


Медична

Друге видання, виправлене
ЗАТВЕРДЖЕНО
МОЗ України як підручник для студентів вищих медичних навчальних закладів
І-ІІІ рівнів акредитації
київ
ВСВ «МЕДИЦИНА» 2012
УДК
61 ;57 ББК 5я73 Б24
директор Львівського державного медичного коледжу ім. Андрея Крупинсь- кого, доктор медичних наук, професор М. Б. Шегедин;
завідувач кафедри фармакогнозії з медичною ботанікою Тернопільського державного медичного університету ім. 1.Я. Горбачевського, доктор фармацевтичних наук, професор С.М. Марчишин.
Барціховський В.В.
П.Я.
Шерстюк. — 16 с. кольор. вкл.
е вид. випр. — К.: ВСВ “Медицина”, 2012. — 312 с. + 1БВЫ 978-617-505-215-0
Матеріал підручника викладено відповідно до навчальної програми з медичної біологи. У підручнику відображено молекулярно-генетичний, клітинний та онтогенетичний рівні організації життя, ураховуючи специфіку організму людини; подано основи генетики людини, біосферний рівень організації життя і місце людини в ньому. Особливу увагу приділено спадковим хворобам і медико-біологічним аспектам паразитизму.
Важливого значення надано формуванню практичних умінь і навичок розв’язування типових задач, запропоновано широкий спектр контрольних завдань для самоперевірки знань.
Додатковим мотиваційним чинником до опанування знань виступає актуалізація кожної теми, висвітлення п медичної, наукової та соціальної значущості. Зв’язок із практикою, принцип історизму забезпечують блоки науково-популярної інформації, які за своїм змістом доповнюють і розширюють теми, шо вивчаються.
Для студентів вищих медичних навчальних закладів І—III рівнів акредитації.
УДК 61;57 ББК 5я73
ISBN
978-617-505-215-0
РОЗДІЛ І
ПРЕДМЕТ МЕДИЧНОЇ БІОЛОГІЇ. СТРУКТУРНО-ФУНКЦІОНАЛЬНА ОРГАНІЗАЦІЯ КЛІТИНИ. РОЗМНОЖЕННЯ НА КЛІТИННОМУ РІВНІ
Чому це актуально?
Медична біологія — інтегральна наука, яка комплексно аналізує основи життєдіяльності людини і відкриває перспективи для її благополуччя.
Особливості життєвих процесів визначаються закономірностями існування клітин, тому практично всі хвороби пов'язані з порушенням структури і функцій клітин, із яких утворюються тканини та органи.
Патологія клітин зумовлюється багатьма різноманітними чинниками, знання яких є ключовими для розуміння механізмів виникнення захворювань та умов збереження здоров'я людини.
Предмет медичної біології
Медична біологія — наука, що вивчає спадковість людини, її генетичну систему, генотипові й індивідуальні відмінності людей, їх екологію, фізіологію, особливості поведінки.
Завдання медичної біологи:
вивчення патологічних явищ на внутрішньоклітинному, тканинному, органному і системному рівнях;
поглиблення знань щодо структурних, функціональних, генетичних особливостей людського організму;
систематизація спадкових хвороб, реалізація програми «Геном людини»;
вивчення етіологічних чинників генних, хромосомних і геномних мутацій;
розкриття взаємодії біологічних і соціальних чинників у виникненні патології людини;
захист геному людини від шкідливих антропогенних впливів різного походження;
застосування методів клітинної інженерії і досягнень щодо використання генів і клітин рідкісних тварин і рослин, створення відповідних банків генетичного матеріалу, що дасть змогу зберегти унікальний живий світ нашої планети.
З
Характеристики життя, рівні організації живого
Життя — це біологічна форма руху матерії — найбільш складна форма всесвіту, яка існує на космічному тілі — планеті Земля.
Стратегією життя є:
стійка здатність до передачі інформації та а реалізації;
адаптація до умов навколишнього середовища;
поступальний розвиток.
Основні властивості життя:
обмін речовин та енергії;
подразливість;
самооновлення;
саморегуляція;
самовідтворення;
спадковість і мінливість;
ріст і розвиток;
дискретність та цілісність.
Виділяють такі рівні організації живої матерії (рис. 1.1, див. кольорову вклейку):
Молекулярно-генетичний рівень. Елементарними молекулярними структурами є коди спадкової інформації, які відтворюються в ході реплікагп ДНК та реалізуються внаслідок трансляції.
Екологічні проблеми рівня: зростання мутагенних впливів і збільшення частки мутацій у генофондах.
Клітинний рівень. Елементарними структурами є клітини, які об’єднуються у тканини і системи органів. Елементарними явищами є життєві цикли клітин.
Екологічні проблеми рівня: ріст клітинної патологи внаслідок забруднення середовища, порушення відтворення клітин.
Організмовий рівень. Елементарні структури — організми та системи органів, з яких вони складаються, шо володіють комплексом фізіологічних процесів, які забезпечують життєдіяльність.
Екологічні проблеми рівня: зниження адаптаційних можливостей організмів, розвиток граничних станів у людини (стан між здоров'ям і хворобою).
Популяційно-видовий рівень. Елементарні структури — популяції з характерним генофондом як основні одиниці еволюті. Елементарні явиша — видоутворення на підставі природного добору.
Екологічні проблеми рівня: погіршення екологічних показників популяції (чисельність, щільність, віковий склад тошо).
Бюсферно-біогеоценотичний рівень. Елементарні структури — біогеоценози, які характеризуються динамічним взаємозв’язком у масштабах біосфери.
Екологічні проблеми рівня: збільшення кількості антропоценозів та їх глобальне поширення, забруднення середовища, руйнування озонового екрана Землі.
Хімічний склад клітини. Органічні та неорганічні сполуки
Жива клітина містить обмежений набір хімічних елементів, причому шість із них становлять понад 99 % її загальної маси: С, Н, Т4, О, Р, 5. З атомів цих елементів утворені практично всі молекули клітин різних організмів.
\
/ о.
5-
}
Водневий
зв’язок
\
/ о.
8-Частковий
заряд
'5-
■н
н*
о. .
Рис. 1.2. Будова молекули води
Усі хімічні елементи, які входять до складу клітин, можна поділити на чотири групи:
органогенні — це оксиген, гідроген, карбон і нітроген, їх загальний вміст становить 95—98 %;
макроелементи — кальцій, калій, фосфор, сульфур, силіцій, натрій, хлор, магній, ферум, які містяться в десятих частках відсотка;
мікроелементи — кобальт, цинк, куп- рум, манган, хром, бром, бор, йод, літій, радій, їх вміст становить близько 0,01 %;
ультрамікроелементи — усі інші хімічні елементи, вміст яких менше 0,01 %.
Функції хімічних елементів:
входять до складу органічних і неорганічних сполук клітини;
активують діяльність низки ферментів;
створюють різницю потенціалів на біомембранах тощо.
Серед неорганічних сполук важлива роль належить воді, її вміст у биіьшості клітин становить 60—70 %, а в деяких клітинах може сягати 90 % маси клітини.
Унікальні властивості води полягають у полярному характері її молекул, завдяки чому дві молекули води можуть притягуватися за рахунок сил електростатичної взаємодії між частково негативним зарядом на атомі оксигену однієї молекули та позитивним зарядом на атомі гідрогену іншої. Такий тип зв’язку називається водневим (рис. 1.2).
Функції води у клітині:
добрий розчинник (розчинні речовини — гідрофільні, нерозчинні — гідрофобні);
визначає фізичні властивості клітини — об’єм, внутрішньоклітинний тиск (тургор);
є середовищем для хімічних реакцій;
забезпечує в клітині терморегуляцію, зокрема за рахунок високої теплоємності, яка визначається здатністю молекул утворювати асоціати.
Органічні сполуки. Клітини значною мірою складаються з білків, на які припадає більше половини маси сухої речовини клітини. Білки є полімерами (поліпептидами), оскільки побудовані з 20 різних амінокислот, що здатні зв’язуватися між собою за допомогою пептидних зв ’язків. Комбінації з 20 амінокислот можуть утворювати незліченну кількість різних за структурою і функціями білків.
Прості білки утворені тільки амінокислотами. До складу складних білків можуть входити ліпіди (ліпопротеїди), вуглеводи (глікопротеіди). Білки мають кілька рівнів структурної організації (рис. 1.3).
Функції білків:
каталітична — ферменти в тисячі разів пришвидшують хімічні процеси, забезпечують усі основні функції клітини;
« г
Рис.
1.3.
Структури
білкової молекули: а
— первинна структура; б
— вторинна структура; в
— третинна структура; г
— четвертинна структура
регуляторна — гормони білкової природи беруть участь у регуляції і координації багатьох метаболічних і фізіологічних процесів;
структурна — білки забезпечують утворення всіх елементів клітин і організмів: органел, мембран, тканин, органів;
захисна — речовини білкової природи захищають організм, формують його захисні покриви, оболонки органів і клітин, утворюють антитіла;
рецепторна — сигнальні білки розпізнають характер та інтенсивність діючих подразників зовнішнього середовища;
транспортна — полягає у перенесенні речовин до місця використання;
рухова — забезпечується м’язовими білками: актином, міозином та ін.;
енергетична — пов’язана з можливістю використання молекул білків для отримання енергії.
Нуклеїнові кислоти — унікальні молекули (полінуклеотиди), необхідні кожній клітині для збереження і передачі генетичної інформації. Вони забезпечують процеси синтезу білків, а отже, визначають характер обміну речовин, закономірності росту і розвитку, явища спадковості та мінливості. Нуклеїнові кислоти мають такі різновиди:
ДНК
НУКЛЕЇНОВІ
КИСЛОТИ

Вуглеводи — органічні речовини, що мають загальну формулу Сх(Н20)у, де X та У можуть мати різні значення. їх поділяють на три основні класи: моносахариди, дисахариди, полісахариди.
Моносахариди (глюкоза, фруктоза, галактоза, рибоза, дезоксирибоза) — джерело енергіГ'для різних метаболічних процесів:
С, Н О + 602 -> 6С02 + 6Н20 + енергія (АТФ).
Дисахариди (мальтоза, сахароза, лактоза) — є також джерелом енергії для клітин.
Полісахариди (глікоген, крохмаль) — запасають велику кількість енергії, деякі зв'язуються з білками й утворюють глікопротеїни.
Ііпіди — гідрофобні речовини, що є складними естерами жирних кислот і гліцерину. Слугують джерелом енергії, виконують захисну функцію (низький коефіцієнт теплопровідності), беруть участь у побудові клітинних мембран, забезпечуючи їх напівпроникність.
Структурно-функціональна організація еукаріотичної клітини
Клітина складається з великої кількості чітко упорядкованих різноманітних молекул. Молекулярні комплекси утворюють органели, які є складовою частиною клітинної системи. Внутрішній простір клітини поділено мембранами на відсіки (ком- партменти), де відбуваються тільки специфічні для цього простору реакції. Таким чином, клітина є складною системою макромолекул кількох рівнів організації. Це цілісна неподільна система, в якш можна виділити низку підсистем, відповідальних за специфічні функції: мембрани (перетинки), цитоплазму, ядро, мітохонорії тощо. Клітинні органели структурно і функціонально зв’язані між собою. Життєдіяльність клітин може здійснюватися тільки за умов скоординованого зв’язку між ними.
Будь-яка клітина складається з поверхневого апарату, цитоплазми, органел та інших внутрішньоклітинних структур.
Поверхневий апарат клітини утворюють такі структури:
плазматична мембрана (має ліпопротеїдну природу);
надмембранш структури (у бактерій, рослин і грибів — клітинна стінка, у тварин — глікокалікс);
тОмембранш структури.
Плазматична мембрана — це напівпроникний ліпідний бішар із вбудованими в нього білками (рис. 1.4, див. кольорову вклейку).
Будову і функції структурних компонентів клітини відображено на рис. 1.4— 1.12 (див. кольорову вклейку) і в табл. 1.1.
Таблиця 1.1. Будова і функції структурних комнонентів
Складові частини |
Особливості будови, склад |
Функції |
Підмембранні компоненти |
||
Цитоплазма |
Внутрішнє середовище клітини, міститься між плазматичною мембраною і ядром. Характеризується відносною сталістю будови та властивостей (гомеостаз). Основа (матрикс) цитоплазми — гіалоплазма — перебуває у рідинному стані (золь) або драглистому (гель) і містить білки, ліпіди, нуклеїнові кислоти, солі |
Об’єднує складові частини (компартменти) клітини; забезпечує гліколіз, синтез білка за участю рибосом |
Цитоскелет (рис. 1.5) |
Утворений мікрониточками (актин, міозин), мікротрубочками (тубумн), проміжними філамен- тами (цитокератин); є єдиною системою |
Забезпечує локалізацію структур, форму клітини, транспорт компонентів |
Складові частини |
Особливості будови, склад |
Функції |
Клітинний центр (центросома, 0,1— 0,3 мкм) (рис. 1.12) |
Складається з 1—2 центріолей (циліндричних тілець), оточених центросферою |
Утворює мікротрубочки для внутрішньоклітинного транспорту |
Рибосоми (15—20 мкм) (рис. 1.6, 1.11) |
Тільця округлої форми, утворені нуклеопротеїда- ми, мають дві нерівні субодиниці: малу — для приєднання іРНК та тРНК, велику — для синтезу поліпептидного ланцюга |
Матричний синтез білка |
Включення |
Тимчасові зерна (вуглеводи і білки) та краплі (жири) |
Запасні речовини |
Одномембранні органели (вакуолярна система клітини) |
||
Ендоплазматична сітка, діаметр 25— 30 нм (рис. 1.6) |
Система порожнин у вигляді канальців і цистерн (потовщень канальців), сполучених між собою й оточених мембранами (схожі на зовнішню клітинну мембрану). Залежно від будови є два типи ЕС: гладенька (агранулярна) — без рибосом і шорстка {,гранулярна) — з рибосомами |
Синтез білків (шорстка), ліпідів, вуглеводів (гладенька), розщеплення токсинів, дозрівання і накопичення білків, транспорт речовин у різні ділянки клітини |
Апарат Голь- джі (20-2000 нм) (рис. 1.7) |
Утворений стопкою плоских цистерн (мішечків), вкритих мембранами, поряд з якими розміщені міхурці та канальці |
Накопичення, дозрівання, пакування і виведення (секретування речовин)', формування лізосом |
Лізосоми, діаметр — 1 мкм (рис. 1.8) |
Округлі тільця, утворюються в міхурцях апарату Гольджі. Оточені мембраною, містять гідролітичні ферменти |
Руйнування речовин, ор- ганел, клітин (внутрішньоклітинне травлення), утворення травних вакуолей |
Вакуолі |
У рослин — мішечки, заповнені рідиною; у тварин — скоротливі, травні, фагоцитарні |
Накопичення речовин |
Двомембранні органели |
||
Пластиди (у рослин), 3—10 мкм (рис. 1.9) Види: хлоропласти, хро- мопласти, лейкопласти |
Паличкоподібні, пластинчасті, у вигляді лусок і зерен. У хлоропластів зовнішня мембрана гладенька, внутрішня — утворює грани — пакети плоских мішечків (тилакоїдів). Тилакоїди сусідніх гран сполучаються мембранними каналами {ламели). На поверхні гран знаходиться хлорофіл. У пластидах є ДНК, РНК, включення. Здатні розмножуватися |
Хлоропласти забезпечують фотосинтез — перетворення світлової енергії на енергію хімічних зв’язків |
Мітохондрії (0,5—5 мкм) (рис. 1.10) |
Паличкоподібні, округлі, ниткоподібні; зовнішня мембрана гладенька, внутрішня має багато виростів (крист), які збільшують поверхню. Внутрішній вміст — мітохондріальний матрикс, містить рибосоми, ДНК, ІРНК, тРНК |
Відбуваються окисно- відновні процеси, ВИДІЛЯЄТЬСЯ енергія, що нагромаджується в АТФ. Синтезуються власні білки, РНК, ДНК |
Складові частини |
Особливості будови, склад |
Функції |
Ядро |
||
Поверхневий апарат |
Складається з 2 мембран, які мають пори |
Обмеження ядра, зв’язок з цитоплазмою |
Каріоплазма |
Рідинне середовище ядра |
Місцезнаходження ядерних елементів |
Хроматин |
Речовина, що складається з ДНК, білків, іРНК (інтерфазна форма існування хромосом) |
Носій спадкової інформації |
Рибонуклео- протеїдні комплекси |
Містять ДНК, РНК |
Забезпечують утворення рибосом |
Клітина — відкрита система. Організація потоків речовини та енергії в клітині
Клітина є відкритою системою, тому що вона не ізольована від зовнішнього середовища. Для життя та функціонування клітинам необхідно постійно взаємодіяти з навколишнім середовищем. Зокрема, між середовищем і клітинами постійно відбувається обмін речовиною, енергією й інформацією. Ці процеси забезпечують упорядкований у часі та просторі, координований перебіг усіх метаболічних і фізіологічних процесів.
Обмін речовиною полягає в тому, що між середовищем і клітинами (як відкритими системами) відбувається обмін молекулами. Клітина вибірково поглинає необхідні і виводить непотрібні їй речовини Потік речовин пов’язаний насамперед із метаболізмом клітин, що являє собою єдність асиміляції і дисиміляції.
Асиміляція — процес «уподібнення» речовин, що надходять у клітину, специфічним речовинам, які характерні для неї. Це — ендотермічний процес, тобто процес, шо потребує витрати енергії. Установлено, що синтез речовин у клітинах відбувається за рахунок метаболічного фонду, який включає: продукти перетравленої їжі та продукти дисиміляції, які утворюються в клітинах. Потік речовин у клітині підтримує стабільний молекулярний склад її цитоплазми, що включає неорганічні й органічні сполуки. У травному тракті їжа перетворюється на суміш низькомолекулярних органічних речовин: амінокислот, нуклеотидів, вуглеводів — неспецифічних і однакових для всієї живої природи. У такому вигляді вони і надходять у клітину та утворюють метаболічний фонд. Унаслідок процесів асиміляції виконуються дві дуже важливі функції:
підтримання структури клітини;
забезпечення енергією, що надходить з органічними речовинами.
Дисиміляція — екзотермічний процес, тобто процес вивільнення енергії внаслідок розпаду речовин клітини. Речовини, що утворюються при дисиміляції, також підлягають подальшому перетворенню і використовуються клітиною.
Особливості транспорту речовин через мембрану пов’язані з тим, що ліпідний бішар практично непроникний для більшості полярних водорозчинних молекул, оскільки внутрішня частина його гідрофобна. Такий бар’єр запобігає втраті водорозчинного вмісту клітини. Різні речовини мають неоднакову здатність проникати через цей бар’єр.
Великі макромолекули (білки, жири) та їх агрегати не можуть проникати крізь мембрану. Для перенесення існує «макромеханізм» — захоплення клітиною і перенесення в певному напрямку (ендоцитоз і екзопитоз).
Ендоцитоз — складний активний процес поглинання клітиною великих молекул, часток, мікроорганізмів. Різновиди: пінопитоз, фагоцитоз, опосередкований рецепторами ендоцитоз.
