

Контрольні запитання
Дайте визначення коефіцієнтів динамічної та кінематичної в’язкості.
Наведіть температурну залежність коефіцієнта динамічної в’язкості.
Запишіть закон Пуазейля для нетурбулентної течії рідини.
Наведіть класифікацію рідин за їх реологічними властивостями.
Наведіть формули для середнього часу релаксації, енергії активації молекули, осередненої початкової швидкості стрибка молекули.
Лабораторна робота 4 зміна властивостей рідких кристалів і біологічних мембран при нагріванні
Мета та основні завдання роботи: визначення температури за допомогою рідких кристалів.
Обладнання, прилади і матеріали: термометр, пробірки, еталонна і досліджувана рідиннокристалічні плівки, нагрівач, штатив.
Основні теоретичні відомості
Клітини всіх живих організмів, а також клітинні органели еукарiот оточені мембранами – звідси ясно, яка велика роль цих утворень у живій природі. Будова всіх мембран у своїй основі однакова, проте їхній склад сильно змінюється залежно від походження, навіть у межах одного організму.
Склад мембран. Біологічні мембрани складаються з білків, ліпідів, води і неорганічних компонентів. Відносний вміст цих складових частин і їх природа можуть істотно відрізнятися (наприклад, мембрани Azotobacter agilis і Agrobacterium tumefaciens на 100% складаються з фосфоліпідів). Фосфоліпідний склад мембран також різний. Ця різниця стосується як вуглеводних ланцюгів, так і полярних головок.
Майже в такій же кількості, як і фосфоліпіди, у мембранах може знаходитися холестерин. Однією з перших, достатньо детальних моделей мембран є модель, побудована в 1935 р. Даніеллі і Девсоном. Ці автори припустили, що ліпіди утворюють подвійний шар, у якому полярні головки спрямовані у бік води, а жирні (гідрофобні) ланцюжки занурені в товщу мембрани (рис. 5.1). Ця модель була потім удосконалена, але її суть залишилася незмінною.
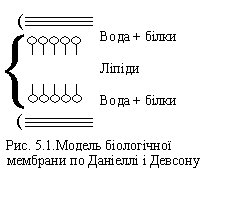
Мембрани мають таку текучість, що дозволяє їм функціонувати при розподілі клітин, виконувати метаболічні функції і т.д. Щоб зрозуміти як функціонують мембрани, необхідно розглянути взаємодію між ліпідами і білками і властивості різних сумішей цих з'єднань. Оскільки важливим компонентом мембран є фосфоліпіди, зупинимося на них докладніше. Фізичні властивості безводних фосфоліпідів дуже складні через термотропний поліморфізм. Сполуки з довгими вуглеводними ланцюжками часто утворюють декілька кристалічних форм. Можна згадати моделі, у яких припускається, що ланцюги розташовані перпендикулярно, або похило стосовно площини, утвореної полярними головками.
Важливу роль у визначенні властивостей водно-ліпідних систем відіграє висока полярність води. При температурах нижче критичної вода не може проникати в ліпідну фазу. Коли ланцюжки плавляться, молекули води можуть дифундувати до полярних головок, чому сприяє зменшення взаємодії між ланцюжками внаслідок інтенсифікації теплового руху. Отже, Ткр залежить від природи ланцюжків, природи полярних головок, кількості води і, крім того, від наявності в розчині заряджених частинок.
М
49
Розподіл фосфоліпідів неоднорідний. Фосфоліпіди, що містять холіни, розташовуються в основному з зовнішнього боку мембрани, а в яких є аміногрупи – з внутрішнього. Це призводить до того, що білки переважно розміщуються з внутрішнього боку мембрани.
Часто мембранні білки мають форму – спіралі (до 40%). Такі білки можуть бути занурені в мембрану, а іноді навіть пронизують її так, що модель, у якій ліпіди і білки утворюють послідовні прошарки, не відповідає дійсності.
Ліпіди розміщені таким чином, що утворюються подвійні прошарки, так що чисто міцелярні моделі також неадекватні.
Нарешті, мембрани повинні бути достатньо текучими, але ця їхня властивість не повинна сильно залежати від температури, що може забезпечуватися зміною ступеня насиченості вуглеводних ланцюжків.
Розподіл білків – нерівномірний. Відповідно до електронно-мікроскопічних даних, концентрація їх на внутрішній поверхні вища, нiж на зовнішнiй. Використання різноманітних міток показало, що бiлки досить рухливі. У мембранах може бути присутнім у тій або іншій кількості холестерин. Системи лецитін - холестерин – вода вивчені дуже докладно. Показано, що холестерин утворює із лецитіном міцні асоціати, тому що площа, що припадає на полярну головку при приєднанні холестерину, сильно зменшується.
Властивості мембран. Якщо мембрана розділяє два водяних розчини хлорида лужного металу різної концентрації, то між цими розчинами виникає різниця потенціалів.
