число каналов информации сенсорных и двигательных процессов при суммарном объеме проводящих путей.
•Кроме того, сальтаторное проведение более экономично.
Лабильность - свойство возбудимой ткани воспроизводить максимальное число потенциалов действия в единицу времени.
Максимальная лабильность - у нервной ткани. Частота раздражений, вызывающая максимальную реакцию называется оптимальной (лат. optimum - наилучший), а вызывающая угнетение реакции - пессимальной (лат. pessimum - наихудший).
*Нервное волокно - до 1000 имп./сек, мышца - 200-250 имп./сек., синапс - до 100-125 имп./сек.
образованиями, обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов осуществляется взаимодействие разнородных по функциям тканей организма, например нервной и мышечной, нервной и секреторной.
№22Структура синапса.
Пресинаптическое окончание аксона нейрона при подходе к иннервируемой клетке теряет миелиновую оболочку, что несколько снижает скорость распространения волны возбуждения. Небольшое утолщение на конце волокна, называемое синоптической бляшкой, содержит синаптические пузырьки размером 20—60 нм с медиатором — веществом, способствующим передаче возбуждения в синапсе.
Синаптическая щель — пространство между пресинаптическим окончанием и участком мембраны эффекторной клетки является непосредственным продолжением межклеточного пространства.
Постсинаптическая мембрана — участок эффекторной клетки, контактирующий с пресинаптической мембраной через синаптическую щель.
Классификация синапсов.
В соответствии с морфологическим принципом синапсы подразделяют на:
• аксо-аксональные (между двумя аксонами);
• аксодендритические (между аксоном одного нейрона и дендритом другого);
• аксосоматические (между аксоном одного нейрона и телом другого);
• дендродендритические (между дендритами двух или нескольких нейронов);
• нервно-мышечные (между аксоном мотонейрона и исчерченным мышечным волокном);
• аксоэпителиальные (между секреторным нервным волокном и грану-лоцитом);
• межнейронные (общее название синапсов между какими-либо элементами двух нейронов).
Все синапсы делят на центральные (в головном и спинном мозге) и периферические (нервно-мышечные, аксоэпителиальные и синапсы вегетативных ганглиев).
В соответствии с нейрохимическим принципом синапсы классифицируют по виду химического вещества — медиатора, с помощью которого происходит возбуждение и торможение эффекторной клетки.
По способу передачи возбуждения синапсы подразделяют на три группы. Первую составляют синапсы с химической природой передачи посредством медиаторов (например, нервно-мышечные); вторую — синапсы с передачей электрического сигнала непосредственно с пре- на постсинаптическую мембрану. Третья группа представлена «смешанными» синапсами, сочетающими элементы как химической, так и электрической передачи.
По конечному физиологическому эффекту, а также по изменению потенциала постсинаптической мембраны, различают возбуждающие и тормозные синапсы.
Механизм проведения возбуждения в синапсах. Передача возбуждения в химическом синапсе — сложный физиологический процесс, протекающий в несколько стадий. Он включает синтез и секрецию медиатора; взаимодействие медиатора с рецепторами постсинаптической мембраны; инактивирование медиатора. В целом синапс осуществляет последовательную трансформацию электрического сигнала, поступающего по нервному волокну, в энергию химических превращений на уровне синаптической щели и постсинаптической мембраны, которая затем снова трансформируется в энергию распространяющегося возбуждения в эффекторной клетке.
Свойства синапсов.
Пластичность синапса.
Одностороннее проведение возбуждения.
Низкая лабильность и высокая утомляемость синапса обусловлены временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности.
Высокая избирательная чувствительность синапса к химическим веществам обусловлена специфичностью хеморецепторов постсинаптической мембраны.
Способность синапса трансформировать возбуждение связана с его низкой функциональной лабильностью и спецификой протекающих в нем химических процессов.
Синаптическая задержка, т.е. время между приходом импульса в преси-наптическое окончание и началом ответа, составляет 1—3 мс. Суммация возбуждений определяется переходом местного возбуждения в распространяющееся в результате временного взаимодействия серии возбуждающих постсинаптических потенциалов.