Піноцитоз — поглинання рідини та розчинених речовин з утворенням специфічних мембранних пухирців.
Фагоцитоз (рис. 1.13, див. кольорову вклейку) — поглинання твердих часток (мікроорганізмів, часток клітин). При цьому утворюються великі щільні ендоцитоз- ні пухирці — фагосоми, які зливаються з лізосомами і формують фаголізосоми.
Опосередкований рецепторами ендоцитоз характеризується поглинанням із позаклітинної рідини певних макромолекул.
Екзоцитоз — процес виведення макромолекул, унаслідок якого внутрішньоклітинні секреторні пухирці зливаються з плазмолемою і їх вміст виводиться із клітини. Під час ендоцитозу ділянка плазматичної мембрани втягується і замикається, утворюється ендоцитозний пухирець, що містить поглинений матеріал. Під час ек- зоцитозу мембрана секреторних пухирців зливається з плазматичною мембраною і вміст вивільняється в позаклітинний простір.
Невеликі молекули переносяться через мембрану за допомогою спеціальних молекулярних механізмів двох типів: пасивного і активного транспорту.
Пасивний транспорт — переміщення невеликих полярних (С02, Н20) і неполярних (02, ТЧ2) молекул за градієнтом концентрації або електрохімічним градієнтом без витрати енергії. Існують різні форми пасивного транспорту:
проста дифузія газів — під час дихання між порожниною альвеол легень і просвітом кровоносних капілярів, характеризується низькою вибірковістю мембрани до речовин, що переносяться;
полегшена дифузія — за участю компонентів мембрани (канали і переносники) переважно в одному напрямку (у клітину) за градієнтом концентрації без витрат енергії, характеризується вибірковістю до речовин;
осмос — процес дифузії розчинника (Н20) через напівпроникну мембрану за концентраційним градієнтом із високої концентрації розчинника у бік з низькою концентрацією. Клітина має два класи мембранних транспортних білків, що формують наскрізні шляхи через гідрофобний шар: численні білки-переносники й іонні канали. Білки-переносники — це складні глобулярні білки, що мають спорідненість до певних молекул, забезпечують їх перенесення через мембрану. Іонні канали складаються із кількох зв’язаних між собою білкових субодиниць, що формують у мембрані велику пору. Через неї за електрохімічним градієнтом проходять іони.
Активний транспорт — перенесення молекул через мембрану за допомогою спеціальних білків проти концентраційного або електрохімічного градієнта з використанням енергії аденозинтрифосфату (АТФ). Білки-переносники є одночасно ферментами і називаються АТФазами.
Обмін енергією полягає в тому, що у клітину разом з органічними речовинами надходить енергія, акумульована в хімічних зв’язках між молекулами й атомами, яка потім вивільняється і перетворюється на АТФ. Енергія необхідна для підтримання стабільності клітинної системи. Вона забезпечує у клітині її структуру, гомеостаз, метаболізм, функції.
Енергія вивільняється в процесі дисиміляції. Обмін енергією в групах організмів забезпечується різними процесами: гліколізом, фотосинтезом, хемосинтезом, диханням. Для тварин основним є процес дихання. Це сукупність біохімічних реакцій розщеплення (окиснення) органічних речовин (глюкози, жирних кислот, амінокислот) до С02 і Н20 і використання енергії розірваних хімічних зв’язків для утворення висококалорійного клітинного «палива» у вигляді АТФ. Як відомо, енергетичним матеріалом у клітині є молекула АТФ. Вона накопичує енергію, вільно переміщується в клітині й віддає енергію для здійснення ендотермічних процесів. АТФ утворюється в результаті приєднання до аденозиндифосфату (АДФ) третьої фосфатної групи. Процеси утворення АТФ відбуваються на внутрішній мембрані мітохондрій, яка містить спеціальні ферменти — АТФ-синтетази.
Потік енергії — це сукупність реакцій розщеплення органічних речовин з вивільненням енергії хімічних зв’язків, яка акумулюється в макроергічних зв’язках АТФ і надалі використовується клітиною.
Обмін енергією відбувається в декілька етапів: підготовчий, безкисневий і кисневий.
На підготовчому етапі за допомогою травних ферментів великі органічні молекули розщеплюються на мономери: білки — до амінокислот, жири — до гліцерину і жирних кислот, полісахариди — до моносахаридів, нуклеїнові кислоти — до нук- леотидів. Енергія, яка при цьому вивільняється, розсіюється у вигляді тепла.
Безкисневий (анаеробний) етап енергетичного обміну відбувається у клітинах. На цьому етапі молекули глюкози розщеплюються шляхом гліколізу на дві молекули піровиноградної (С3Н403) або (особливо у м’язових клітинах) молочної кислоти (С,Н603). Сумарне рівняння гліколізу має такий вигляд:
С6Н1206 + 2АДФ + 2Н3Р04 = 2С3Н603 + 2АТФ + 2НгО. (1)
Під час гліколізу виділяється близько 200 кДж енергії. Частина її (близько 80 кДж) витрачається на синтез двох молекул АТФ, а решта (приблизно 120 кДж) — розсіюється у вигляді тепла. Незважаючи на низьку ефективність, гліколіз має надзвичайно велике фізіологічне значення. Завдяки йому організм може отримувати енергію в умовах дефіциту кисню, а його кінцеві продукти (піровиноградна та молочна кислоти) зазнають подальшого ферментативного перетворення в аеробних умовах.
Кисневий (аеробний) етап енергетичного обміну можливий лише в аеробних умовах (за наявності кисню), коли органічні сполуки, що утворилися на безкисне- вому етапі, окиснюються в клітинах до кінцевих продуктів — С02 і Н20. Окиснення сполук пов’язане з відщепленням від них гідрогену, який передається за допомогою особливих біологічно активних речовин-переносників до молекулярного кисню, з утворенням молекули води. Цей процес називають тканинним диханням. Він відбувається в мітохондріях і супроводжується виділенням великої кількості енергії та акумуляцією її в макроергічних зв’язках молекул АТФ (окисне фосфори- лювання). Сумарне рівняння аеробного процесу має такий вигляд:
2С3Н603 + 602 + 36Н3Р04 + 36АДФ = 6С02 + 36АТФ + 6Н20. (2)
Таким чином, на кисневому етапі утворюється у 18 разів більше АТФ, ніж на безкисневому.
Універсальною речовиною, яка накопичує енергію в процесі енергетичного обміну, є аденозинтрифосфорна кислота (АТФ). Молекула АТФ — це нуклеотид, який складається із залишків азотистої основи (аденіну), вуглеводу (рибози) і трьох залишків ортофосфатної кислоти. Якшо під дією ферменту відщеплюється один залишок ортофосфатної кислоти, АТФ перетворюється на АДФ з вивільненням близько 40 кДж енергії. Коли від молекули АТФ відщеплюються два залишки ортофосфатної кислоти, утворюється аденозинмонофосфат (АМФ), при цьому вивільняється близько 80 кДж енергії. Молекула АМФ також може розщеплюватися.
Отже, під час розщеплення АТФ виділяється велика кількість енергії, яка використовується для синтезу необхідних організму сполук, підтримання певної температури тіла тощо. Частина енергії, що вивільняється, витрачається на синтез АТФ із АДФ чи АМФ і молекул ортофосфатної кислоти, які зв’язуються макроергічними зв’язками (виникають між залишками ортофосфатної кислоти в молекулах АДФ або АТФ). Таким чином, молекули АТФ є універсальним хімічним акумулятором енергії у клітинах. Хімічна природа макроергічних зв’язків остаточно ще не з’ясована.
Якщо додати рівняння (1) і (2), то одержане сумарне рівняння (3) віддзеркалюватиме енергетичний баланс процесу повного розщеплення глюкози до вуглекислого газу і води:
С6Ніг06 + 602 + 38АДФ + 38Н3Р04 = 6С02 + 38АТФ + 44Н20. (3)
Обмін інформацією полягає в тому, що клітина сприймає зміни в навколишньому середовищі (сигнали) і здатна на них адекватно реагувати. Завдяки цьому клітина може пристосовуватися до мінливих умов існування. Це забезпечується утворенням нових, потрібних у цих умовах ферментів та інших макромолекул. Пристосувальні внутрішньоклітинні процеси призводять до змін форми, розмірів і функціонування клітин. Зрештою адекватна реакція на сигнали зовнішнього середовища дає змогу клітині вижити в умовах, що змінюються.
Певна організація живого пов’язана зі збереженням і використанням потоку інформації для підтримання структурно-функціональної організації клітини та її тривалого стабільного існування як системи. Спадкова інформація зберігається в молекулах ДНК у вигляді генетичного коду — послідовності триплетів нуклеотидів. Інформація переписується з ДНК на молекули РНК, що забезпечують синтез необхідних структурних білків і ферментів. Утворені білки причетні до появи певних властивостей клітини. Іншими словами, потік інформації в клітині спрямований від ДНК до ознаки:
ДНК РНК ->• білок -* ознака.
Ще один інформаційний потік спрямований від ДНК однієї клітини до ДНК дочірньої клітини. Цей потік пов’язаний із процесом розмноження. Він реалізується реплікацією молекул ДНК материнської клітини, утворенням хромосом, процесом рівномірного розподілу спадкового матеріалу між дочірніми клітинами (мітозом): ДНК -» дві дочірні ДНК -> дві дочірні клітини.
Задача 1. Скласти рівняння реакції, що відображає процес: АТФ + 2Н20 -> ?
Розв’язання.
При взаємодії АТФ з однією молекулою води відщеплюється один залишок ортофосфатної кислоти, вивільняється 40 кДж енергії й утворюється молекула АДФ:
АТФ + Н20 = АДФ + Н3Р04+ 40 кДж. (1)
Далі АДФ буде розщеплюватися другою молекулою води до молекул АМФ і Н3Р04 з вивільненням ще 40 кДж енергії:
АДФ + Н20 = АМФ + Н3Р04 + 40 кДж. (2)
Додавши рівняння (1) і (2), матимемо:
АТФ + 2Н20 = АМФ + 2Н3Р04+ 80 кДж.
Задача 2. Знайти загальний енергетичний ефект повного розщеплення 900 г глюкози у клітинах людини.
Розв’язання.
ш
= 900 г
ГЛЮКОЗИ
Е - ?
повн.
розщ
Ураховуючи, що для утворення із одного моля АДФ одного моля АТФ необхідно 40 кДж енергії, розрахуємо загальний енергетичний ефект наведеного процесу:
40 кДж/моль х 38 моль = 1520 кДж.
Перепишемо рівняння (1) у спрощеній формі, враховуючи цей енергетичний ефект:
С6Н1206 + 602 = 6С02 + 44Н20 + 1520 кДж. (2)
Мг (С6НІ206) = 72 + 12 + 96 = 180. М (С6Н1206) = 180 г/моль. Із рівняння (2) бачимо, що у реакцію вступає 1 моль глюкози. Звідси ш (С6Н1206) = М (С6Н1206) х 1 моль (С6Н1206) або
ш (С6НІ206) = 180 г/моль х 1 моль = 180 г.
Складемо пропорцію: якщо розщеплення 180 г глюкози вивільняє 1520 кДж енергії, то розщеплення 900 г глюкози вивільнятиме Еч кДж енергії.
. 900 гх 1520 кДж
Звідси І = 7660 кДж.
180 г
Відповідь. При повному розщепленні 900 г глюкози виділяється 7660 кДж енергії.
Життя клітин поза організмом
Досліджуючи властивості нормальних та пухлинних клітин, учені використовують метод клонування. Кілька десятиліть тому було встановлено, що соматичні клітини еукаріотів можна розмножувати in vitro, тобто підтримувати у вигляді так званих клітинних культур. Розподіл хромосом при розмноженні клітин культури відбувається мітотичним шляхом. Клітинна лінія утворюється з однієї клітини (і, таким чином, є клоном).
Клоном називають групу генетично ідентичних індивідуумів, отриманих шляхом безстатевого розмноження: або один від одного, або від деякого загального предка. Найпростішим прикладом такого клону може бути популяція бактерій, усі клітини якої утворилися внаслідок повторних поділів з однієї батьківської клітини. Усі бактерії у такому клоні успадковують гени цієї батьківської клітини.
Кюнуванням є утворення монозиготних близнят у людини, які виникають з однієї зиготи на ранніх стадіях її дробіння шляхом мітозу. Клонування клітин за-
стосовують у клітинній інженерії, мікробіологічній промисловості для одержання корисних штамів мікроорганізмів.
У лабораторних умовах рослинні й тваринні клітини частіше за все проходять лише обмежену кількість поділів, а потім гинуть. Виняток становлять ракові клітини. Для клітин прокаріотів характерна гаплоїдність, а для еукаріотів — диплоїдність. Це обмежує можливості генетичного аналізу, оскільки рецесивні алелі не виявляються у гетерозиготи.
У зв’язку з експериментами із пересадження клітинних ядер відкрилася можливість для клонування тварин чи людини. Метод клонування використовують також і для ідентифікації генів. Створюють міжвидові гібридні клітини, в яких і визначають розташування генів. Отже, можна дійти висновку, що цей метод принесе багато наукових здобутків на користь людству.
Ядро — центральний інформаційний апарат клітини. Хроматин
Провідна роль в явищах спадковості належить ядру клітини (рис. 1.14, див. кольорову вклейку).
Ядро містить ядерну оболонку, ядерце, каріоплазму, хроматин (хромосоми). Генетичний матеріал (ДНК) в еукаріотів розміщений майже виключно в хромосомах ядра.
Хромосоми — структури ядра клітини, в яких знаходяться гени (ДНК). Хромосоми здатні до самовідтворення. Перед кожним клітинним поділом вони подвоюються, а потім точно порівну розподіляються між дочірніми клітинами.
Головні хімічні компоненти хромосом — ДНК, основні (гістонові) і кислі (негіс- тонові) білки, частки яких складають відповідно 40 і близько 20 %. У невеликій кількості містяться РНК, ліпіди, полісахариди, іони металів. Хромосоми разом із позахромосомними структурами забезпечують збереження, відтворення, реалізацію і передачу генетичної інформації в ряді поколінь.
Хромосоми здатні до спіралізації (конденсації) і деспіралізації (деконденсації). При цьому лінійні розміри (довжина, товщина) хромосом змінюються.
Мітотичні хромосоми (цей стан властивий хромосомам під час мітозу) характеризуються особливостями:
найзручніші для вивчення під час метафази мітозу;
спіралізовані, щільні і компактні;
спостерігаються в світловий мікроскоп у вигляді структур певної форми (ниток, паличок), які інтенсивно забарвлюються основними барвниками;
виконують транспортну функцію: забезпечують точний розподіл і перенесення генетичного матеріалу в дочірні ядра.
Інтерфазні хромосоми:
максимально деспіралізовані;
індивідуально не розрізняються і займають весь об’єм ядра, утворюючи хроматин;
виконують синтетичну функцію: забезпечують синтез ДНК (реплікація) і синтез РНК (транскрипція).
Хроматин — інтерфазна форма існування хромосом. Його видно в світловий мікроскоп як сукупність зерен, грудочок, що добре забарвлюються основними барвниками. Самі хромосоми видно лише під час поділу клітини. Хімічний склад хроматину і хромосом однаковий. Це комплекс ДНК і білків — дезоксирибонуклео- протеїн (ДНП). На ультраструктурному рівні хроматин складається із хромосомних фібрил (ниток) завтовшки 20—25 нм, які, своєю чергою, побудовані з фібрил завтовшки 10 нм. Основою структурної організації фібрил є нуклеосоми.
Хроматин інтерфазного ядра поділяють на еухроматин і гетерохроматин, які різняться між собою за ступенем спіралізації та генетичною активністю.
Еухроматин — деспіралізовані в інтерфазі ділянки хромосом. Це генетично активний хроматин, де розташовані структурні гени і відбувається транскрипція. Еухроматин не забарвлюється барвниками і його не видно в світловий мікроскоп. З початком мітозу весь еухроматин конденсується в складі хромосом і стає видимим.
Гетерохроматин — надміру спіралізовані і сконденсовані ділянки хромосом, які знаходяться в генетично неактивному стані (не транскрибуються). Гетерохроматин поділяють на структурний і факультативний.
Структурний (конститутивний) гетерохроматин — ділянки хромосом, які спіралізовані впродовж усього клітинного циклу (інтерфаза і мітоз). Він постійно займає однакові ділянки в обох гомологічних хромосомах — близько центромер, вторинних перетяжок, на кінцях хромосом (теломерах). Цей хроматин інтенсивно забарвлюється барвниками і його видно в світловий мікроскоп.
Факультативний гетерохроматин локалізується в одній із двох гомологічних хромосом. В окремих випадках одна з них гетерохроматизується повністю. Приклад факультативного хроматину — Х-статевий хроматин, або тільце Барра. Це одна з двох Х-хромосом особин жіночої статі (ссавці, людина), яка гетерохроматизується і переходить у генетично неактивний стан на ранніх стадіях ембріогенезу.
Будову хромосом вивчають у метафазі, коли вони найбільш конденсовані і їх добре видно в світловий мікроскоп. До складу метафазної хромосоми входять:
дві сестринські хроматиди (дочірні хромосоми), які розташовані поруч і з’єднані між собою в ділянці первинної перетяжки (центромери);
центромера — звужена, незабарвлена і найменш спіралізована ділянка хромосоми, містить кінетохор, до якого прикріплюються мікротрубочки веретена поділу;
плечі — ділянки тіла хромосоми, на які її поділяє центромера; кінці плечей називають теломерами. Це ділянки із щільно упакованою ДНК, які перешкоджають з’єднанню хромосом між собою або з їхніми фрагментами (позбавлений теломери кінець хромосоми стає «липким» і легко приєднує фрагменти хромосом).
За положенням центромери розрізняють характерні типи симетрії метафазних хромосом (рис. 1.15).
Метацентричні — рівноплечі хромосоми, в яких центромера знаходиться посередині хромосоми і плечі рівної або майже рівної довжини.
Субметацентричні — нерівноплечі хромосоми, в яких центромера зміщена від середини і одне плече довге, а друге — коротке.
Акроцентричні — хромосоми, в яких центромера розміщена близько до одного з кінців й одне плече довге, а друге — дуже коротке і часто малопомітне.
Телоцентричні хромосоми можуть виникати в результаті втрати одного плеча, у них залишається тільки одне плече з центромерою на кінці. У нормі такі хромосоми не трапляються.
Деякі хромосоми є супутниковими. Вони мають вторинні перетяжки, які відокремлюють тонкою ниткою від основної частини невеликі кінцеві ділянки, які називають супутниками. Вторинні перетяжки можуть знаходитися в прицентромер-
■
о
в
Рис. 1.15. Типи метафазних хромосом людини:
/ — метацентрична; 2 — субметацентрична; 3 — акроцентрична; 4 — акроцентрична із супутником; а — первинна перетяжка (центромера); б — супутник; в — вторинна перетяжка;
д — довге плече; р — коротке плече
них районах довгих плечей (хромосоми 1, 9, 16) і на кінцевих ділянках коротких плечей (хромосоми 13—15 і 21, 22). У зоні вторинних перетяжок деяких хромосом розташовані ядерцеві організатори — місце утворення ядерець.
