

Состав пищи шпрота в Черном море, % по весу
Компоненты пищи |
Годы. |
|||||
1957- 1976 |
1981- 1985 |
2001 |
2002 |
2004 |
2005 |
|
Calanus helgolandicus (Claus) |
47,6 |
43,9 |
27,8 |
24,4 |
42,5 |
70,5 |
"Pseudocalanus elongatus (Boeck) |
8,9 |
19,0 |
3,4 |
2,9 |
1,1 |
15,7 |
Paracalanus parvus (Claus) |
5,9 |
5,5 |
2,7 |
0,6 |
1,4 |
1,1 |
Acartia clausi (Giesbr) |
17,9 |
18,7 |
30,1 |
40,9 |
33,5 |
9,4 |
Centropoges kroyeri (Kroyer) |
- |
0,6 |
<0,1 |
0,4 |
- |
- |
Всего Calanoida |
80,3 |
87,7 |
64,0 |
69,2 |
78,5 |
96,7 |
Oithona similis (Claus) |
<0,1 |
<0,01 |
0,1 |
1,7 |
0,4 |
0,2 |
Oithona nana (Giesbr) |
<0,1 |
<0,01 |
- |
- |
- |
- |
Всего Cyclopoida |
<0,1 |
<0,01 |
0,1 |
1,7 |
0,4 |
0,2 |
Harpacticoidae |
<0,01 |
0,3 |
0,5 |
2,5 |
0,3 |
- |
Всего Copepoda |
80,3 |
88,0 |
64,6 |
73,4 |
79,2 |
96,9 |
Podon leuckarti (Sars) |
0,2 |
- |
18,5 |
5,7 |
0,9 |
<0,1 |
Evaohna spinifera (Miiller) |
<0,1 |
- |
- |
- |
- |
- |
Penilia avirostris (Dana) |
0,6 |
0,8 |
<0,1 |
2,0 |
з,з |
- |
Всего Cladocera |
0,8 |
0,8 |
18,5 |
7,7 |
4,2 |
<0,1 |
N. Cirripedia |
<0,1 |
- |
1,0 |
1,3 |
0,1 |
- |
Cipris cirripedia (Latreille) |
0,2 |
0,2 |
- |
- |
- |
- |
Всего Cirripedia |
0,2 |
0,2 |
1.0 |
1,3 |
0,1 |
- |
Oikopleura dioica |
- |
2,8 |
1,3 |
3,6 |
15,5 |
1,1 |
L. Lamellibranchiata |
2,0 |
0,2 |
11,4 |
11,7 |
0,3 |
2,0 |
L. Ostracoda |
- |
<0,1 |
1,2 |
2,3 |
0,4 |
- |
Sagitta euxina (Moltshanov) |
13,9 |
4,8 |
- |
- |
- |
- |
Многощетинковые черви |
2,1 |
0,1 |
0,4 |
- |
- |
<0,01 |
Креветки (личинки) |
0,1 |
0,3 |
0,1 |
<0,1 |
0,3 |
<0,1 |
Рыба (личинки) |
0,1 |
<0,1 |
- |
- |
- |
<0,1 |
Ихтиопланктон (икра) |
<0,1 |
0,3 |
- |
<0,1 |
- |
- |
Фитопланктон (Coscinodiscus) |
<0,1 |
<0,1 |
- |
<0,1 |
- |
<0,1 |
Насекомые |
<0,01 |
0,3 |
- |
<0,01 |
- |
- |
Было установлено, что наблюдаемые изменения являлись особенностью летнего питания шпрота (в период образования температурного скачка) и отмечались в северо-западной и северо-восточной частях моря уже с 1992,1993 it. Так, в начале 90-х гг. прошлого столетия в рационе шпрота отсутствовала S. euxina и в 2 раза уменьшилась доля Р elonqatus. С 2000 по 2003 г. в летнем рационе шпрота практически не встречались копеподы С. helqolandicus и P. elonqatus (рис.3 ).
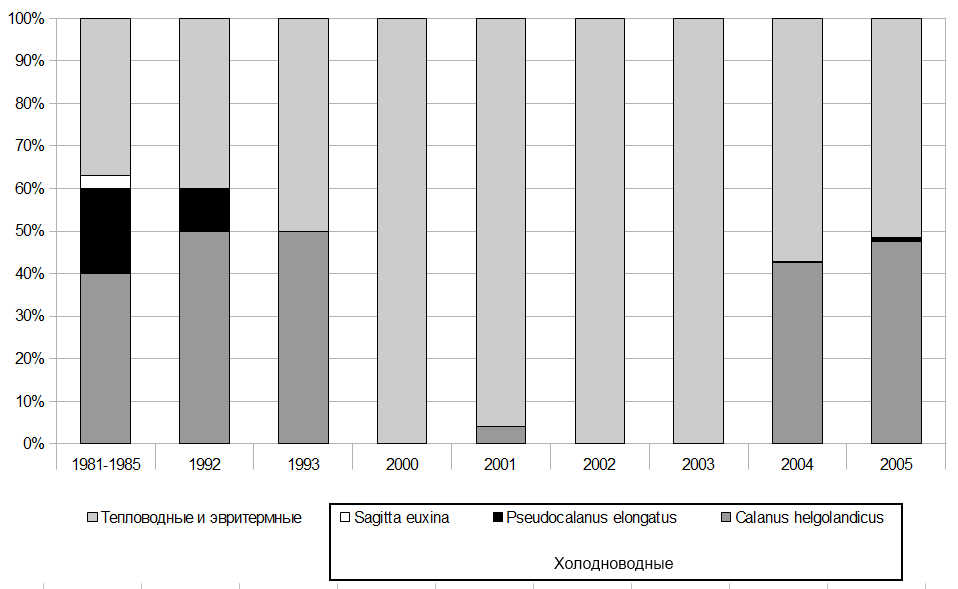
Рисунок 3. Доля холодноводных и тепловодных организмов в пище шпрота в северо-западном и северо-восточном районах Черного моря в летний период, в % от массы пищевого комка
Ухудшение условий нагула в 2002-2003 гг. позволило выявить четкие различия в питании крупного, размерами более 86 мм, и мелкого, размерами 60-85 мм, шпрота. Эти различия сводились к тому, что крупные особи, образующие скопления под слоем термоклина, вместо традиционной и энергетически ценной пищи - копепод С. helqolandicus - питались различными представителями подвижного бентоса (L. ostracoda, L. polychaeta, L. lamellibranchiata,- Oikopleura dioica). Менее ограниченный в вертикальном распределении мелкий шпрот дефицит основного корма компенсировал за счет тепловодных организмов (Глущенко Т.И.).
В осенне-зимний период, в условиях гомотермии, в пищевом спектре крупного и мелкого шпрота различий не установлено. Шпрот всех размерных групп питался взрослыми формами копепод С. helqolandicus, P. elongatus и A. clausi (табл. 8).
Таблица 8