Трофическая функция синапсов
Нейромедиаторы - физиологически активные вещества, вырабатываемые нервными клетками. С помощью нейромедиаторов нервные импульсы передаются от одного нервного волокна другому волокну или другим клеткам через синаптическую щель.
Нейромодуляторы - химические вещества, которые действуют как нейромедиаторы, но не ограничиваются синаптической щелью, а рассредотачиваются повсюду, модулируя действие многих нейронов в определенной области.
Отличие химических синапсов от электрическиха) наличие относительно широкой синаптической щели (20-80 нм, т. е. 200-800 А)
б) относительно высокое удельное электрическое сопротивление контактирующих мембран
в) морфологическая особенность (пресинаптические везикулы с медиатором)
г) при таких структуре и свойствах синапса пресинаптический ПД не может петлей своего тока возбудить постсинаптическую клетку.
Ток, выходящий через пресинаптическую мембрану, здесь почти целиком уходит через широкую синаптическую щель мимо постсинаптической клетки, обладающей значительным входным сопротивлением .
•А очень небольшая часть этого тока, которая все же входит в постсинаптическую клетку, вызывает лишь ничтожное изменение ее МП.
Суть работы химического синапса
•Пресинаптический ПДздесь работает как инициатор нейро-секреторного акта. При развитии ПД терминали (а также и при искусственной ее деполяризации) в нее из среды входят ионы кальция. А это стимулирует выброс медиатора из 100-200 пресинаптических везикул в синаптическую щель.
•Выход медиатораочень сильно зависит от величины деполяризации терминали. Эта зависимость объясняет резкое снижение выхода медиатора при падении амплитуды ПД терминали. В нормальных условиях в ответ на нервный импульс освобождается около миллиона молекул АХ.
•Медиатор диффундируетк постсинаптической мембране, где для него имеются рецепторы (холинорецепторы-ХР). При взаимодействии АХ и ХР в постсинаптической мембране открываются многочисленные ионные каналы (D∼6,5A). Сопротивление этой мембраны падает в 4000 раз, что приводит к ее частичной деполяризации, т. е. к развитию возбуждающего постсинаптического потенциала (ВПСП). В нервно-мышечном синапсе этот ВПСП называется потенциалом концевой пластинки (ПКП). ВПСП (ПКП), в свою очередь, создает ток, раздражающий соседнюю с постсинаптической злектровозбудимую мембрану мышечного волокна, что и порождает в ней ПД.
Нарушение нервно –мышечной передачи при утомлении
•При длительном раздражении нерва нарушение нервно –мышечной передачи развивается задолго до того, как мышца, а тем более нерв в силу утомления утрачивают способность к проведению возбуждения. Объясняется это тем, что
•в нервных окончаниях при длительном раздражении уменьшается запас «заготовленного» медиатора. Поэтому порции ацетилхолина, выделяющиеся в синапсах в ответ на каждый импульс, уменьшаются и соответственно снижают до подпороговых величин постсинаптические потенциалы.
•При длительном раздражении нерва под влиянием продуктов обмена в мышце происходит постепенное снижение чувствительности постсинаптической мембраны к ацетилхолину.
•В результате величина потенциалов концевой пластинки снижается.
•При снижении амплитуды ПКП ниже некоторого критического уровня возникновение ПД в мышечном волокне прекращается, следовательно, синапсы утомляются быстрее, чем нервные волокна и мышцы.
Механизм передачи возбуждения в синапсах
•В состоянии покоя на постсинаптической мембране можно зарегистрировать периодически возникающие слабые процессы деполяризации, т.н. «миниатюрные потенциалы». Напряжение их приблиз. 0,5 мВ. Их происхождение связывают с периодическим опорожнением медиаторов из отдельных везикул пресинаптической области на постсинаптическую мембрану.
•Мембраны везикул обладают протеолитической активностью. При соприкосновении с пресинаптической мембраной, происходит лизис везикулярной и пресинаптической мембран и содержимое везикулы изливается на постсинаптическую мембрану, вызывая ее деполяризацию, типа местного потенциала.
•Медиатор изменяет проницаемость пор постсинаптической мембраны. Ионы Na+ устремляются с поверхности через постсинаптическую мембрану внутрь протоплазмы клеток постсинаптической ткани. Ионы K+ , наоборот, устремляются на поверхность постсинаптической мембраны.