Хромосомний і геномний рівні організації спадкового матеріалу
Хромосоми залежно від типу клітин можуть мати різні набори.
Диплоїдний (подвійний) набір хромосом властивий ядрам соматичних клітин і позначається 2п. У ньому всі хромосоми парні. Хромосоми однієї пари називають гомологічними, різних пар — негомологічними. Гомологічні хромосоми однакові за розмірами, формою, будовою, але одна з них походить з яйцеклітини (материнська), інша — зі сперматозоїда (батьківська).
Гаплоїдний (одинарний) набір хромосом характерний для ядер статевих клітин. Він удвічі менший і позначається п. У ньому з кожної пари диплоїдного набору міститься лише одна хромосома. Усі хромосоми гаплоїдного набору різні, тобто негомо- логічні. При заплідненні статеві клітини зливаються і в зиготі відновлюється диплоїдний набір хромосом (п + п = 2п). Хромосомні набори соматичних клітин чоловічих і жіночих особин того самого виду різняться між собою за однією парою хромосом.
Каріотип — диплоїдний набір хромосом соматичної клітини, який характеризується сукупністю ознак:
— кількістю хромосом;
розмірами хромосом;
формою хромосом (типом симетрії);
особливостями їхньої будови.
Каріотип є видовою ознакою, йому властиві сталість і специфічність; в особин одного і того ж біологічного виду він точно відтворюється з покоління в покоління завдяки мітозу і мейозу. Тому всі соматичні клітини певного виду несуть повний об’єм генетичної інформації, властивий цьому виду. Каріотипи ж різних видів різняться між собою як за кількістю хромосом, так і їх розмірами, формою та особливостями будови.
Видова специфічність хромосом забезпечується їх здатністю до самовідтворення. Зміни каріотипу можуть відбуватися внаслідок мутацій.
Число хромосом не залежить від рівня організації і розмірів організму. Найбільше число хромосом (1600) мають, наприклад, радіолярії (найпростіші), найменше (2) — аскарида коня.
Статеві хромосоми (гетерохромосоми) — пара хромосом, за якими хромосомні набори соматичних клітин чоловічих і жіночих особин того самого виду різняться між собою. Ці хромосоми визначають стать організму: у жінки — XX, у чоловіка — ХУ.
X — хромосома середня субметацентрична, у ній відомо близько 150 генів (у тому числі рецесивні гени, які зумовлюють дальтонізм (кольорова сліпота), гемофілію (порушення зсідання крові), атрофію зорового нерва).
У — найдрібніша акроцентрична. У ній виявлено небагато генів. У людини У- хромосома відіграє вирішальну роль у формуванні чоловічої статі.
Нестатеві хромосоми (автосоми) — хромосоми, однакові в обох статей, тобто це всі хромосоми, крім статевих.
Нормальний каріотип людини (чоловіка і жінки) складається з 46 хромосом, з них 22 пари автосом і одна пара статевих хромосом. Нормальний каріотип жінки записують 46, XX, чоловіка — 46, ХУ. Нормальний каріотип — важлива умова формування здорової людини. Зміни числа і структури хромосом (мутації) спричинюють хромосомні хвороби.
Вивчення каріотипу (каріотипування) проводять на мікрофотографіях метафаз- них пластинок, рідше при безпосередньому мікроскопуванні.
Для ідентифікації хромосом використовують такі ознаки:
розмір хромосом;
положення центромери, центромерний індекс (відношення довжини короткого плеча до довжини всієї хромосоми, %);
наявність вторинних перетяжок і супутників.
У метацентричних хромосом центромерний індекс становить близько 50 %, у субметацентричних — менше 50 %. Розмір метафазних хромосом варіює від 11 до 2,3 мкм.
Ідіограма (каріограма; рис. 1.16) — розташування пар хромосом у порядку зменшення їх розмірів. Розроблена єдина міжнародна класифікація хромосом людини — денверівська (Денвер, США, 1960 р.), за якою всі пари хромосом пронумеровані й об’єднані в 7 груп — А, В, С, Ц, Е, Ц, в (табл. 1.2). Найбільша пара хромосом має номер 1, наступна — номер 2 і т. д. Статеві хромосоми (XX і ХУ) на ідіограмі розташовують останніми.
? її ** V <?
, Д ,йц -/ її *
«»/ 'У V \ + * ї>л(,,і
*т%' / \ Л*г*
Ч'їд&'Г V* * и^
4^ >
II |
К |
|
К8 |
»і |
№ |
М К |
Ж |
а ні |
1» |
||
] |
2 |
3 |
4 |
5 |
6 |
9 1 1 |
2 |
3 |
4 |
5 |
6 |
м |
д |
м |
и |
М |
И |
и |
м |
д |
Д |
м |
М |
7 |
8 |
9 |
10 |
11 |
12 |
7 |
8 |
9 |
10 |
11 |
12 |
Об |
АА |
ІА |
и |
АХ |
ЛЛ |
ЙЛ |
ІЛ |
м |
п |
ХА |
ДИ |
13 |
14 |
15 |
.16 |
17 |
19 |
13 |
14 |
15 |
іб |
17 |
18 |
хк |
хк |
* А |
А А |
|
п |
XX |
XX |
«А |
«А |
|
Кд |
19 |
20 |
21 |
22 |
|
XX |
19 |
20 |
21 |
22 |
|
ХУ |
Рис. 1.16. Каріотип людини
На рис. 1.16 каріотип жінки — ліворуч, чоловіка — праворуч; угорі — метафаз- ні пластинки, внизу — ідіограми. В ідіограмах хромосоми розташовані попарно в порядку зменшення довжини.
Окремо виділені статеві хромосоми: у жінки — XX, у чоловіка — ХУ.
Таблиця 1.2. Класифікація хромосом людини за розміром і положенням центромери
Групи хромосом |
Номери пар хромосом |
Характеристика хромосом |
А (І) |
1, 2, 3 |
1,3 — великі метацентричні, 2 — велика субметацентрична |
В (II) |
4, 5 |
Великі субметацентричні |
С (III) |
6-12 |
Середні субметацентричні |
в (IV) |
13-15 |
Середні акроцентричні |
Е (V) |
16-18 |
Дрібні субметацентричні |
и (VI) |
19, 20 |
Найдрібніші метацентричні |
С (VII) |
21, 22 |
Найдрібніші акроцентричні |
Х-хромосома (III) |
23 |
Середня субметацентрична |
У-хромосома (VII) |
23 |
Найдрібніша акроцентрична |
Правила хромосом. Для хромосом мають місце певні закономірності їх існування у клітинах, які відомі під правилами хромосом:
Правило постійності числа хромосом. Кожний вид організмів має певне і постійне число хромосом. Так, 2п людини = 46, 2п шимпанзе = 48, 2п дрозофіли =
2п гороху = 14.
Правило парності хромосом. Кожна хромосома в соматичній клітині дип- лоідного організму представлена парою: у людини 23 пари, у шимпанзе — 24, у дрозофіли — 4, у гороху — 7 пар хромосом.
Правило індивідуальності хромосом. Кожна пара хромосом характеризується індивідуальними особливостями і зберігає їх у ряді поколінь.
Правило неперервності хромосом (хромосома від хромосоми). Хромосоми, як і клітини, не виникають заново, а утворюються від уже наявних хромосом шляхом самовідтворення в S-періоді інтерфази (реплікація).
Задача 1. Гаплоїдне число хромосом у людини дорівнює 23. Скільки автосом і гетерохромосом міститься у: а) сперматозоїдах; б) клітинах шкіри; в) яйцеклітинах до запліднення; г) зиготі людини?
Розв’язання:
а) у сперматозоїдах наявний гаплоїдний набір хромосом, тобто 23. З них 1 гетерохромосома (враховуючи гетерогаметність у людини чоловічої статі — це X- або Y-xpo- мосома) і 22 автосоми (23 — 1 = 22);
б) клітини шкіри — соматичні, у них диплощний набір хромосом, тобто 46. Із них 2 гетерохромосоми і 44 автосоми;
в) яйцеклітина належить до гамет, як і сперматозоїд, тому кількісні співвідношення хромосом будуть такі самі, як у сперматозоїді;
г) зигота має диплоїдний набір хромосом, як і клітини шкіри, тому кількості хро
мосом будуть аналогічними.
Відповідь. У сперматозоїдах і яйцеклітинах автосом — 22, гетерохромосом — 1; у
зиготі та клітинах шкіри автосом — 44, гетерохромосом — 2.
Задача 2. Як зрозуміти, що у людини XY-гетерохромосоми визначають чоловічу стать, а у курей — жіночу?
Розв’язання.
X та Y — це лише символи, за допомогою яких розрізняють гетерохромосоми. Статеві ознаки визначаються якісним складом генів, що містяться у хромосомах, а не самими хромосомами. У людини гени, що визначають чоловічу стать, знаходяться в Y- хромосомі, а у курей — у Х-хромосомі.
Життєвий цикл клітини. Мітоз, його порушення
Клітинний цикл (життєвий цикл клітини) — період існування клітини від поділу до поділу або від поділу до смерті. У ньому виділяють інтерфазу та мітоз (рис. 1.17).
Інтерфаза (від лат. inter — між) — проміжок між двома мітозами. Це період підготовки клітини до наступного поділу, коли в ній відбуваються інтенсивні синтетичні процеси, з яких основним є синтез ДНК.
Періоди інтерфази:
— пресинтетичний (G,) — клітина посилено росте, синтезує білки, нагромаджує енергію, збільшує кількість органел, готується до синтезу ДНК; у клітині диплоїдний набір хромосом (2п) і відповідна кількість ДНК (2с); хромосоми — однохрома- тидні і кожна з них містить одну молекулу ДНК;
синтетичний (S) — відбувається синтез ДНК (реплікація ДНК), унаслідок чого кількість ДНК збільшується вдвічі (2с -> 4с); число хромосом 2п не збільшується, а кожна з них стає подвоєною і відтепер складається з двох ідентичних сестринських хроматид і, відповідно, з двох ідентичних молекул ДНК; кількість гістонових білків, з якими зв’язується синтезована ДНК, теж достовірно подвоюється;
постсинтетичний (G2) — клітина синтезує РНК, білки, у тому числі білки веретена поділу — тубуліни, її об’єм збільшується вдвічі; клітина, готова до поділу, вступає в мітоз; генетична формула клітини — 2п 4с.
Мітоз (каріокінез, непрямий поділ) — основний тип поділу еу- карютичних клітин, у результаті якого з однієї материнської клітини утворюються дві дочірні, генетично ідентичні материнській.
Розрізняють такі фази мітозу (рис. 1.18):
Профаза. Хромосоми спіралізуються (конденсуються) і стають видимими в світловий мікроскоп у формі довгих тонких переплетених ниток (стадія щільного клубка). Поява хромосом — найхарактерніша ознака початку мітозу, звідки він і одержав свою назву (від грец. mitos — нитка). Кожна хромосома має по дві хрома - тиди, які щільно прилягають одна до одної. Унаслідок подальшої спіралізації хромосоми вкорочуються, потовщуються і відокремлюються одна від одної (стадія пухкого клубка). Між хроматидами з’являється щілина.
Друга важлива подія профази — утворення мітотичного апарату, у тварин і нижчих рослин — за участю центріолей, у вищих рослин — без них (центріолі відсутні). Подвійні центріолі розходяться і формують два протилежні полюси клітини. Навколо кожної з них утворюється промениста фігура — центросфера. У міру розходження центріолей між ними виникають нитки веретена поділу. Одні з них (центральні) йдуть від полюса до екватора і далі до протилежного полюса, інші (кінето- хорні) — прикріплюються до кінетохорів центромер. Дві центросфери і веретено між ними утворюють мітотичний апарат, який забезпечує впорядкований рух хромосом в анафазі. Усі його структури побудовані з мікротрубочок, які утворюються шляхом полімеризації тубулінових та інших білків.
У кінці профази зникають ядерця, руйнується ядерна оболонка.
Метафаза починається рухом хромосом до екватора клітини (метакінез). Хромосоми вишиковуються в екваторіальній площині, утворюючи метафазну пластинку, або материнську зірку. При цьому центромери лежать точно в екваторіальній площині перпендикулярно до осі веретена, плечі хромосом обернені до периферії. Конденсація хромосом досягає максимуму. Метафазну пластинку використовують для вивчення каріотипу.

Рис. 1.18. Схема мітозу:
— центріоля;
2 — ядерце; 3 — хромосоми; 4 — рання профаза; 5 — пізня профаза; 6 — метафаза;7 — рання анафаза; 8 — пізня анафаза; 9 — рання телофаза
7 8 9
Анафаза. Центромери одночасно розділяються і хроматиди (тепер їх називають дочірніми хромосомами) синхронно рухаються до протилежних полюсів. Одна хроматида (половинка) від кожної хромосоми йде до одного полюса, інша — до протилежного. Рух хромосом забезпечують мікротрубочки веретена поділу. На полюсах клітини формуються два ідентичні диплоїдні набори хромосом (у людини — по 46). Хромосоми однохроматидні. Генетична формула цієї стадії — 4п4с.Телофаза — заключна фаза мітозу, яка характеризується зворотними процесами порівняно з профазою: хромосоми деспіралізуються, видовжуються і їх вже не можна розрізнити як окремі структури; мікротрубочки веретена деполімеризуються і зникають, навколо хромосом формується ядерна оболонка, з’являються ядерця; завершується поділ материнського ядра і настає поділ цитоплазми (цитокінвз); утворюються дві дочірні клітини, у кожній з яких 2п хромосом і 2с ДНК (генетична формула — 2п2с).
У ссавців мітоз триває 1—1,5 год, G, — 2—5 год, S — 6—10 год, тривалість G, коливається значно більше. У клітинному циклі виділяють ще період G0, коли клітина перестає розмножуватися і переходить цілком до виконання спеціалізованих функцій.
Біологічне значення мітозу:
— забезпечує точний і рівномірний розподіл хромосом між двома дочірніми кптинами, кожна з них одержує стільки хромосом, скільки їх мала материнська;
дочірні клітини генетично ідентичні материнській та ідентичні між собою (у цьому сенсі мітоз є засобом клонування);
підтримує сталість числа хромосом у соматичних клітинах і наступність їх у ряді клітинних поколінь;
збільшує кількість клітин в організмі і забезпечує у такий спосіб ріст і регенерацію, а в багатьох видів приводить до збільшення їхньої чисельності (найпростіші).
Порушення мітозу пов’язані найчастіше з ушкодженнями структури хромосом, мітотичного апарату, порушенням цитокінезу.
Мітотичний апарат особливо чутливий до зовнішніх факторів: радіації, хімічних речовин (у тому числі лікарських препаратів), алкоголю, вірусних інфекцій, високої температури, рослинних отрут (колхіцин). Це призводить до гальмування мітозу, нерозходження хромосом і зміни числа хромосом у каріотипі дочірніх клітин.
У разі порушення цитокінезу утворюються дво- і багатоядерні клітини. Патологічні мітози часто спостерігають при канцерогенезі, променевій хворобі, вірусних інфекціях. З патологією мітозу пов’язане виникнення соматичних мутацій. Якщо мітоз порушується під час утворення гамет, виникають генеративні мутації.
Мейоз і його біологічне значення
Одним з основних положень клітинної теорії є положення про те, що клітини не виникають заново, а утворюються від уже існуючих клітин при розмноженні. Це положення сформулював німецький патолог Р. Вірхов (1858): «Кожна клітина з клітини».
Основні типи поділу еукаріотичних клітин:
мітоз — тип поділу, яким поділяються всі соматичні клітини і у результаті якого з однієї материнської клітини утворюються дві дочірні, генетично ідентичні материнській; оскільки мітотичний механізм забезпечує точний і рівномірний розподіл хромосом між двома дочірніми клітинами, кожна з них одержує стільки хромосом, скільки їх мала материнська;
мейоз — відбувається при гаметогенезі;
амітоз — прямий поділ соматичних клітин, коли ядро ділиться перетяжкою без утворення мітотичного апарату; вважають, що цей тип поділу трапляється в клітинах злоякісних пухлин.
Мейоз (від грец. теіозіз — зменшення) — особливий тип клітинного поділу, який зменшує число хромосом у 2 рази (2п ->■ п), тому цей тип поділу називають редукційним (від лат. гедисіїо — зменшення). У мейозі з однієї диплоїдної клітини утворюються чотири гаплоїдні. Мейоз відбувається при гаметогенезі на стадії дозрівання гамет.
Мейоз складається з двох послідовних поділів:
першого мейотичного, який і є власне редукційним',
другого мейотичного, або екваційного (зрівняльного, практично відбувається як звичайний мітоз).
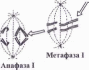
Кожний з цих поділів має чотири фази: профазу, метафазу, анафазу і телофазу (рис. 1.20).
Фази першого мейотичного поділу позначають римською цифрою І, другого — римською цифрою II.
Синтез ДНК відбувається один раз — в інтерфазі І, за якою настає профаза І.
Інтерфаза
Діакінез


у— /Ж
:ж:

Метафаза II

Телофаза II Анафаза II Рис. 1.19. Схема мейозу
Профаза І — найбільш тривала і найбільш складна за процесами, що в ній відбуваються. Від профази мітозу вона відрізняється наявністю кон’югації гомологічних хромосом і кросинговеру між ними. Виділяють п’ять стадій профази І:
Лептонема — стадія тонких ниток. Хромосоми слабо спіралізовані і мають вигляд довгих тонких ниток.
Зигонема — стадія з’єднаних ниток. Гомологічні хромосоми, одна з яких батьківська, а інша — материнська, тісно зближуються по всій довжині і щільно прилягають одна до одної: бік у бік, хромомера до хромомери. Цей процес називають кон’югацією, або синопсисом (контакт), а пару гомологічних хромосом, що кон’югують, — бівалентом.
Пахінема — стадія товстих ниток. Усі хромосоми з’єднані в біваленти, число яких дорівнює гаплоїдному (23). Біваленти вкорочуються і потовщуються за рахунок спіралізації хромосом. Між гомологічними хромосомами відбувається взаємний обмін ділянками — кросинговер.
Диплонема — стадія подвійних ниток. Хромосоми бівалента частково розходяться одна від одної, ніби відштовхуються. Процес відштовхування починається з пентромерних ділянок і поширюється в напрямку кінців. Чітко видно, що кожний
бівалент складається з двох хромосом, а кожна хромосома — з двох хроматид. Біва- лент містить чотири хроматиди і його називають тетрадою.
Діакінез — стадія, протягом якої продовжується процес вкорочення і потовщення хромосом, зникають ядерця, руйнується ядерна оболонка, формується веретено поділу.
Метафаза І. Біваленти (23) вишикуються в зоні екватора, утворюючи мета- фазну пластинку. Центромери хромосом розташовуються на однаковій відстані від екватора — над і під ним. Центромера кожної хромосоми, на відміну від мітозу, з’єднана лише з одним полюсом клітини, але центромери гомологічних хромосом бівалента завжди зв’язані з протилежними полюсами.