Цю різницю можна обчислити за формулою Е = 0,058 lg(а1/а2), де а1 і а2 – активності (приблизно рівні концентраціям, якщо розчин слабкий) розчинів по різні боки мембрани; поверхня, що межує з більш розведеним розчином, заряджена позитивно, тобто відштовхує катіони.
Мембрана поводиться як випрямляч: якщо до неї прикласти різницю потенціалів V, що постійно змінюється, і виміряти силу струму I, що виникає при цьому, то отримана крива I = f(V) виявиться несиметричною стосовно вісі абсцис (рис. 5.2). Чим менше гiдратовані катіони (частіше усього тi, що мають великий іонний радіус), тим легше вони проходять крізь мембрану (рис. 5.3).
Органічні іони селективно виштовхують неорганічні. Проникність мембрани для різних катіонів різнa: мембрана пермоселективна. Якщо з одного її боку знаходиться NaCl, а з іншого КСl такої ж концентрації, то поверхня, що межує з розчином NaCl, заряджається позитивно, що свідчить про більшу проникність мембрани для іонів калію, ніж натрію.
Е лектропровідність
мембрани зазвичай
низька і залежить від природи іонів,
при цьому внесок катіонів дуже малий
порівняно з внеском аніонів. Відзначимо,
що адсорбовані іони рухливі і вносять
вклад у провідність. Важливу біологічну
роль може відігравати і рух іонів уздовж
поверхні мембрани: наприклад, рухливість
іонів на поверхні ракових клітин
відрізняється від такої для нормальних
клітин.
лектропровідність
мембрани зазвичай
низька і залежить від природи іонів,
при цьому внесок катіонів дуже малий
порівняно з внеском аніонів. Відзначимо,
що адсорбовані іони рухливі і вносять
вклад у провідність. Важливу біологічну
роль може відігравати і рух іонів уздовж
поверхні мембрани: наприклад, рухливість
іонів на поверхні ракових клітин
відрізняється від такої для нормальних
клітин.
П
51
Проникність мембран залежить від природи утворюючих їх прошарків: прошарки з малою проникністю утворюються прямими насиченими жирно-кислотними ланцюгами, а також ланцюгами спиртів і інших сполук, що містять велику кiлькiсть атомів вуглецю (С16 і вище).
У даному гомологічному ряду проникність пов'язана з довжиною ланцюга і залежить від розміру полярної головки. Найбільшу проникність мають прошарки, утворені з ненасиченних жирних кислот (наприклад, олеїнової), холестерину і білків.
У мономолекулярному прошарку можуть існувати пари, що виконують визначені функції; їх положення не обов'язково фіксоване. Можна визначити енергію активації проникності Е, припустивши, що кількість молекул, що проходять через мембрану, пропорційна ехр(-E/k); тільки молекули, чия енергія перевищує Е, перетинають мембрану.
Експериментально встановлено, що між двома боками клітинної мембрани існує різниця потенціалів (потенціал спокою), що складає 50 – 80 мВ, при цьому середовище заряджене стосовно внутрішньої поверхні мембрани позитивно. Така полярність підтримується спеціальним механізмом, локалізованим у мембрані і який потребує енергію (іонний насос).
В
54![]() ),
і ця різниця потенціалів призводить до
виникнення електричного поля дуже
високої напруженості (104 – 105
В см-1).
У цих умовах молекули води поблизу
порівняно сильно гідратованого іона,
такого, як натрій, знаходяться одночасно
під впливом мембранного поля і поля,
утворюваного іоном, у результаті чого
вони намагаються відокремитися від
іона і Na+
стає менше сольватованим, у граничному
випадку взагалі позбавленим гідратованої
води. Оскільки радіус іона Na+
менший, нiж радіус менш гідратованого
іона К+,
вільні іони натрію з більшою легкістю
проходять крiзь мембрану, нiж іони калію
(це встановлено за допомогою дослідів
із використанням ізотопів). Описаний
процес зветься активним транспортом і
відрізняється від пасивного, спонтанного
транспорту, що відбувається за рахунок
простої дифузії іонів.
),
і ця різниця потенціалів призводить до
виникнення електричного поля дуже
високої напруженості (104 – 105
В см-1).
У цих умовах молекули води поблизу
порівняно сильно гідратованого іона,
такого, як натрій, знаходяться одночасно
під впливом мембранного поля і поля,
утворюваного іоном, у результаті чого
вони намагаються відокремитися від
іона і Na+
стає менше сольватованим, у граничному
випадку взагалі позбавленим гідратованої
води. Оскільки радіус іона Na+
менший, нiж радіус менш гідратованого
іона К+,
вільні іони натрію з більшою легкістю
проходять крiзь мембрану, нiж іони калію
(це встановлено за допомогою дослідів
із використанням ізотопів). Описаний
процес зветься активним транспортом і
відрізняється від пасивного, спонтанного
транспорту, що відбувається за рахунок
простої дифузії іонів.