Механизм передачи возбуждения в синапсах
•При поступлении к пресинаптической области распространяющегося процесса возбуждения пресинаптические везикулы начинают массивно концентрироваться около пресинаптической мембраны. При этом происходит ее массивный лизис и большое кол-во медиатора поступает на постсинаптическую мембрану.
•В случае, если везикулы содержат возбуждающий медиатор, это приводит к обширной деполяризации постсинаптической мембраны, суммации на ней местных потенциалов и возникновению распространяющегося спайкового процесса возбуждения. На постсинаптической мембране регистрируются ВПСП.
•В случае, если пресинаптические везикулы синапса содержат тормозной медиатор , н-р ГАМК, при ее поступлении к постсинаптической мембране происходит гиперполяризация и регистрируется ТПСП.
•Механизмы химической теории возбуждения синапсов заложены русским физиологом А.Ф. Самойловым и америк. исследователем, лауреатом Нобелевской премии, Дж. Экклсом.
Природа постсинаптического возбуждения
•При поступлении возбуждающего медиатора, н-р. ацетилхолина к постсинаптической мембране активируются ее Na+ -K+ каналы.
•При этом ионы Na+ устремляются внутрь клетки постсинаптической области а K+ -наружу. Происходит деполяризация мембраны.
•Ацетилхолин активно разрушается в постсинаптической области холинэстеразой с образованием холина и уксусной кислоты.
•В результате этого процесс возбуждения прекращается и возникает снова только при поступлении следующей достаточной порции АХ. Т.е. возбуждение постсинаптической мембраны носит дискретный, квантовый характер.
•В возбуждениипостсинаптической мембраны принимают участие и др. выделяющиеся в пресинаптическую область вещества: Ca+ , олигопептиды, вещ.П, факторы роста нервов, витамины.
В тормозных синапсах
•поступающий на постсинаптическую мембрану тормозной медиатор блокирует Na+ -K+ каналы и создает процесс гиперполяризации мембраны.
•В последние годы химическая теория возбуждения синаптической области значительно расширилась. В 70-е годы П.К.Анохин впервые указал на ограниченность мембранной теории возбуждения и призвал исследовать постсинаптические процессы возбуждения.
В настоящее время постсинаптические процессы представлены следующим образом:
•медиаторы на постсинаптических мембранах активируютспециальные белковые молекулы –рецепторы. Эти рецепторы в свою очередь активируют специальные G-белки, которые возбуждают ферменты аденилатциклазу, гуанилатциклазу и фосфолипазу C.
•Аденилатциклаза и гуанилатциклаза активируют циклические АМФ и ГМФ. Будучи ферментами, цАМФ и цГМФ через соответствующие протеинкиназы фосфорилируют белки.
•Наряду с этим функцию вторичных передатчиков играют диацетилглицерин, инозитолы и Ca++.
Процесс передачи возбуждения с нервного волокна на скелетные мышечные представлен схемойнервный импульс =>поступление Ca++ внутрь нервного волокна =>освобождение из окончания АХ =>взаимодействие АХ с холинорецептором =>активация хемовозбудимых каналов постсинаптической мембраны в результате взаимодействия медиатора с холинорецепторами =>возникновение ПКП =>критическая деполяризация околосинаптической электровозбудимой мембраны =>генерация ПД. Функциональные свойства синапсов
определяются особенностями их строения и механизмом проведения возбуждения.
•1) Одностороннее проведение через синапс.
Это свойство определяется особенностями химического механизма проведения возбуждения через синапс (поскольку везикулы с медиатором располагаются в пресинаптической области, поступающий в пресинаптическую щель медиатор действует только в одном направлении –деполяризации постсинаптической мембраны).
•2) Синаптическая задержка.
Возбуждение в синапсах задерживается от 1 до 3 мсек. Синаптическая задержка определяется перемещением пресинаптических везикул к пресинаптической мембране, процессами лизиса пресинаптической мембраны, поступлением медиатора в синаптическую щель и действием его на проницаемость каналов постсинаптической мембраны.
•3) Суммация возбуждения в синапсе.
Определяется процессами перехода местного процесса возбуждения на постсинаптической мембране в спайковый распространяющийся ПД.