Анафаза І. Центромери хромосом не розділяються. До полюсів розходяться не хроматиди від кожної хромосоми, як у мітозі, а цілі гомологічні хромосоми від кожного бівалента. На кожному полюсі формується геплоїдний набір хромосом (23), тобто відбувається редукція числа хромосом. Хромосоми двохроматидні.
Телофаза І. Утворюються дві дочірні клітини. В ядрі кожної з них гаплоїдний набір хромосом (п), але оскільки хромосоми двохроматидні, кількість ДНК відповідає не гаплоїдному, а дипло'ідному набору.
Другий мейотичний поділ настає після короткої інтерфази II, під час якої синтез ДНК не відбувається, і приводить у відповідність число хромосом і кількість ДНК. Він проходить як мітоз. Профаза II дуже коротка. У метафазі II хромосоми (23) вишикуються в екваторіальній площині веретена. В анафазі II центромери розділяються — сестринські хроматиди стають дочірніми хромосомами і розходяться до протилежних полюсів. У телофазі II формуються чотири клітини. Кожна з них містить гаплоїдний набір хромосом (п) і відповідну кількість ДНК.
Особливості обох мейотичних поділів чіткіше проявляються в ході їх порівняльної характеристики (табл. 1.3).
Таблиця 1.3. Порівняльна характеристика мейотичних поділів
Фаза |
Мейоз І |
Мейоз II |
Інтерфаза |
Синтез ДНК відбувається |
Синтез ДНК не відбувається |
Профаза |
Гомологічні хромосоми кон’югують і утворюють пари-біваленти (23). Відбувається кросинговер |
Гомологічні хромосоми не кон’югують і біваленти не утворюються, кросинговер не відбувається |
Метафаза |
На екваторі розміщені біваленти (23). Центромери розташовані над і під екватором. У біваленті 2 хромосоми і 4 хроматиди |
На екваторі розміщені хромосоми (23). Центромери розташовані в екваторіальній площині. У кожній хромосомі — 2 хроматиди |
Анафаза |
Центромери хромосом не розділяються. До протилежних полюсів відходять цілі хромосоми з кожного бівалента. На кожному полюсі формується гаплоїдний набір двохроматидних хромосом |
Центромери хромосом розділяються. До протилежних полюсів відходять хроматиди з кожної хромосоми. На кожному полюсі формується гаплоїдний набір однохроматидних хромосом |
Телофаза |
Утворюються дві дочірні клітини. У кожній клітині гаплоїдний набір хромосом, але кількість ДНК відповідає ще дип- лоїдному набору |
Утворюються чотири клітини. У кожній клітині гаплоїдний набір хромосом і відповідна кількість ДНК |
Біологічне значення мейозу. Гаплоїдність гамет, що утворюються внаслідок мейозу після їх злиття в ході запліднення, дає можливість підтримувати з покоління в покоління постійне чисельне значення хромосом (каріотип) певного біологічного виду. Мейоз забезпечує генетичну неоднорідність гамет унаслідок двох механізмів:
кросинговеру, що зумовлює нові комбінації алелей у хромосомах;
незалежного поєднання внаслідок розходження негомологічних хромосом в анафазі І, що зумовлює різні комбінації батьківських і материнських хромосом у гаметах.
Задача 1. У жінки в процесі овогенезу під час мейозу не відбулося розходження однієї пари гомологічних хромосом. Скільки хромосом буде мати зріла яйцеклітина? Розв’язання.
Правильне розходження хромосом зумовлює появу 23-хромосомних яйцеклітин. Якщо в одній з гомологічних пар розходження не відбулося, це означає, що в одну дочірню клітину надійшло одразу дві гомологічні хромосоми, тобто у неї буде 23 + 1 = 24 хромосоми, а в іншу — не надійшло жодної і її хромосомний комплекс становитиме 23 — 1 = 22 хромосоми.
Відповідь. У жінки можливим є утворення двох типів зрілих яйцеклітин: в одній із них буде 24 хромосоми, а в іншій — 22 хромосоми.
Задача 2. Диплоїдний набір людини становить 46 хромосом. Яка загальна кількість хроматид надходить до кожного полюса: а) в анафазі першого мейотичного поділу; б) в анафазі другого мейотичного поділу?
Розв’язання:
а) в анафазі першого мейотичного поділу, який за своєю суттю є редукційним, до полюсів клітини розходиться по гомологічній хромосомі від кожного бівалента, що зумовлює розходження до кожного полюса загалом 23 двохроматидні хромосоми, або 46 хроматид;
б) у метафазі другого мейотичного поділу метафазна пластинка містить 23 двохроматидні хромосоми, тому в анафазі до кожного полюса надходить по 23 хроматиди.
Відповідь. В анафазі першого мейотичного поділу до кожного полюса надходить по 46 хроматид, а в анафазі другого мейотичного поділу — по 23 хроматиди.
Цікаво знати
Щоб дати уявлення про життєдіяльність клітини, у журналі «Ме\у$\уеек (Ньюсуїк)» наводиться така аналогія: «Кожна із цих ста трильйонів клітин функціонує як оточене стіною місто. Електростанції виробляють для клітини енергію. Фабрики продукують білки — необхідні для хімічного товарообміну продукти. Складні транспортні системи перевозять певні хімікалії всередині клітини від одного місця до іншого, а також за її межі. На прикордонних пунктах стражі перевіряють експорт і імпорт, контролюючи зовнішній світ стосовно ознак небезпеки. Дисципліновані біологічно озброєні сили перебувають напоготові, щоб прийняти необхідні заходи проти загарбників. Центральний генетичний уряд підтримує порядок».
Спостереження за клітинами сполучної тканини, що були вирощені в умовах лабораторії, показали, що здатність кожної клітини цієї лінії до поділу обмежена 50 подвоєннями. Після цього лінія клітин гине. Отже, у клітини «вмонтовано годинник», тобто вони можуть точно відлічувати кількість поділів своєї лінії.
Культивовану лінію клітин можна перетворити на «безсмертну», обробивши вірусом раку. На сьогодні існує близько 600 безсмертних, трансформованих клітинних ліній.
У ході кросинговеру число математичних рекомбінацій між генами людини настільки велике (враховуючи наявність 23 груп зчеплення та можливість обміну між хроматидами у кожному випадку різними ділянками), що вірогідність утворення гамет з одним і тим же геномом мізерно мала. Математичне моделювання процесів кросинговеру в клітинах людини засвідчило, що для реалізації вірогідності народження у однієї і тієї ж шлюбної пари двох дітей з однаковим генотипом необхідне народження мільярдного потомства!
За все життя сім'яники чоловіка здатні виробити 2000 мільярдів сперматозоїдів. В одному його еякуляті наявна кількість сперматозоїдів, здатна запліднити усіх жінок сучасної Європи. Ось який дивовижний запас успішності запліднення і продовження людського життя закладено природою!
Комплекс різних гетерохромосом чоловічої та жіночої статі визначає ознаки статевого диморфізму між чоловіками і жінками, які проявляються не лише в анатомо-мор- фологічних особливостях.
Так, жінки краще чують. їхнє вміння розпізнавати і локалізувати звуки перевищує можливості чоловіків усіх вікових категорій. Чоловіки, своєю чергою, краще бачать. Дівчатка починають говорити раніше, ніж хлопчики, вони мають більший словниковий запас, чіткіше вимовляють звуки, швидше засвоюють орфографію і письмо. Але хлопчики краще орієнтуються у просторі, розташуванні об'єктів за певною схемою. Чоловік фізично сильніший, але менш гнучкий, більш незалежний, агресивніший, здатний ризикувати, честолюбний. Йому притаманний дух змагання більше, ніж жінкам. Він краще володіє просторовими, числовими і механічними здібностями, більше схиляється до того, щоб будувати світ у термінах об'єктів, ідей і теорій.
Жінці із самого початку притаманні властивості, які полегшують міжособистісне спілкування. її фізичний і психологічний розвиток відбувається швидше, вона більш вихована, вишукано і правильно користується мовою, дружелюбніша і послідовніша, ніж чоловіки, більше схиляється будувати світу термінах особистісних, моральних і естетичних цінностей.
Контрольні завдання до розділу 1
Питання для самоконтролю та обговорення
Які найважливіші властивості життя?
Чим відрізняються захисні функції білків і ліпідів?
Яка будова та функції мітохондрій, апарату Гольджі, хлоропластів?
Які відмінності між активним і пасивним транспортом речовин?
Що спільного і чим відрізняються процеси піноцитозу та фагоцитозу?
Який процес більш продуктивний — гліколіз чи дихання? Чому?
Що таке клон?
Які хромосоми є гомологічними?
Які відомі типи симетрії хромосом?
Як ви розумієте відмінності між поняттями «каріотип» і «каріограма»?
Що таке клітинний цикл?
Який із періодів інтерфази найважливіший? Чому?
Що спільного і які відмінності між мітозом, першим та другим мейотичним поділами?
У чому біологічне значення мітозу і мейозу?
Чому в людини жіночу стать називають гомогаметною, а чоловічу — гетерогаметною?
Чим пояснюється статистично однакове співвідношення між потомками чоловічої і жіночої статі ’
Яка особливість статевих хромосом зумовила їх назву?
Контрольно-навчальні завдання
Тести
До органогенних елементів належать:
Бе, Са, СІ, Мё.
Б. N1, гп, Р, І.
О, 14, Н, С.
Г. Б, АІ, Аь, В.
Г. К, N8, Мп, Со.
Синтез АТФ у рослинних клітинах здійснюється:
В ядрі.
Б. На рибосомах.
У мітохондріях.
Г. В апараті Гольджі.
Г. У хлоропластах.
Моносахариди не входять до складу:
Крохмалю.
Б. Глікогену.
Целюлози.
Г. Гемоглобіну.
Ґ. Сахарози.
ДНК - це:
Дисахарид.
Б. Мономер.
Поліпептид.
Г. Полісахарид.
Г. Полінуклеотид.
Власну ДНК мають:
Рибосоми.
Б. Мітохондрії.
Апарат Гольджі.
Г. Лізосоми.
Г. Ендоплазматична сітка.
Функцію внутрішньоклітинного травлення забезпечують:
Хлоропласти.
Б. Рибосоми.
Лізосоми.
Г. Мітохондрії.
Г. Апарат Гольджі.
Перевагою молекули ДНК як носія генетичної інформації над РНК є те, що ДНК здатна до:
Інверсії.
Б. Репарації.
Реплікації.
Г. Транскрипції.
Г. Трансляції.
Хроматин — це комплекс:
А. РНК з білком.
Б. Вуглеводу з білком.
В. ДНК з білком.
Г. Білків і ліпідів.
Ґ. ДНК з РНК.
Кристи — елемент будови:
Хлоропластів.
Б. Рибосом.
Гранулярної ендоплазматичної сітки. Г. Мітохондрій.
Г. Лізосом.
Пасивний транспорт речовин забезпечується:
Простою дифузією.
Б. Піноцитозом.
Фагоцитозом.
Г. Екзоцитозом.
Ґ. Ендоцитозом.
У нормі в каріотипі людини відсутні хромосоми:
Акроцентричні.
Б. Метацентричні.
Субметацентричні.
Г. Супутникові.
Г. Телоцентричні.
В якій фазі мітозу відновлюється ядерна оболонка:
У профазі Б. У метафазі.
В інтерфазі.
Г. В анафазі.
Ґ. У телофазі?
Каріотип людини вивчають, коли клітина знаходиться в стані:
Профази.
Б. Метафази.
Анафази.
Г. Телофази.
Ґ. Інтерфази.
У процесі гліколізу в енергію макроергіч- них звя’зків АТФ переходить:
40 кДж.
Б. 720 кДж.
1520 кДж.
Г. 1440 кДж.
Ґ. 80 кДж.
У синтетичний період інтерфази:
А. Відбувається репарація ДНК.
Б. Відбувається транскрипція.
В. Відбувається реплікація ДНК.
Г. Зменшується число хромосом.
Ґ. Клітина нічого не синтезує.
Якщо хромосоми перебувають у конденсованому стані, то:
Функціонують усі гени.
Б. Кількість хромосом збільшується вдвічі.
Відбувається реплікація ДНК.
Г. Хромосоми набувають однохрома- тидного стану.
Г. Гени не функціонують.
Ділянки хромосом, до яких приєднуються нитки веретена поділу, називають:
Теломерами.
Б. Кінетохорами.
Плечами хромосом.
Г. Супутниками.
Г. Вторинними перетяжками хромосом.
У дочірніх клітинах людини після першого мейотичного поділу (редукційного) міститься така загальна кількість хроматид:
23.
Б. 46.
24.
Г. 92.
Г. 69.
Гаплоїдна кількість двохроматидних хромосом знаходиться в екваторіальній площині клітини в:
Анафазі першого мейотичного поділу.
Б. Метафазі другого мейотичного поділу.
Телофазі першого мейотичного поділу.
Г. Телофазі другого мейотичного поділу.
Ґ. Метафазі першого мейотичного поділу.
Кон’югація хромосом відбувається в:
Профазі другого мейотичного поділу.
Б. Профазі першого мейотичного поділу.
Метафазі другого мейотичного поділу.
Г. Анафазі першого мейотичного поділу.
Г. Телофазі першого мейотичного поділу.
Задачі та вправи
Клітина, що проходила життєвий цикл, під час синтетичного періоду опинилася в несприятливих умовах. Ця клітина мала 20 хромосом, але одна з її молекул ДНК не зазнала реплікації. Незважаючи на це, відбувся мітотичний поділ цієї клітини. Скільки хромосом одержали дочірні клітини?
Клітина, каріотип якої складався з 20 хромосом, нормально пройшла інтерфазу, але в процесі мітотичного поділу одна з ниток мітотичного веретена не утворилася. Скільки хромосом одержали дочірні клітини?
У клітині А в пресинтетичний період виник мутантний ген. У клітині Б мутантний ген виник у постсинтетичний період. В обох клітинах відбувся нормальний мітоз. Яка кількість дочірніх клітин отримала мутантні гени?
Унаслідок порушення овогенезу у жінки утворився овоцит другого порядку з 24 хромосомами. Яка ймовірність того, що яйцеклітина також матиме 24 хромосоми?
Унаслідок порушення мейозу утворився сперматозоон з нормальним набором ауто- сом, але з двома Х-хромосомами. У чому полягало порушення мейозу?
Яка кількість речовини АТФ утвориться при гліколізі 450 г глюкози?
Олімпіада для допитливих
Довести, що виникнення хлоропластів і мітохондрій є ароморфозом в еволюції живої природи.
Чому вважають, що всі особливості і властивості білка визначаються його первинною структурою, а не вторинною чи третинною?
Відомо, що багато тварин (корови, кози, верблюди) не здатні синтезувати незамінні амінокислоти. Живляться вони рослинною їжею, в якій немає незамінних амінокис-
лот. Яким же чином у білку молока, м’язовій тканині з’являються всі 20 амінокислот? Де і як вони утворюються в організмі тварин, якщо їх ні синтезувати, ні одержувати з рослинною їжею тварини не можуть?
Описати фізичні, хімічні та деякі аномальні властивості води, які біологічні об’єкти використовують у своїй життєдіяльності.
Чому у вищих рослин відкладаються про запас переважно вуглеводи, а у тварин — жири?
Багато комах та їх личинки (міль платтяна, лубоїди, пухоїди тощо) живляться виключно сухою їжею і зовсім не споживають води. Звідки таким чином вони беруть воду, необхідну для життєдіяльності?
У каріотипі людини є 46 хромосом, шимпанзе — 48, кролика — 44, річкового рака — 116, мухи-дрозофіли — 8, капусти — 18. Чому ці каріотипні значення виражаються тільки парними числами?
В якій хромосомі людини гени завжди залишаються зчепленими?
Упродовж життя спадкова інформація людини практично не змінюється. Як же зрозуміти, що діти в одній і тій самій сім’ї, тобто народжені одними і тими ж батьками, не є абсолютними генетичними двійниками, а іноді дуже суттєво різняться за низкою ознак?
Відповіді на тести
1В, 2В, ЗГ, 4Ґ, 5Б, 6В, 7В, 8В, 9Г, 10А, 11Ґ, 12Ґ, 13Б, 14Ґ, 15В, 16Ґ, 17Б, 18Б, 19Б,
20Б.
Відповіді на задачі та вправи
20 і 19, бо одна з хромосом залишилася однохроматидною і її одержала лише одна з дочірніх клітин.
19 і 21, бо дві хромосоми не розійшлися і разом досягли одного з полюсів.
Три клітини: дві дочірні від клітини А і одна з дочірніх від клітини Б.
100 %.
Нерозходження хроматид в анафазі другого поділу мейозу.
5 моль.
Відповіді на олімпіадні завдання
Поява хлоропластів зумовила принципово й якісно новий етап живлення й енергетичних перетворень первинних хлорофілоносних структур — фотосинтез із подальшим його вдосконаленням. Завдяки мітохондріям з’явилася можливість здійснювати ефективне вивільнення хімічної енергії з поживних речовин за допомогою біологічного окиснення їх до простих кінцевих продуктів — С02і Н20.
Усі численні білки, що існують у природі, різняться за характером лінійної послідовності сполучення між собою амінокислот. Саме ця чітко визначена послідовність, яка зберігається незмінною в наступних поколіннях і становить суворо індивідуальну первинну структуру білка; звідси і його специфічні індивідуальні властивості.
Незамінні амінокислоти синтезуються мікроорганізмами, які живуть у перших відділах складного шлунка цих тварин. Після того як рослинна маса пережована, вона разом із мікроорганізмами надходить до наступного відділу, де остаточно перетравлюється. Амінокислоти мікробних тіл з іншими поживними речовинами їжі потрапляють в організм тварини і використовуються за призначенням.
У рослин продуктами фотосинтетичної діяльності є розчинні сахариди, які перетворюються на більш хімічно інертні полісахариди. У тварин зручнішим є механізм перетворення надлишку поживних речовин (глікогену печінки) на хімічно інертні жири.
Ці організми використовують так звану метаболічну воду, молекули якої у значній кількості утворюються під час реакцій розкладання полісахаридів, білків і жирів споживаної ними їжі — деревини, шерсті тощо.
Каріотипне значення завжди виражається диплоїдним набором хромосом — 2п. З математичної точки зору при будь-якому п, 2п буде числом парним.
В У-хромосомі, оскільки вона не бере участь у кросинговері (за винятком невеликої ділянки).
Кожна гамета в одного і того ж організму генетично унікальна і неповторна, що забезпечується кросинговером.
РОЗДІЛ 2
МОЛЕКУЛЯРНІ ОСНОВИ СПАДКОВОСТІ. РЕАЛІЗАЦІЯ СПАДКОВОЇ ІНФОРМАЦІЇ
Чому це актуально?
Молекулярні процеси, що відбуваються за участю ДНК та РНК, забезпечують здатність клітин підтримувати високу впорядкованість своєї організації завдяки генетичній інформації, що зберігається, відтворюється, реалізується й удосконалюється.
Незмінність молекулярної структури нуклеїнових кислот є необхідною умовою збереження генетичного здоров'я. Тому знання будови і функцій ДНК та РНК дає змогу втручатися в тонкі механізми спадковості з метою лікування багатьох хвороб людини.

Загальновідомо, що білки клітини визначають її структуру, форму і функції. Проте в природі не доведено жодного випадку, коли б молекула білка синтезувала собі подібну молекулу.
Біологічна інформація зберігається в генах у структурі ДНК у такій формі, що вона може точно копіюватися і передаватися наступним клітинам.
Спадкова інформація ДНК записана в лінійній послідовності нуклеотидів. Передавання {реплікація) інформації розпочинається поділом двох комплементарних ланцюгів, на кожному з яких утворюється нова молекула ДНК. Під час транскрипції певних фрагментів ДНК утворюється мРНК або ІРНК. Молекули РНК одноланцюгові, вони коротші за ДНК.
Транскрипція генетичної інформації з ДНК на РНК і є першим кроком потоку біологічної інформації.
Наступний крок — трансляція мРНК. Молекула мРНК залишає ядро, виходить у цитоплазму і скеровує синтез певного білка на рибосомах. Перенесення Рис. 2.1. Структура ДНК
інформації від мРНК до білка грунтується на принципі комплементарності основ, як і перенесення генетичної інформації від ДНК до РНК.
Отже, генетична інформація записана в лінійній послідовності нуклеотидів ДНК. За участю РНК ця інформація надходить (транслюється) до рибосом з утворенням білкового поліпептиду.
Макромолекула ДНК — це два довгі полімерні ланцюги, що тісно з’єднані між собою (рис. 2.1). Кожен із цих ланцюгів є полінуклеотидом, тобто їх мономерами слугують нуклеотиди, які, своєю чергою, є тримерами, що складаються з азотистої основи, моносахариду — дезоксирибози (у нуклеотидах РНК-рибози) та залишку ор- тофосфатної кислоти. Азотисті основи бувають: пуринові — аденін (А) і гуанін (Г) та піримідинові — тимін (Т) і цитозин (Ц). Загалом чотири різновиди азотистих основ визначають чотири типи нуклеотидів.
Сполучення нуклеотидів у молекулі ДНК відбувається внаслідок взаємодії фосфату одного нуклеотиду з гідроксильною групою дезоксирибози іншого. У результаті утворюється фосфодіефірний зв’язок, що об’єднує нуклеотиди в довгий ланцюжок — полінуклеотид.
Два полінуклеотидні ланцюги ДНК не є ідентичними, але вони комплементарні один одному. Це пов’язано зі строгою відповідністю основ одного ланцюга основам іншого. Відстань між двома ланцюгами ДНК — 2 нм, що дає змогу вмістити лише одну пару А-Т або Г-Ц, які відповідають цим розмірам. Тільки аденін і тимін, а також гуанін і цитозин мають відповідні просторові структури для утворення водневих зв’язків. Отже, аденін в одному ланцюгу має завжди відповідати тиміну в іншому, а гуанін завжди повинен мати навпроти себе цитозин.
Аденін з’єднується з тиміном двома водневими зв’язками, а гуанін з цитозином — трьома водневими зв’язками. Таке сполучення азотистих основ забезпечує міцний зв’язок обох ланцюгів ДНК.
Два полінуклеотидні ланцюги ДНК антипаралельні. Тобто, 5'-кінець одного ланцюга з’єднаний із З'-кінцем іншого, і навпаки. Генетична інформація записана послідовністю нуклеотидів у напрямку від 5'-кінця до З'-кінця. Таку нитку називають «змістовною», саме тут розташовані гени (матричний ланцюг). Другий ланцюг ДНК необхідний як «еталон» збереження генетичної інформації і набуває значення у процесах реплікації та репарації.
Саме таку модель будови молекули ДНК запропонували в 1953 р. Д. Уотсон і Ф. Крік.
Спадкова інформація зберігається в молекулі ДНК, проте ДНК не бере безпосередньої участі в життєдіяльності клітин. Роль посередників у передачі спадкової інформації від ДНК у цитоплазму відіграють РНК. Взаємовідносини ДНК, РНК і білків можна уявити у вигляді схеми: ДНК -> РНК -* білок.
РНК мають вигляд довгих нерозгалужених полімерних молекул, що складаються з одного ланцюга. РНК — полімер рибонуклеотидів, що складаються із залишків ортофосфатної кислоти, рибози й азотистих основ (аденін, гуанін, цитозин, урацил). Усі різновиди РНК синтезуються на молекулах ДНК за участю ферментів РНК- полімераз на основі принципу комплементарності. При цьому в синтезованій молекулі аденін ДНК комплементарний урацилу РНК, а гуанін — цитозину.
Молекули РНК відрізняються за низкою ознак від молекул ДНК:
вуглеводом РНК є рибоза;
РНК не містить тиміну, його місце в молекулі займає урацил;
РНК — одноланцюговий полі- нуклеотид;

Рис. 2.2. Будова молекули тРНК:
1—3 — петлі; 4 — ОНЗ'-кінець; 5—У-кшець; 6 — антикодон; 7 — модифікаційні нуклеотиди
правила Чаргаффа не виконуються.
На основі розміру, структури і функції молекул розрізняють три типи РНК.
Інформаційна РНК (іРНК). її молекули утворюються на певних ділянках ДНК, що мають назву структурних генів, у вигляді комплементарної копії ділянки одного з її ланцюгів. іРНК є матрицею для синтезу поліпептидів (білків), тому її також називають матричною. Матрична РНК є шаблоном, на якому будуються поліпептиди відповідно до закладеної генетичної інформації.
Транспортна РНК (тРНК). Молекули тРНК утворюються на спеціальних генах, вони короткі, мають трійчасту форму, що нагадує листок конюшини (рис. 2.2). Молекули тРНК переносять до місць синтезу білків тільки відповідні їм амінокислоти з цитоплазми. Кожній амінокислоті відповідає своя тРНК. Молекули тРНК мають чотири важливі ділянки:
транспортну;
антикодон;
ділянку приєднання ферменту;
ділянку зв’язування з рибосомою.
До транспортної ділянки приєднується специфічна амінокислота. Вона утворена двома комплементарними кінцевими ділянками РНК, З'-кінець якої закінчується послідовністю ЦЦА з вільною ОН-групою. До цієї групи приєднується амінокислота, що транспортується.
У центрі антикодону наявний триплет, комплементарний до триплету на іРНК, який називають кодоном. У період синтезу білка антикодон знаходить відповідний йому кодон на іРНК і тимчасово приєднується до нього водневими зв’язками.
Ділянка приєднання ферменту — це спеціальна частина молекули тРНК для специфічного зв’язування з ферментом аміноацил-тРНК-синтетазою, що каталізує приєднання до молекули тРНК амінокислоти.
Ділянка зв’язування з рибосомою — особлива частина молекули (певна послідовність нуклеотидів) тРНК, що потрібна для прикріплення до рибосоми.
Рибосомна РНК (рРНК) утворюється на спеціальних генах ДНК в ядерці. У каріоплазмі рРНК різні білки об’єднуються для утворення малих і великих субоди- ниць рибосом. Рибосомна РНК утворює структурний каркас рибосоми, забезпечує зв’язування іРНК з рибосомами за допомогою певних послідовностей нуклеотидів.
Універсальність та індивідуальна специфічність ДНК. Правила Чаргаффа
Молекули ДНК у різних рослин і тварин побудовані з одних і тих самих нук- леотидів чотирьох типів: аденілового, гуанілового, цитидилового і тимідилового (А, Г, Ц, Т), але різняться між собою за кількістю і порядком чергування нуклеотидів. Ця обставина визначає універсальність й одночасно унікальність ДНК. ДНК кожного виду характеризується відносною постійністю і видовою специфічністю. Але будь- яка ДНК характеризується певними закономірностями, встановленими в 1950 р. американським біохіміком Е. Чаргаффом.
Правила Чаргаффа:
Сума пуринових основ дорівнює сумі піримідинових основ: А + Г = Т + Ц.
Кількість основ з кетогрупою в 6-му положенні дорівнює кількості основ з аміногрупою в 6-му положенні: Г + Т = А + Ц, або (Г + Т) / (А + Ц) = 1.
Вміст аденіну дорівнює вмісту тиміну, а вміст гуаніну дорівнює вмісту цитозину (правило еквівалентності): А = Т; Г = Ц.
Відповідно до цих правил склад ДНК різних організмів може варіювати лише за співвідношенням (А + Т) / (Г + Ц), але для організмів цього виду таке співвідношення є постійним і називається коефіцієнтом специфічності ДНК (Ксп ДНК). КСП ДНК людини дорівнює 1,54. Ксп ДНК бактерії кишкової палички дорівнює 1,0.
Задача. Знайти кількісні співвідношення нуклеотидів у ділянці молекули ДНК, якщо відомо, що вона містить 240 тимідилових нуклеотидів, що становить 20 % загальної кількості нуклеотидів.
Розв’язання.
Якщо 20 % — 240 нуклеотидів,
то 100 % — х нуклеотидів.
„ 240x100% ,ААА ,
Звідси х = = 1200 (нуклеотидів)
20%
За правилом Чаргаффа, Т = А і, відповідно до умови, дорівнює 240.
Отже, А + Т = 240 + 240 = 480. Тоді Г + Ц = 1200 - 480 = 720. Оскільки Г = Ц, то Г = Ц = 720/2 = 360.
Відповідь. Ділянка ДНК містить: 240 тимідилових, 240 аденілових, 360 гуанілових і 360 цитозилових нуклеотидів.
Генетична роль нуклеїнових кислот
Оскільки відомо, шо з ДНК побудовані гени, які контролюють усі процеси в клітині шляхом синтезу ферментів та інших білків, що, своєю чергою, визначають синтез інших речовин клітини, то, по суті, ДНК є носієм генетичної (спадкової) інформації. Таким чином, генетична інформація — це інформація про білки. У зв’язку з цим розрізняють такі функції ДНК як спадкового матеріалу:
1. Збереження снадкової інформації, на основі якої формується фенотип, що стає можливим унаслідок існування генетичного коду. Генетичний код — система запису спадкової інформації, за якою послідовність нуклеотидів у молекулах нуклеїнових кислот визначає послідовність амінокислот у молекулах білків. Оскільки в процесі реалізації генетична інформація переписується з ДНК на іРНК, генетичний код читається за і PH К і записується за допомогою чотирьох азотистих основ PH К (А, У, Г, Ц).
Генетичний код (табл. 2.1) характеризується такими властивостями:
триплетність — кожна амінокислота кодується трьома сусідніми нуклеотидами (триплетом);
універсальність — код єдиний для всіх організмів, одні й ті самі триплети кодують одні й ті самі амінокислоти у різних біологічних видів. Завдяки універсальності генетичного коду гени скеровують синтез відповідних білків навіть у чужих клітинах, якщо ці клітини включили їх у свої ДНК;
виродженість, або множинність, — одна амінокислота може кодуватися більш ніж одним триплетом. Оскільки існує чотири типи нуклеотидів, генетичний код складається з 64 кодонів (43 = 64), з них 61 кодон кодує 20 амінокислот. Три колони (УАГ, УАА, УГА) — нонсенс-кодони, не кодують жодної амінокислоти і для них не існує тРНК; вони виконують роль сигналів завершення трансляції (стоп-кодо- ни, кодони-термінатори). Одним триплетом кодуються метіонін і триптофан, інші 18 з 20 амінокислот — кількома (2, 3, 4 і 6).
Виродженість захищає генетичний код від мутаційних замін. Якщо в кодоні АГЦ, який кодує аргінін, станеться заміна цитозину на аденін, то зміст спадкової інформації від цього не порушиться і в білковій молекулі не відбудеться жодних змін, оскільки кодон АГА також кодує аргінін;
специфічність — кожний окремий триплет кодує лише одну певну амінокислоту. Специфічність не суперечить множинності. Наприклад, триплет УУА кодує лише лейцин, тобто для цієї амінокислоти він є специфічним. Однак із цього не випливає, що лейцин не може кодуватися іншими триплетами, але це вже відображається такою властивістю, як множинність;
Таблиця 2.1. Генетичний код
Перша основа |
Друга основа |
Третя основа |
||||
У |
П |
А |
г |
|||
|
ФЕН |
СЕР |
ТИР |
цис |
У |
|
|
ФЕН |
СЕР |
ТИР |
цис |
Ц |
|
У |
ЛЕИ |
СЕР |
Ї^ОИ 1 |
ІЧОИ 2 |
А |
|
|
лей |
СЕР |
N014 3 |
ТРИ |
Г |
|
|
ЛЕЙ |
ПРО |
Г1С |
АРГ |
У |
|
ц |
ЛЕИ |
ПРО |
ГІС |
АРГ |
ц |
|
ЛЕИ |
ПРО |
глн |
АРГ |
А |
||
|
ЛЕИ |
ПРО |
глн |
АРГ |
Г |
|
|
ІЛЕ |
ТРЕ |
АСН |
СЕР |
У |
|
|
ІЛЕ |
ТРЕ |
АСН |
СЕР |
Ц |
|
А |
ІЛЕ |
ТРЕ |
ЛІЗ |
АРГ |
А |
|
|
МЕТ |
ТРЕ |
ЛІЗ |
АРГ |
Г |
|
|
ВАЛ |
АЛА |
АСИ |
ГЛІ |
У |
|
|
ВАЛ |
АЛА |
АСИ |
ГЛІ |
Ц |
|
1 |
ВАЛ |
АЛА |
ГЛУ |
ГЛІ |
А |
|
|
ВАЛ |
АЛА |
ТЛУ |
ГЛІ |
Г |
|
неперекривність — конкретний нуклеотид входить до складу лише одного триплету, тобто якщо нуклеотид увійшов до складу попереднього триплету, то до складу наступного він уже входити не може;
колінеарність — послідовність триплетів у ДНК визначає порядок розташування амінокислот у молекулі білка. Це означає, що положення кожної амінокислоти в білковому поліпептиді залежить від положення триплету у молекулі ДНК. По суті, ген і поліпептид, який він кодує, є колінеарними;
переважання двох перших нуклеотидів, тобто із трьох нуклеотид і в кодону третій може варіювати.
Генетичний код має дивовижну компактність. Так, запис інформації, яка міститься в хромосомі тваринної клітини у вигляді літер, що символізують триплети нуклеотидів, становитиме книгу обсягом понад 500 тис. сторінок.
Відтворення спадкової інформації та її передавання наступним поколінням клітин. Відтворення відбувається у 8-період інтерфази і забезпечується реплікацією молекули ДНК, передавання здійснюється в ході мітотичного механізму.
Реплікація (самоподвоєння, самовідтворення) — синтез ДНК на матриці ДНК, який забезпечує точне копіювання спадкової інформації. Здатність до самоподвоєння — унікальна властивість ДНК, притаманна лише їй і можлива завдяки ком- плементарності двох полінуклеотидів її молекули.
Етапами реплікації є:
розкручування біоспіралі ДНК;
локальне розведення її ланцюгів;
стабілізація однониткових полінуклеотидів ДНК;
Рис.
2.3.
Напівконсервативний механізм реплікації
ДНК: 1
— материнська молекула ДНК; 2
— нова дочірня молекула ДНК; 3
— материнський ланцюг; 4
— дочірній ланцюг
— поява з двох розведених ланцюгів У-подібної структури (реплікативної вилки).
У такий спосіб утворюються дві дочірні молекули, ідентичні материнській. Кожна дочірня молекула складається з однієї старої (материнської) нитки й однієї нової нитки. Реплікація ДНК моделює процес самовідтворення на молекулярному рівні організації.
Оскільки тільки один материнський полінуклеотид збережений у кожній дочірній молекулі, такий тип реплікації має назву напівконсервативного (рис. 2.3).
Схема подвоєння ланцюгів ДНК за Р. Окадзакі наведена нижче. Один ланцюг (передовий) синтезується безперервно в напрямку руху реплікативної вилки, а другий (неметкий) — переривчасто з утворенням коротких фрагментів, так званих фрагментів Окадзакі (японський учений, який уперше їх відкрив), у напрямку, протилежному руху реплікативної вилки:
З' Молекула «материнської»
ДНК

Забезпечення специфічних процесів у ході реалізації спадкової інформації, тобто під час біосинтезу білка.
Функції РНК полягають у тому, що всі види РНК забезпечують процеси, які супроводжують етапи синтезу білка: транскрипцію, трансляцію, реакції матричного синтезу.
Центральна догма молекулярної біології
Положення про те, що носієм генетичної інформації є ДНК і що ця інформація реалізується в процесі біосинтезу білка, відоме як центральна догма (основний постулат) молекулярної біології. Її суть виражається схемою, запропонованою у 1958 р. Ф. Кріком.
днк
( > дНК Транскрипція _ рнк
Трансляція
>
У 1960 р. український генетик С.М. Гершензон на основі своїх досліджень одного з вірусних захворювань шовкопряда вперше висловив припущення про існування РНК-залежного синтезу ДНК. Американські генетики Г. Темін, Д. Балтимор (1970) виявили в РНК-вмісних онкогенних вірусів фермент, який каталізує синтез ДНК на матриці іРНК (зворотна транскрипція).
У зв’язку з відкриттям зворотної транскрипції схема центральної догми молекулярної біології набула такого вигляду:
ДНК ^ ДНК <=>РНК Білок.
Насправді розвиток будь-яких ознак є результатом складних взаємодій між генами на рівні генетичного матеріалу, у процесі біосинтезу білка, між білками-фер- ментами. Ознака формується під впливом не одного, а багатьох ферментів за певних умов середовища.
Ген. Сучасний стан теорії гена
Одним з основних положень генетики є положення про те, що всі ознаки і властивості організмів визначаються одиницями спадковості — генами (від грец. — рід, походження).
Ген — елементарний, дискретний, матеріальний спадковий фактор — функціональна одиниця генетичної інформації; ділянка молекули ДНК (у деяких вірусів РНК), яка містить інформацію про послідовність амінокислот у молекулі поліпептиду (білка) або послідовність нуклеотидів у молекулах РНК.
Ген характеризується такими ознаками:
Кожний ген складається приблизно з 1000 пар нуклеотидів. Набір нуклеотидів та їх послідовність у кожному гені унікальні, що зумовлює специфічність гена, тобто здатність контролювати лише певну ознаку.
Більшість генів в еукаріотів є унікальними і представлені в геномі лише однією копією. Поряд з цим існують гени (гени рРНК, тРНК, гістонові гени), які представлені блоками, що складаються з тандемно повторюваних (один за одним) ідентичних генів або груп генів.
Найменшу функціональну одиницю генетичного матеріалу, яка кодує синтез певного поліпептиду, називають цистроном. Один цистрон кодує один поліпептид. Терміни «цистрон» і «ген» вживають практично як синоніми.
Ген має складну будову і здатний до мутацій та рекомбінацій. Найменшу ділянку цистрона, зміна якої може спричинити мутацію, називають мутоном (одиниця мутації), а найменшу ділянку, яка змінюється шляхом рекомбінації, — реконом (одиниця рекомбінації). Довжину мутонів і реконів найчастіше визначає одна (рідше дві) пара нуклеотидів.
Гени розташовані у хромосомах у певній послідовності в лінійному порядку (один за одним). Кожний ген займає певне місце (локус).
Існує кілька класифікацій генів. За функцією гени поділяють на:
структурні — кодують синтез білків (по суті, це гени ІРНК);
гени тРНК — кодують синтез тРНК;
гени рРНК — кодують синтез рРНК;
регуляторні — контролюють активність структурних генів.
Структурні гени еукаріотів мають мозаїчну будову. Вони містять екзони (інформативні ділянки кодують амінокислоти) та інтрони (неінформативні ділянки не кодують амінокислот).
Ген не бере участі в біосинтезі білка, він є матрицею для утворення посередників — різних молекул РНК, які безпосередньо забезпечують синтез.
Активність гена знаходиться під генним контролем та контролем факторів зовнішнього середовища.
Молекулярна організація геному еукаріот характеризується надлишковою кількістю ДНК. Так, розмір геному людини достатній для утворення близько 1,5 млн генів, але фактично в ньому міститься лише 35 тис. генів і більша частина ядерної ДНК не транслюється в амінокислотну послідовність.
Молекули ДНК здатні до репарації, тому не всі ушкодження гена ведуть до мутації.
Гени мають фіксовану локалізацію на хромосомі, тобто займають певний локус. Відомі також мобільні (міграційні) генетичні елементи (МГЕ), або мобільні гени. Це послідовності нуклеотидів ДНК, здатні до переміщення (транспозиції) по геному клітини. З ними пов’язують виникнення мутацій і варіацій.
Гени, за хромосомною теорією спадковості, розташовані в хромосомах лінійно в певній послідовності і не перекриваються. (Перекривні гени — винятки, виявлені в деяких вірусах.)
В еукаріотів відкриті псевдогени — послідовності нуклеотидів, які схожі на структурні гени, але не транслюються у функціонально активні поліпептиди.
Задача. Полінуклеотид молекули ДНК із 1444 нуклеотидів, що кодує білок, містить п’ять інтронних ділянок — завдовжки 100, 120, 135 та дві по 150 нуклеотидів. Скільки амінокислот у закодованому білку?
Розв’язання.
Оскільки амінокислоти кодуються екзонними ділянками ДНК, то із загальної кількості нуклеотидів слід вилучити нуклеотиди інтронних ділянок:
1444 — (100 + 120 + 135 + 150 х 2) = 879. Ураховуючи, що одна амінокислота кодується трьома нуклеотидами (триплетом), знаходимо кількість амінокислот: па.т= 879 : 3 = 293.
Відповідь. У закодованому білку 293 амінокислоти.
Мутації і рекомбінації генів. Репарація ДНК
Генетична інформація у вигляді певного генотипу успадковується разом із хромосомами батьківських форм і в ідеалі вона не повинна змінюватися впродовж життя. Однак за певних умов, під дією різних за природою факторів (мутагенів), у генах можуть виникати зміни на молекулярному рівні ДНК (мутації чи рекомбінації).
Оскільки молекула ДНК представлена подвійною спіраллю, виникнення мутації відбувається в два етапи. На першому етапі зміна стосується лише одного ланцюга молекули ДНК. Цей стан називають молекулярною гетерозиготністю, або потенційною мутацією. Якщо зміна поширюється на гомологічний локус комплементарного ланцюга, то такий стан називають молекулярною гомозиготністю, або істинною мутацією. Істинна мутація успадковується.
За якісною характеристикою мутації і рекомбінації генів класифікують так:
Трансверсія — переміщення азотистих основ з одного полінуклеотиду в інший:
-А-Г-Ц- -А-Ц-Ц-
-т-ц-г- -т-г-г-
Трансверсія — це рекомбінація (співвідношення азотистих основ до і після не змінюється), тому змінена ділянка гена — рекон, його довжина у наведеній схемі становить одну комплементарну пару.
Транзиція — заміна однієї комплементарної пари на іншу: -А-Г-Ц- -А-Т-Ц-
-Т-Ц-Г- -Т-А-Г-
Транзиція — це мутація (у початковій ланці ДНК співвідношення азотистих основ було 1А:1Т:2Г:2Ц, а після стало 2А:2Т:1Ц:1Г), довжина відповідного мутона — одна комплементарна пара.
Інверсія — повернення ділянки гена на 180":
-А-Г-Ц- -А-Ц-Г-
-т-ц-г- -т-г-ц-
Довжина рекона дорівнює двом комплементарним парам.
Делеція — випадання ділянки цистрона:
-А-Г-Ц- -А-Ц-
-т-ц-г- -т-г-
Дуплікація — подвоєння ділянки цистрона, рідше збільшення його довжини у кратному відношенні:
-А-Г-Ц- - А —Г —Г—Ц-
-т-ц-г- - т-ц-ц-г-
Як правило, мутації і рекомбінації генів перетворюють природний «дикий» домінантний алель на мутантний алель рецесивного характеру, тобто переважна більшість генів, що зазнали змін у власній структурі, є рецесивними, а отже, фенотипово вони здатні проявитися лише у гомозиготному рецесивному генотипі, вірогідність якого мала, так як для його виникнення необхідне одночасне успадкування мутантного гена як від батька, так і від матері. Очевидно, переважна рецесивність мутантних генів є одним із механізмів захисту живими організмами власної спадкової інформації.
У деяких випадках мутантний алель здатний повертатися внаслідок зворотної мутації (реверсії) у початковий стан.
Для забезпечення основних характеристик клітин і організмів цієї популяції необхідне точне зберігання структури і стабільності генетичного матеріалу впродовж тисяч і мільйонів років, незважаючи на дію різних мутагенних чинників. Для підтримання стабільної функції ДНК існує кілька механізмів.
По-перше, це висока хімічна стабільність самої молекули ДНК.
По-друге, — наявність спеціальних механізмів самокорекції і репарації (від лат. герагагіо — відновлення) змін, які виникли. Процес репарації ДНК полягає в тому, що генетична інформація подана ДНК двома копіями — по одній у кожному з двох ланцюгів подвійної спіралі ДНК. Завдяки цьому випадкове ушкодження в одному з ланцюгів може бути видалене реплікаційним ферментом, а ушкоджена ділянка ланцюга — ресинтезована у своєму нормальному вигляді за рахунок інформації, що міститься в неушкодженому ланцюгу.
За часом здійснення у клітинному циклі розрізняють такі види репарації:
дореплікативну — процес відновлення ушкодженої ДНК до її подвоєння, що здійснюється ферментом лігазою або повною ферментативною системою репарації;
реплікативну — сукупність процесів відновлення ДНК у ході реплікації, суть яких полягає у видаленні ушкодженого ланцюга в зоні його росту; при цьому висока точність реплікації забезпечується ДНК-полімеразою;
постреплікативну — процес вирізання ушкодженої ділянки і зшивання кінців, механізм якого ще до кінця не вивчений.
Біосинтез білка. Транскрипція, трансляція
Реалізація генетичної інформації відбувається шляхом матричного біосинтезу білків і здійснюється в два етапи.
Транскрипція — перший етап (від лат. іїатсгірНо — переписування) — синтез РНК на ДНК-матриці. Полягає у переписуванні генетичної інформації з послідовності нуклеотидів ДНК на комплементарну послідовність нуклеотидів іРНК.
Особливості транскрипції:
як матриця використовується лише один ланцюг ДНК;
транскрибується не вся молекула ДНК, а лише певна ділянка (ген чи група генів);
у молекулі ДНК є сигнальні послідовності нуклеотидів, які вказують на початок і кінець транскрипції;
транскрипція каталізується ферментами — ДНК-залежними РНК-полімераза- ми трьох типів: перший — каталізує синтез рРНК, другий — синтез ІРНК, третій — синтез тРНК; у мітохондріях існує своя власна РНК-полімераза.
У свою чергу розрізняють три етапи транскрипції:
Ініціація (початок синтезу ІРНК) — це приєднання першого нуклеотиду. Перед цим РНК-полімераза розпізнає промотор (ділянка ДНК) і зв’язується з ним. Це зумовлює локальне розходження ланцюгів подвійної спіралі ДНК.
Елонгація (нарощування ланцюга іРНК). Фермент послідовно приєднує нук- леотиди до вільної гідроксогрупи рибози, ланцюг РНК синтезується в напрямку 5-3'.
Термінація (закінчення синтезу іРНК). При досягненні ферментом терміна- торів (стоп-кодонів) транскрипція припиняється.
Послідовність нуклеотидів іРНК комплементарна такій ДНК (А-У, Г-Ц, Т-А, Ц-Г). Після завершення транскрипції і звільнення іРНК двоспіральна структура ДНК відновлюється. У ході термінації відбуваються специфічні процеси процесин- гу і сплайсингу.
Процесинг — молекулярні механізми, пов’язані з «дозріванням» різних типів РНК, яке здійснюється в ядрі перед виходом РНК із ядра в цитоплазму.
Сплайсинг — явище вирізання спеціальними ферментами інтронів і зшивання активних ділянок іРНК — екзонів. Відбувається в процесі «дозрівання» іРНК.
Трансляція — другий етап (від лат. ШтІаНо — переклад) — синтез поліпеп- тидних ланцюгів білків на матриці іРНК; зчитування генетичної інформації і переведення її з послідовності кодонів іРНК у послідовність амінокислот поліпептиду. Роль проміжних молекул (адаптерів) між іРНК-матрицею і поліпептидом у процесі декодування генетичної інформації виконують тРНК.
У процесі трансляції фермент аміно- ацил-тРНК-синтетаза за участю АТФ активує амінокислоту і приєднує її до акцепторного кінця тРНК. Комплекс тРНК з активованою амінокислотою має назву аміноацил-тРНК (аа-тРНК). Кожна із 20 амінокислот має власну аміноацил-тРНК-синтетазу, яка здатна розпізнавати (рекогніція) певну амінокислоту і приєднувати її до відповідної тРНК.
Рис.
2.4.
Механізм елонгації поліпептидно- го
ланцюга (пояснення див. у тексті)


А-центр (аміноацильний) приєднує аміноацил-тРНК;
П-центр (пептидильний) зв’язаний із тРНК попередньої амінокислоти поліпептиду, що тут нарощується.
Процес трансляції перебігає у кілька етапів:
Ініціація трансляції — початок синтезу. «Шапочка» (метилгуанозин) іРНК зв’язується з малою субодиницею рибосоми. Навпроти П-центру рибосоми розміщується ініціювальний кодон іРНК — АУГ, що відповідає першій ініціювальній амінокислоті — метіоніну. Отже, синтез будь-якого поліпептиду починається з метіоніну. В А-центр рибосоми прибуває друга тРНК з активованою амінокис-
лотою, яка відповідає першому значущому кодону іРНК. Між двома амінокислотами утворюється пептидний зв’язок.
Елонгація — продовження поліпептидного ланцюга. Друга, навантажена, наприклад проліном, тРНК з’єднується з рибосомою на ділянці А (рис 2.4, а, б). Її антикодон зв’язується з комплементарним кодоном ланцюга іРНК. На ділянці П метіонін звільняється від своєї тРНК і з’єднується пептидним зв’язком з проліном (рис. 2.4, в). Процес каталізує фермент пептидилтрансфераза. У цьому процесі зв’язок між першою амінокислотою та її тРНК розривається і — СООН-група першої амінокислоти утворює пептидний зв’язок з вільною — МН2-групою другої амінокислоти. Таким чином, друга тРНК уже несе дипептид. Перша тРНК, тепер вільна, відокремлюється від П-ділянки рибосоми і повертається у загальний фонд тРНК у цитоплазмі (див. рис. 2.4, в). Тут вона знову може зв’язуватися зі своєю амінокислотою.
тРНК-дипептидний комплекс разом з ІРНК переміщується в напрямку П-ді- лянки рибосоми (рис. 2.4, г). Цей процес називають транслокацією (від лат. Штіосаіїо — переміщення).
Третя молекула тРНК зі специфічною їй амінокислотою, наприклад аргініном, надходить до А-ділянки рибосоми і приєднується своїм антикодоном до комплементарного кодону іРНК (див. рис. 2.4, г). Дипептид метіонін-пролін знову приєднує амінокислоту аргінін. У такий спосіб дипептид збільшується до трипептиду. Друга тРНК відокремлюється, залишає ланцюг ІРНК, звільняючи П-ділянку. тРНК- трипептидний комплекс переноситься з А-ділянки на П-ділянку.
Увесь процес, що включає надходження тРНК-амінокислотного комплексу, утворення пептидного зв’язку і транслокацію, багаторазово повторюється. У міру просування іРНК щодо рибосоми всі її кодони переміщуються по А-ділянці один за одним, і пептидний ланцюг зростає. У процесі елонгації беруть участь спеціальні білкові фактори, що регулюють ці процеси.
Термінація трансляції — закінчення синтезу. Завершення трансляції відбувається, як тільки рибосома досягне одного з кодонів-термінаторів іРНК (УАА, УАГ або УГУ), для яких не існує тРНК. На цій стадії трансляційний комплекс розпадається: поліпептид відділяється від П-центру, тРНК переходить у вільну форму, рибосома дисоціює на дві субодиниці, здатні надалі до нового самоскладання. Послідовність амінокислот у синтезованому поліпептиді відповідає послідовності колонів іРНК. Після того як поліпептидний ланцюг відділився, він зазнає посттран- сляційної модифікації й утворення властивої йому унікальної просторової конформації (вторинна, третинна і четвертинна структури) білкової молекули. Надалі білки, залежно від функції (ферментативна, структурна, регуляторна, захисна та ін.), здатні забезпечувати морфофізіологічні особливості клітини.
Усі розглянуті вище синтетичні реакції каталізують спеціальні ферменти. Ці реакції потребують енергетичних витрат, які компенсуються реакціями розщеплення АТФ, що є універсальною енергетичною речовиною. АТФ під дією ферментів легко втрачає один або два залишки ортофосфатної кислоти; при цьому утворюється відповідно АДФ або АМФ і вивільняється енергія з розрахунку 40 кДж на один макроергічний зв’язок.
Експресія генів. Концепція оперону
Генна експресія — це молекулярний механізм реалізації спадкової інформації, завдяки якому ген виявляє свій потенціал конкретною фенотиповою ознакою організму.
Процес експресії гена складається з кількох етапів:
Код гена ДНК перетворюється на код про-іРНК. Перший етап експресії, таким чином, це — транскрипція.
Складна молекула про-іРНК зазнає процесингу, внаслідок чого значно зменшується за розмірами. Утворюється зріла іРНК, зчитування інформації з якої спрощується, у чому і полягає біологічний зміст процесингу.
іРНК за участю тРНК вибирає необхідні амінокислоти і зв’язує їх на рибосомі відповідно до послідовності амінокислот у молекулі білка. Цей процес є трансляцією.
Синтезований поліпептид зазнає модифікації і впливає на морфологічну або функціональну ознаку (фенотип) клітини або організму. Цей процес і є безпосередньо експресією.
У 1961 р. французькі біологи Ф. Жакоб і Ж. Моно запропонували механізм регуляції генів, який було названо гіпотезою оперону. Оперон — це послідовність спеціальних функціональних сегментів ДНК та структурних генів, які кодують синтез певної групи білків одного метаболічного ланцюга, наприклад ферментів гліколізу. До складу регульованої одиниці транскрипції входять такі структурні частини:
ген-регулятор, який контролює утворення білка-репресора, що, своєю чергою, вмикає або вимикає оператор — ділянку молекули ДНК, яка контролює функцію цистронів (структурних генів);
промотор — ділянка ДНК, до якої приєднується РНК-полімераза і з якої розпочинається транскрипція;
оператор — ділянка промотора, яка може зв’язувати репресор;
структурні гени — ділянки ДНК, які кодують ІРНК конкретних білків;
термінаторна ділянка ДНК, яка несе сигнал про припинення транскрипції.
Розглянемо типові задачі. Для кращого усвідомлення кінцевих результатів реалізації спадкової інформації змоделюємо процеси транскрипції і трансляції на основі невеликої інформаційної ділянки гена. Конкретні амінокислоти, що входять до складу білкового поліпептиду, який утворюється внаслідок трансляції, слід визначати за кодоном іРНК (див. табл. 2.1):
он о |
Н О |
н о |
II 1 II |
1 II |
1 II |
с-и -с- с |
—14 —с —с - |
-N-0 -С -1 |
/\ |
/\ |
/\ |
Н Я| |
Н ц2 |
н |
-Т-Ц-А 1 1 1 |
- г-г-т І 1 1 |
- Ц-А-А- і і і |
1 1 1 -А-Г-Т |
1 1 1 - Ц-Ц-А |
1 1 1 - г- т-т- |
-А-Г-У |
- Ц-Ц-А |
- Г-У-У- |
У-Ц-А, |
Г-Г-У, |
Ц-А-А |
-СЕР |
-про |
-ВАЛ“ |
■м-
Код ДНК:
Кодон ІРНК Антикодон тРНК Амінокислоти
Для розв’язування задач, що моделюють молекулярні основи спадковості, слід мати на увазі таке:
середня молярна маса одного нуклеотиду М = 345 г/моль;
довжина одного нуклеотиду — 0,34 нм;
середня молярна маса однієї амінокислоти М = 100 г/моль;
для визначення довжини гена враховують кількість нуклеотидів, які містяться в одному ланцюзі ДНК;
для визначення молекулярної маси гена враховують кількість нуклеотидів, що містяться в двох ланцюгах ДНК;
молекулярну масу низькомолекулярних білків можна визначити, знаючи масові частки компонентів, які входять до складу білка, їх атомну чи молекулярну масу. Для цього слід скористатися формулою:
М = а/Ь х 100 %,
де М — мінімальна молекулярна маса білка; а — відносна атомна або молекулярна маса компонентів; Ь — масова частка компонентів білка.
Задача 1. Ділянка молекули ДНК має таку послідовність нуклеотидів: АЦЦ-АТА- ГЦТ-ЦАА-ГГА. Визначити послідовність амінокислот у відповідному поліпептидному ланцюзі.
Розв’язання.
Спочатку запишемо послідовність нуклеотидів ДНК, потім за принципом компле- ментарності визначимо послідовність нуклеотидів іРНК і, користуючись таблицею генетичного коду, — послідовність амінокислот у поліпептиді (білку).
Код ДНК: |
АЦЦ - |
АТА |
- ГЦТ - |
ЦАА |
- ГГА |
Кодон іРНК: |
УГГ - |
УАУ |
- ЦГА - |
ГУУ |
- ННУ |
Амінокислоти: |
ТРИ - |
ТИР |
- АРГ - |
ВАЛ |
- ПРО |
Відповідь. Триптофан — тирозин — аргінін — валін — пролін.
Задача 2. На одному з ланцюгів ДНК синтезована іРНК, в якій А — 14 %. Г — 20 %, У — 40 %, Ц — 26 %. Визначити вміст (%) нуклеотидів у молекулі ДНК. Розв'язання.
За умовою склад іРНК: А (14 %) Г (20 %) У (40 %) Ц (26 %)
Матричний полінуклеотид Т (14 %) Ц (20 %) А (40 %) Г (26 %)
ДНК матиме:
Склад комплементарного А (14 %) Г (20 %) Т (40 %) Ц (26 %)
полінуклеотиду ДНК:
Отже, %А = %Т = (40 + 14) : 2 = 27 %. %Г = %Ц = (26 + 20) : 2 = 23 %.
Відповідь. А - 27 %, Т - 27 %, Г - 23 %, Ц - 23 %.
Задача 3. Білковий поліпептид містить 248 амінокислот. Що важче: білок чи ген, який його кодує?
Розв'язання.
Визначимо молярну масу білка. Оскільки молярна маса однієї амінокислоти М і( = 100 г/моль, то МЛІДШ = Мд пі х па т= 100 г/моль х 248 = 24 800 г/моль.
Визначимо кількість нуклеотидів у ділянці ДНК, що відповідає цьому гену: 248 х 3 х 2 =1488.
Оскільки молярна маса одного нуклеотиду Mhvkji = 345 г/моль, то молярна маса гена становитиме: М = М х 1488 = 345 г/моль х 1488 = 5 133 360 г/моль.
ієна нукл ' '
Таким чином, ген важчий від битка, який ним визначається, у 20,7 разу (5 133 360 : 24 800 = 20,7).
Відповідь. Маса гена більша від маси білка у 20,7 разу.
Задача 4. Молекулярна маса білка 18 000. Визначити довжину інформативної частини гена, який кодує цей білок.
Розв'язання.
Оскільки молярна маса однієї амінокислоти дорівнює 100 г/моль, а білка за умовою — 18 000 г/моль, то білок складається із 18 000/100 = 180 (амінокислот). Як відомо, кожну амінокислоту кодують три нуклеотиди, тому полінуклеотид ДНК, що кодує білок, складатиметься із 3 х 180 = 540 (нуклеотидів). Відстань між сусідніми нуклеотидами — 0,34 нм. Довжина гена буде: 0,34 нм х 540 = 183,6 нм.
Відповідь. Довжина інформативної частини гена становить 183,6 нм.
Задача 5. Початкова ділянка молекули білка має такий склад: АСП-ЛЕЙ-АЛА- СЕР-АЛА. Визначити кількісне співвідношення А + Т до Г + Ц (коефіцієнт специфічності) ланцюга ДНК, який кодує цю ділянку білка.
Розв'язання.
Скориставшись таблицею генетичного коду, визначимо структуру ІРНК, за нею, керуючись принципом комплементарності, — матричну ділянку ДНК. Коефіцієнт специфічності розрахуємо на основі формул Чаргаффа.
Білок: АСП - ЛЕЙ - АЛА - СЕР-АЛА
Кодон ІРНК: ГАУ - ЦУУ - ГЦУ - АГУ-ГЦУ
Код ДНК: ЦТА-ГАА—ЦГА-ТЦА—ЦГА
III III III I I I I I I
Г A T - ЦТТ - ГЦТ - АГТ-ГЦТ
A + T = 16, Г + Ц = 14, Ксп ДНК = 16/14 =1,1.
Відповідь. Коефіцієнт специфічності ланцюга ДНК — 1,1.
Задача 6. Гемоглобін крові людини містить 0,34 % феруму. Розрахувати молекулярну масу гемоглобіну.
Розв'язання.
Відносна атомна маса феруму — 56. Використовуючи формулу М = а/Ь х 100 %, визначимо відносну молекулярну масу гемоглобіну:
М = 56/0,34 % х 100 % = 16 471.
Відповідь. Відносна молекулярна маса гемоглобіну — 16 471.
Генна інженерія. Біотехнологія
Генна (генетична) інженерія — сукупність експериментальних методів, за допомогою яких переносять гени з одного організму в інший з метою спрямованого надання останньому нових спадкових ознак. Вона включає такі етапи:
одержання генів шляхом штучного синтезу (хімічного, матричного) або шляхом виділення їх із природних джерел;
включення гена у векторну молекулу ДНК, тобто створення рекомбінантних молекул ДН К;
введення векторної молекули ДНК із включеним геном у реципієнтну клітину;
створення умов для експресії перенесеного гена та його стабільного успадкування;
відбір клітин з діючим перенесеним геном — молекулярне клонування.
Роль векторів виконують плазміди, бактеріофаги, деякі віруси, мітохондріальна
ДНК. Найчастіше векторами слугують плазміди. Плазміди — невеликі кільцеві молекули ДНК бактерій, які здатні до автономної реплікації; вони розташовані в цитоплазмі бактеріальної клітини або інтегровані в її хромосому (ДНК нуклеоїда) і тоді мають назву епісоми. Векторну молекулу ДНК кільцевої форми розрізають ферментами рестриктазами на лінійні фрагменти. Фрагменти векторної ДНК і фрагменти чужорідної ДНК своїми комплементарними («липкими») кінцями здатні об’єднуватися в одну молекулу ДНК. Таку молекулу ДНК називають рекомбі- нантною, або гібридною. Фосфодіефірні зв’язки між нуклеотидами утворюються за допомогою ферментів лігаз. Перенесення необхідних, генів (трансгеноз) забезпечується різними способами — трансформацією (якщо вектор — плазміда), трансдукцією (вектор — бактеріофаг).
Методами генної інженерії одержані трансгенні організми, тобто організми з чужорідними генами. Трансгенні тварини використовують у біомедицині як моделі тих чи інших захворювань людини (миші з геном раку, свині з патологіями серця, які трапляються у людини, корови з людськими імуноглобулінами в крові). Вирішується проблема одержання білків крові людини в молоці трансгенних тварин. Можливі методи лікування спадкових хвороб шляхом перенесення в організм певних генів для виправлення генетичних дефектів розробляє генна терапія.
Своїми успіхами генна інженерія значною мірою зобов’язана створенню банків (бібліотек) генів. Банком генів називають набір генів, одержаний на основі реком- бінантних молекул. Створені банки генів дрозофіли, кишкової палички і багатьох інших організмів, у тому числі і людини. Як читач бібліотеки може вибрати потрібний том книги з необхідними сторінками, так і генетик може відібрати в бібліотеці генів необхідні для дослідження гени за допомогою спеціально розроблених генетичних, біохімічних, радіоізотопних чи імунологічних методів.
Генна інженерія народилася в 1972 p., коли американські генетики П. Берг, Г. Бойер і С. Коен створили першу рекомбінантну ДНК, яка об’єднала у своєму складі генетичний матеріал з трьох різних джерел: повний геном онкогенного вірусу мавпи, частину геному помірного бактеріофага і гени галактозного оперону бактерії кишкової палички. Проте сконструйована рекомбінантна молекула ДНК не була досліджена на функціональну активність, оскільки в авторів виникли побоювання, що методи генної інженерії можуть призвести до створення організмів, небезпечних для здоров’я людини. Втручання в генотип організму може зумовити непередбачувані наслідки для людини, рослин, тварин і довкілля в цілому. Наприклад, бактерія кишкової палички, нешкідлива в звичайних умовах, може перенести онкогенні віруси тварин у кишки людини. На Міжнародній конференції з біобез- пеки в м. Асиломарі (США) у 1975 р. були розроблені правила, дотримання яких усуває ймовірність шкідливих наслідків генної інженерії.
Питання біобезпеки — дуже актуальне. Агенти, які спеціально сконструйовані за допомогою генетичних методів для ураження живих організмів, належать до біологічної зброї. У 1985 р. на міжнародному рівні створена інформаційна робоча група з біобезпеки. Проблеми моралі відносно всього живого, захист людей у генетичних дослідженнях, кодекс лікаря — це перелік лише деяких проблем, які розробляє наука біоетика.
Біотехнологія (від грец. bios — життя, techne — майстер, logos — наука) — наука, яка вивчає застосування живих організмів або біологічних процесів у промисловості з метою одержання біологічно активних речовин (антибіотиків, гормонів, ферментів, вакцин). Сучасна біотехнологія грунтується на застосуванні останніх досягнень генної інженерії. У мікробіологічній промисловості використовують трансгенні штами бактерії кишкової палички, у геном яких методами генної інженерії введені гени людини, що програмують синтез інсуліну, інтерферону, соматотропіну людини. І бактерії в промислових умовах синтезують ці лікарські речовини.
Цікаво знати
Інструкції всередині ДНК клітини, якщо виписати їх, заповнили б тисячу книг у 600 сторінок кожна. Такої дивовижної інформативної компактності на сьогодні не має жодний технічний пристрій. Це зайвий раз підтверджує неперевершеність матінки- природи.
Кожна клітина — це світ, що містить майже двісті трильйонів мізерних атомних груп — молекул. Усім цим мікросвітом злагоджено й оперативно керує генетична інформація ДНК.
Якщо з'єднати «нитки» наших 46 хромосом, то їхня довжина становитиме близько 2 м. Однак ядро, в якому вони знаходяться, має діаметр менше однієї сотої міліметра.
Синтез поліпептидного ланцюга у клітинах відбувається з доволі великою швидкістю. У середньому ця швидкість в еукаріотів сягає близько 2 амінокислот за секунду, а у прокаріотів — близько 15 амінокислот за секунду.
Мікробіологічний синтез — надзвичайно ефективний процес. Так, щоб одержати 5 мг соматотропіну (гормон росту), потрібно використати мозок 500 000 овець упродовж 5 років; аналогічну кількість гормону дають 9 л бульйонної суспензії кишкової палички.
До 1 944 р. генетики вважали, що спадковою речовиною є не ДНК, а білок хромосом. Генетична роль ДНК була встановлена під час дослідження процесів трансформації і трансдукції в мікроорганізмах.
Трансформація — перенесення генетичної інформації від бактерії до бактерії за допомогою ізольованої ДНК, відкрита вперше у 1928 р. англійським мікробіологом Ф. Гриффітсом. Механізм трансформації полягає в тому, що ДНК бактерії-донора адсорбується на поверхні клітини-реціпієнта, проникає всередину клітини, де фрагмент цієї ДНК вбудовується в ДНК донора і починає функціонувати, надаючи організму нових властивостей, які успадковуються.
Трансдукція — перенесення фрагментів ДНК від бактерії до бактерії за допомогою помірних бактеріофагів, відкрита у 1 952 р. у сальмонел. При цьому бактеріофаг, вивільняючись з бактерії, захоплює фрагмент ДНК бактерії-донора і переносить його в бактерію-реціпієнта.
Контрольні завдання до розділу 2
Питання для самоконтролю та обговорення
Які відмінності у молекулярній організації ДНК та РНК?
Які функції ДНК як спадкового матеріалу Вам відомі?
Що таке генетичний код і які його властивості?
Як зрозуміти доцільність множинності генетичного коду?
Що таке ген і які існують його різновиди?
Які відомі мутації і рекомбінації генів?
Що означають терміни «інтрон», «екзон»?
Чим відрізняється мутон від рекона?
Які Ви знаєте етапи біосинтезу білка?
Чому не відбувається репарація РНК?
Чи однаковою є довжина молекул ДНК та РНК, якщо вони складаються з однакової кількості нуклеотидів?
Чи однаковою є молекулярна маса молекул ДНК та РНК, якщо ці молекули мають однакову довжину?
З якою властивістю генетичного коду пов’язане приєднання амінокислоти фенілаланіну до поліпептидного ланцюга, що синтезується на рибосомі, коли в ній з’являється кодон У-У-У?
Чи можуть клітини з однаковим набором генів мати однаковий склад білків?
Який механізм реплікації ДНК і які її основні етапи?
Який механізм елонгації в процесі трансляції?
Які розрізняють форми репарації ДНК?
Контрольно-навчальні завдання
Тести
ТРнк транспортують до рибосом:
Амінокислоти.
Б. АТФ.
ІРНК.
Г. Іони металів.
Ґ. Синтетази.
Змістовні ділянки ДНК хромосом мають назву:
Мутони.
Б. Рекони.
Екзони.
Г. Інтрони.
Ґ. Кодони.
На структурних генах еукаріотів спочатку синтезується молекула про-іРНК, яка потім формується в ІРНК. Цей процес називають:
Трансляцією.
Б. Процесингом.
Термінацією.
Г. Сплайсингом.
Г. Транскрипцією.
Утворення всіх видів РНК у клітині здійснюється:В ядрі.
Б. На рибосомах.
На мембранах ендоплазматичної сітки.
Г. У мітохондріях.
Ґ. У лізосомах.
Скільки водневих зв’язків утворюється між гуаніном і цитозином у молекулі ДНК:
Один.
Б. Два.
Три.
Г. Чотири.
Ґ. Жодного?
Скільки полінуклеотидних ланцюгів має молекула ДНК:
Жодного.
Б. Один.
Два.
Г. Три.
Г. Чотири?
Синтез ДНК відбувається в:
Ядрі
Б. Цитоплазмі.
Апараті Гольджі.
Г. Ендоплазматичній сітці.
Г. Рибосомах.
Процес подвоєння молекули ДНК називають:
Транскрипцією.
Б. Трансляцією.
Трансверсією.
Г. Реплікацією.
Ґ. Репарацією.
Трансгенні організми — це організми:
Що зазнали мутацій.
Б. Гз вродженими аномаліями розвитку.
Одержані шляхом біотехнології.
Г. Створені як біологічна зброя.
Ґ. Із чужорідними генами.
Властивість генетичного коду, що визначає сувору відповідність послідовності триплетів нуклеотидів ДНК послідовності амінокислот поліпептидного ланцюга, — це:
Колінеарність.
Б. Триплетність.
Виродженість.
Г. Неперекривність.
Г. Універсальність.
Вид зв’язку, що забезпечує первинну структуру білка:
Дисульфідний.
Б. Водневий.
Іонний.
Г. Пептидний.
Г. Донорно-акцепторний.
Кодування іРНК лише амінокислоти фенілаланіну відображає таку властивість генетичного коду:
А. Колінеарність.
Б. Специфічність.
В. Виродженість.
Г. Триплетність.
Ґ. Універсальність.
Парою комплементарних нуклеотидів є:
А та Г.
Б. Г та У.
У та Т.
Г. Ц та Г.
Г. Т та Ц.
Особливість нонсенс-кодонів:
Кодують лише по одній амінокислоті.
Б. Кодують по декілька амінокислот.
Для них не існує тРНК.
Г. Вони перекриваються.
Г. У всіх біологічних видів кодують одні й ті самі амінокислоти.
Інверсія в структурі гена — це:
Повернення його ділянки на 180‘.
Б. Переміщення азотистих основ з одного полінуклеотиду в інший.
Заміна однієї комплементарної пари іншою.
Г. Подвоєння ділянки.
Г. Збільшення довжини ділянки у кратному відношенні.
Явище, коли код ДНК змінився, а якісний склад амінокислот і їх послідовність у поліпептиді залишилися незмінними, здатне забезпечитися такою властивістю генетичного коду, як:
Колінеарність.
Б. Специфічність.
Універсальність.
Г. Неперекривність.
Г. Виродженість.
Триплет ДНК — це:
Азотиста основа.
Б. Нуклеотид.
Локус хромосоми.
Г. Генетичний код.
Ґ. Послідовність трьох нуклеотидів.
Задачі та вправи
В одній із ділянок молекули ДНК, що складається з 1818 нуклеотидів, закодовано три білки однакової довжини. Скільки амінокислотних залишків має кожний з цих білків?
Скільки нуклеотидів входить до складу ділянки молекули ДНК, в якій послідовно закодовано інформацію про чотири білки, кожний з яких має 100 амінокислотних залишків?
Яка довжина молекули ДНК і відсотковий вміст азотистих основ у цій молекулі, якщо на її транскрипцію витрачено 2800 молекул аденіну і 6200 молекул урацилу,
що становить 45 % загальної кількості азотистих основ у молекулі РНК, а довжина одного нуклеотиду ДНК — 0,34 нм?
Скільки поліпептидів закодовано в молекулі ДНК, молекулярна маса якої становить 207 х Ю5, якщо кожен з цих поліпептидів має 499 амінокислотних залишків? Молекулярна маса одного нуклеотиду — 345.
Молекула РНК містить 200 нуклеотидних залишків з аденіном і 400 — з урацилом, які в сумі становлять 20 % загальної кількості нуклеотидних залишків РНК. Скільки молекула ДНК, на якій синтезована РНК, містить фосфатних груп та азотистих основ кожного виду?
У хворого на серпоподібно-клітинну (серпастоклітинну) анемію виявлено такий склад амінокислот 4-го пептиду молекули гемоглобіну: ВАЛ-ГІС-ЛЕЙ-ТРЕ-ПРО- ВАЛ-ГЛУ-ЛІЗ. Визначити відсотковий склад нуклеотидів і довжину відповідного фрагмента ДНК.
Молекулярна маса білка становить 100 000. Визначити довжину і масу екзонної ділянки відповідного цистрона.
У двох бактеріях (А і В) в одному і тому самому триплеті А-А-А одного гена аденін замінився на гуанін. Але в бактерії А змінився перший нуклеотид у триплеті, а в бактерії В — третій. Яка можливість зміни структури білка в обох бактеріях у результаті мутації?
Олімпіада для допитливих
Що відбувається з мРНК і тРНК після того, як вони беруть участь у біосинтезі потрібної кількості білка у клітині?
Відомо, що в організмах існують програмний і складний механізми синтезу специфічних білків. Як саме синтезуються в клітині ті речовини, які не мають білкової природи, але також характеризуються видовою специфічністю?
Чи можуть нуклеотиди нуклеїнових кислот рослинного або тваринного походження, що надходять з їжею, споживатися організмом людини без хімічної видозміни? Чи, як й інші біополімери, вони ферментативно розщеплюються до менш складних сполук і засвоюються організмом?
Відомо, що бджоли живляться нектаром, який має вуглеводний склад. Де ж тоді вони беруть власні білки, а також білки, якими вигодовують своїх личинок?
Відомо, що як віруси, так і рибосоми складаються з нуклеїнових кислот і білків. Чому ж тоді віруси здатні до самовідновлення, а рибосоми — ні?
Відомо, що функції ДНК та РНК різняться між собою. Чи існують організми, в яких функції ДНК та РНК однакові?
Відповіді на тести
1А, 2В, ЗБ, 4А, 5В, 6В, 7А, 8Г, 9Ґ, 10А, 11Г, 12Б, 13Г, 14В, 15А, 16Ґ, 17Ґ.
Відповіді на задачі та вправи
Кожен із закодованих білків має по 100 амінокислотних залишків.
Два комплементарних ланцюги ДНК складаються із 2424 нуклеотидів.
Ланцюг ДНК має довжину 6800 нм. %Т = %А = 22,5, %Г = %Ц = 27,5.
У молекулі ДНК закодовано 20 поліпептидів.
У молекулі ДНК є 6000 фосфатних груп, вона містить азотистих основ аденіну і тиміну — по 600, гуаніну і цитозину — по 2400.
%А = %Т = 33,3; %Г = %Ц = 16,7. Довжина ДНК - 8,16 нм.
Довжина становить 1020 нм, маса — 2 070 000.
У бактерій А зміниться структура білка — лейцин замінить фенілаланін, у бактерій В білок не зміниться.
Відповіді на олімпіадні завдання
За допомогою ферментів мРНК і тРНК розщеплюються до нуклеотидів, які знову використовуються для утворення нових РНК, необхідних клітині.
Біосинтез жирів, вуглеводів, пігментів, глікозидів та інших сполук здійснюється за допомогою специфічних ферментів, які мають білкову природу і контролюються відповідними структурними генами. Отже, синтез у клітинах сполук небілкової природи, зрештою, також контролюється генетично.
Можуть споживатися організмом людини без хімічної видозміни, про що свідчать лікарські препарати із вмістом ДНК, РНК рослинного чи тваринного походження.
Бджоли беруть білки та амінокислоти з пилку, який вони збирають з квіток. Уміст білка й амінокислот у пилку сягає 20 %>.
Більш складна організація вірусів забезпечує прикріплення і проникнення їх у клітину, де, використовуючи нуклеотидний склад клітини-хазяїна, й утворюються нові віруси. Рибосоми такої можливості позбавлені.
У багатьох вірусів наявна лише РНК. Під час розмноження вірусів в ураженій клітині вірусні РНК самоподвоюються, збільшується їх кількість, що дає можливість утворюватися новим вірусам.
РОЗДІЛ З
ЗАКОНОМІРНОСТІ УСПАДКУВАННЯ У ЛЮДИНИ ОЗНАК ЗА ЗАКОНАМИ МЕНДЕЛЯ
Чому це актуально?
За сучасними даними експертів Всесвітньої організації охорони здоров'я, із тисячі новонароджених у 42—65 виявляють моногенні захворювання, що мають менделюючий характер.
Серед загальної кількості спадкових хвороб значна частка успадковується відповідно до законів Менделя. Тільки спадкових хвороб обміну речовин сучасна медицина нараховує понад 600. Усі вони переважно є вираженими менделюю- чими ознаками.
Основні поняття генетики
Як відомо, генетика вивчає закономірності спадковості і мінливості. Успадкування — процес передачі спадкової інформації у ряді поколінь. Застосування терміна «успадкування ознак» є умовним, оскільки успадковуються не ознаки, а гени, які несуть інформацію про всі ознаки і властивості організму. Тому для розуміння багатьох спадкових процесів слід опанувати закономірності успадкування (із покоління в покоління) генів.
Генотип — сукупність усіх генів організму (у широкому розумінні — сукупність спадкових факторів). Генотип успадковується від батьківських форм і здатний потенційно визначати ознаки організму.
Фенотип — сукупність усіх зовнішніх і внутрішніх ознак організму. Він є результатом взаємодії генотипу й умов навколишнього середовища. Фенотип ніколи не буває відображенням усього генотипу, а завжди відображаає лише ту частину його, яка реалізується за цих умов, тому, як правило, ознака має певний діапазон прояву (норму реакції), що відображає характер змін факторів середовища.
Алельні гени, алелі (від лат. аііеіоз — протилежний) — парні гени, що займають одні й ті самі локуси гомологічних хромосом і визначають альтернативні взаємо- виключні стани однієї і тієї самої ознаки. Гени, як і хромосоми, парні. У кожній клітині диплоїдного організму будь-який ген представлений двома алелями, один з яких організм одержав від батька, інший — від матері. (Виняток становлять лише статеві клітини — гамети, в яких міститься лише один алель цього гена.)
Таким чином, алельні гени, або гени однієї алельної пари, — це насправді різні форми (різновиди) того самого гена.
Неалельні гени — гени різних алельних пар. Вони, відповідно, визначають варіанти різних ознак і характеризуються протилежними, порівняно з алельними генами, властивостями.
Алельні гени бувають домінантними і рецесивними.
Домінантний ген (алель) — ген, який проявляється у фенотипі гетерозиготного організму (варіант ознаки, що ним визначається, теж є домінантним).
Рецесивний ген (алель) — ген, який не проявляється в фенотипі гетерозиготного організму (визначає, відповідно, рецесивний варіант ознаки).
Домінантний і рецесивний алелі одного гена позначають однаковою літерою латинського алфавіту, але домінантний алель пишуть великою літерою (А), а рецесивний — маленькою (а).
Гомозиготний організм (гомозигота) — диплоїдний організм, в обох гомологічних хромосомах якого містяться однакові алелі певного гена (наприклад АА чи аа). Гомозигота характеризується, відповідно, гомозиготним генотипом. Гомозиготний генотип буває:
гомозиготний домінантний — АА;
гомозиготний рецесивний — аа.
Гомозиготний організм утворює один тип гамет за цим геном (А або а).
Гетерозиготний організм (гетерозигота) — диплоїдний організм, обидві гомологічні хромосоми якого містять різні алелі певного гена (Аа). Гетерозигота характеризується гетерозиготним генотипом — Аа й утворює два типи гамет за цим геном (Ай а).
Гемізиготний організм (від грец. кеті — напів) — організм, генотип якого виражається не двома, а одним апелем, і цей алель завжди фенотипово проявляється, навіть тоді, коли він рецесивний.
Гемізиготними є гаплоїдні організми, а також диплоїдний гетерогаметний організм, але лише за тими генами, які локалізовані у статевих хромосомах. Наприклад, чоловіки (ХУ) є гемізиготними за генами гемофілії, дальтонізму, які локалізовані в негомологічній ділянці Х-хромосоми, оскільки в них лише одна Х-хромо- сома, а У-хромосома цих генів не містить.
Узагальнюючи все вище сказане, розглянемо для прикладу алельну пару, що контролює стан кисті людини:
А — ген полідактилії (багатопалості, найчастіше шестипалості);
а — ген нормальної будови кисті (п’ятипалості).
Теоретично можливими є такі генотипи та фенотипи:
Г енотигг |
АА гомозиготний домінантний |
аа гомозиготний рецесивний |
Аа гетерозиготний |
Фенотип |
Полідактилія |
Нормальна кисть |
Полідактилія |
Закономірності успадкування ознак встановлені Г. Менделем
Закономірності успадкування ознак уперше в 1865 р. відкрив Г. Мендель у дослідах з гібридизації різних сортів посівного гороху. Ці закономірності відомі тепер як три закони (правила) Г. Менделя і закон (гіпотеза) чистоти гамет. Це основні закони генетики.
Суть гібридологічного методу
Успіх сприяв Г. Менделю завдяки вдалому вибору об’єкта досліджень (горох посівний) і застосуванню розробленого ним гібридологічного методу вивчення спадковості, який став основним методом класичної і сучасної генетики.
Його назва зумовлена тим, що вивчення успадкування ознак здійснювалося шляхом гібридизації (схрещування), тобто поєднання двох генетично різних організмів (гамет). Гетерозиготний організм, який утворюється при цьому, називають гібридом, а потомство — гібридним.
Особливостями гібридологічного методу були:
самозапильність гороху, що визначало чистолінійність усіх поколінь (гомозиготність, чистосортність);
відбір для дослідів тих сортів, які різнилися між собою чіткими і зручними для дослідження альтернативними ознаками;
спостереження за успадкуванням не всіх ознак одночасно, як це робили інші вчені, а однієї або кількох пар альтернативних ознак;
ретельне планування дослідів, ведення точного кількісного обліку потомства за кожною парою досліджуваних ознак у ряді поколінь та його математичний аналіз.
За збігом обставин горох має 7 пар хромосом (це було встановлено пізніше) і гени кожної із 7 пар ознак розташовані в різних парах хромосом. Ця обставина допомогла вченому відкрити явище незалежного успадкування ознак.
Моногібридне схрещування — це схрещування, в якому батьківські організми різняться за однією парою альтернативних ознак, дигібридне — за двома, полігібридне — за трьома парами ознак і більше.
Символіка генетичних схем
Заслугою Г. Менделя було також і те, що він запропонував алгебраїчні символи для позначення генетичних схем схрещувань, які з деякими доповненнями застосовують і в сучасній генетиці:
Р (від лат. parentes — батьки) — батьківські форми;
х — схрещування;
Ç — жіноча стать (дзеркало Венери);
З — чоловіча стать (щит і спис Марса);
велика літера латинського алфавіту — домінантний алель;
маленька літера латинського алфавіту — рецесивний алель;
F (від лат. filii — діти) — гібридне покоління;
Fj — перше покоління, F2 — друге і т.д.
У першому рядку схеми записують генотипи батьків (на перше місце переважно ставлять жіночий організм, на друге — чоловічий), у другому — типи гамет, які вони утворюють, у третьому рядку — усі можливі генотипи дітей.
Вивчення успадкування ознак Г. Мендель почав з моногібридного схрещування. Наприклад, схема його досліду для гороху мала такий вигляд:
А — ген жовтого забарвлення насіння;
а — ген зеленого забарвлення насіння.
в
аа
зелені
гомозиготи9 Аа жовті гомозиготи
Гамети
Г,
100 % Гетерозиготи з жовтим насінням
Перший і другий закони Г. Менделя
Наведена схема ілюструє перший закон Г. Менделя — закон одноманітності гібридів першого покоління: при схрещуванні гомозиготних особин, які різняться за однією парою альтернативних ознак, спостерігається одноманітність гібридів першого покоління Р1 як за фенотипом, так і за генотипом.
За фенотипом усі особини мають домінантну ознаку, за генотипом вони гетерозиготні.
Якщо адаптувати цей закон Менделя (ураховуючи його універсальний характер) стосовно успадкування вище розглянутої патогенної ознаки людини (полідактилія), то матимемо:
А — ген полідактилії;
а — ген нормальної будови кисті.
Р
Гамети
Г,
?
АА
гомозиготи-полідактиліки
гомозиготи з нормальною кистю а
X
Аа
100 % гетерозиготи з полідактилією
Цитологічні основи цього закону: гомозиготні батьки (АА х аа) у процесі мейозу утворюють гамети лише одного типу — А і а відповідно. Поєднання цих гамет дає зиготу лише одного типу — Аа.
Наступні закономірності Г. Менделя розглядатимемо також на прикладі патогенних ознак людини. Якщо зобразити схему шлюбу людей з генотипами, що відповідають матимемо:
в
Аа
гетерозиготи-
полідактиліки
А,
а
аа
гомозигота
рецесивна п ’ятипалість
гетерозиготи- полідактиліки
Гамети А, а
АА, Аа, Аа,
Генотип: гомозигота домінантна гетерозиготи
Фенотип: полідактилія
Другий закон Г. Менделя — закон розщеплення ознак: при схрещуванні (шлюбі) двох гетерозиготних особин (гібридів), які різняться за однією парою альтернативних ознак, у потомстві спостерігається розщеплення за фенотипом 3:1 і за генотипом 1:2:1.
За фенотипом 3/4 особин (75 %) мають домінантну ознаку, а 1/4 (25 %) — рецесивну. За генотипом 1/4 особин (25 %) —домінантні гомозиготи АА, 2/4 (50 %) — гетерозиготи Аа, 1/4 (25 %) — рецесивні гомозиготи аа.
Цитологічні основи другого закону: гібриди першого покоління Б, (Аа) при мейозі утворюють яйцеклітини і спермії двох типів — А і а, поєднання яких при схрещуванні дає три типи зигот: АА, аа, Аа. Тому одноманітності в 1 не може бути, бо в основі закону розщеплення лежить феномен незмішування алелів у гібридних організмів.
Закон чистоти гамет
Той факт, що рецесивна ознака, яка була відсутня в гібридів першого покоління Р,, знову з’явилася в 1/4 гібридів другого покоління її2, Г. Мендель пояснив тим, що успадковуються не самі ознаки, а спадкові фактори (гени), які визначають розвиток цих ознак, і що ці фактори дискретні. У гібридів Б, (Аа) є два спадкові фактори (алель- ні гени), один з них А відповідає за домінантну ознаку, інший а — за рецесивну.
Отже, алельні гени, знаходячись у гетерозиготному стані, не зливаються, не змінюють один одного і, не втрачаючи своєї індивідуальності, передаються в гамети. Гамети є «чистими»: вони несуть лише один з двох алелей певного гена.
Ця закономірність була названа гіпотезою «чистоти» гамет, її цитологічну основу забезпечує механізм мейозу, який був відкритий значно пізніше законів Г. Менделя. Оскільки алельні гени знаходяться в гомологічних (парних) хромосомах, а при мейозі в кожну гамету потрапляє лише одна з гомологічних хромосом, відповідно — й один ген з цієї алельної пари.
Аналітичне схрещування
Фенотип організму з рецесивною ознакою цілком визначається його генотипом, оскільки такий організм обов’язково має бути гомозиготним за рецесивним геном (аа), тоді як організм з домінантною ознакою може мати як гомозиготний домінантний генотип (АА), так і гетерозиготний (Аа). Для визначення генотипу в дослідах на рослинах і тваринах проводять аналітичне схрещування і визначають генотип особини за фенотипом потомства.
Аналітичне схрещування — це схрещування, в якому особина, генотип якої невідомий, але його потрібно встановити (АА чи Аа), схрещується з рецесивною гомозиготою (аа). Якщо все потомство від цього схрещування буде одноманітним, досліджуваний організм має генотип АА (схема а). Якщо у потомстві спостерігається розщеплення за фенотипом у співвідношенні 1:1, досліджуваний організм є гетерозиготою Аа (схема б). а)
Р 9 АА * І аа
Гамети А а
Г, Аа
100 %
б)
Р $ Аа * в аа
Гамети А, а а
Г, Аа, аа
50% 50%
Задача 1. У людини полідактилія (шестипалість) — автосомно-домінантна ознака. П’ятипалий чоловік, батьки якого були шестипалі, одружився із шестипалою жінкою, батько якої п’ятипалий. Які діти можуть народитися у цієї шлюбної пари? Визначити генотипи всіх згаданих осіб.
Розв'язання.
Р
Гамети
Е,
А — ген шестипалості а — ген п’ятипалості ^ - Аа
с? — аа
Б, — фенотип?
Р, її — генотипи?
? Аа |
X |
? аа |
А, а |
|
а |
Аа, |
|
аа |
шестипалі |
|
п ’ятипалі |
50 % |
|
50 % |
Відповідь. Генотипи: чоловіка — гомозиготний рецесивний (аа); батьків чоловіка — гетерозиготні (Аа); жінки — гетерозиготний (Аа); батька жінки — гомозиготний рецесивний (аа).
Задача 2. У здорових батьків народилися син, що хворіє на фенілкетонурію, і здорова дочка. Яка вірогідність того, що дочка є носієм гена фенілкетонурії?
Розв'язання.
$ — здорова сГ — здоровий
К — хворий син, здорова дочка
%Р — дочка-носій?
Можливість народження у здорових батьків хворого сина має місце лише тоді, коли ген хвороби рецесивний, а батьки — гетерозиготні носії цього гена, тобто для нашої умови матимемо:
А — ген відсутності фенілкетонурії а — ген фенілкетонурії
Складаємо відповідну генетичну схему:
Р $ Аа х 6* Аа
Гамети А, а А, а
гетерозиготи
Е, АА, Аа, Аа, аа
здорові хворий
дочки (народилась одна) син
Співвідношення генотипів здорової дочки: 1АА:2Аа, тобто вірогідність гетерозиготної дочки-носія становить 2/3, або 66,7 %.
Відповідь. Вірогідність того, що дочка є носієм гена фенілкетонурії, становить 66,7 %.
Третій закон г. Менделя - закон незалежного комбінування ознак
Цей закон Г. Мендель відкрив для дигібридного і тригібридного схрещувань. Щоб зрозуміти суть тих явищ, які мають місце у дигібридному схрещуванні, розглянемо його генетичну схему, моделюючи успадкування патогенних ознак на основі двох алельних пар людини:
А — ген полідактилії; а — ген нормальної будови кисті;
В — ген нормального вуглеводного обміну; в — ген цукрового діабету.
Як відомо, при схрещуванні Менделем гороху батьківські форми були гомозиготами із суто домінантним і суто рецесивним генотипами, тобто батьківські форми людей будуть із такими генотипами й фенотипами:
ААВВ — гомозигота-полідактилік без цукрового діабету; аавв — гомозигота з нормальною кистю і цукровим діабетом.
Р £ ААВВ х в аавв
Гамети АВ ав
Г, АаВв
100 % дигетерозиготи з полідактилією без цукрового діабету
Для отримання Р розглянемо шлюб біологічних особин Б,:
Р $ АаВв х 3 АаВв
Результати злиття гамет зручніше виразити у формі решітки Пеннета:
Гамети 9 |
АВ |
Ав |
аВ |
ав |
АВ |
ААВВ полідактиліки |
ААВв полідактиліки |
АаВВ полідактиліки |
АаВв полідактиліки |
Ав |
ААВв полідактиліки |
ААвв полідактиліки з цукровим діабетом |
АаВв полідактиліки |
Аавв полідактиліки з цукровим діабетом |
аВ |
АаВВ полідактиліки |
АаВв полідактиліки |
ааВВ здорові |
ааВв здорові |
ав |
АаВв полідактиліки |
Аавв полідактиліки з цукровим діабетом |
ааВв здорові |
аавв діабетики |
Оскільки закони Менделя мають статистичний характер, то слід розуміти, що схема ілюструє не кількість фактично народжених дітей у подружньої пари, а лише дає можливість знайти вірогідність певних генотипів чи фенотипів потомства при заданих генотипах батьків.
Отже, у другому поколінні Н. наявне потомство чотирьох фенотипових класів у співвідношенні 9 полідактилію в : 3 полідактиліки з цукровим діабетом : 3 здорових : 1 із цукровим діабетом (9:3:3:1). Нескладно побачити, що за однією, окремо взятою парою ознак, потомство розподіляється приблизно у співвідношенні 3:1, як у моногібридному схрещуванні, тобто таке кількісне співвідношення можна пояснити незалежним розподілом спадкових факторів (генів), що контролюють стан кисті й вуглеводний обмін. З математичної точки зору маємо:
(З + 1) х (3 + 1) = (3 + І)2 = 9 + 3 + 3 + 1.
На цих висновках грунтується третій закон Г. Менделя — закон незалежного комбінування ознак: при схрещуванні гомозиготних особин, які різняться двома (і більше) парами альтернативних ознак, ^ другому поколінні Г, спостерігається незалежне успадкування і комбінування ознак, якщо гени цих ознак розташовані в різних парах хромосом.
Цитологічною основою цього закону є мейоз. У мейозі негомологічні хромосоми розходяться незалежно і можуть комбінуватися в будь-яких поєднаннях. Тому алель А може виявитися в одній гаметі з однаковою вірогідністю з алелем В або з алелем в, але не з алелем а. З такою ж імовірністю алель а може потрапити в гамету з алелем В або з алелем в, але не з алелем А. Гібриди І у мейозі утворюють яйцеклітини чотирьох типів: АВ, Ав, аВ і ав і стільки ж типів сперміїв. При випадкому поєднанні цих гамет можливе утворення 16 типів зигот (4x4), які можна визначити за решіткою Пеннета. Ці типи зигот дають чотири фенотипових класи у співвідношенні 9:3:3:1 і дев’ять генотипових класів у співвідношенні: 1ААВВ : 2АаВВ : 2ААВв : 4АаВв : ІААвв : 2Аавв : ІааВВ : 2ааВв : Іаавв.
Підрахунок розщеплення в Я, за кожною парою ознак показує, що воно в обох випадках дорівнює 12:4, тобто 3:1, як при моногібридному схрещуванні. Отже, дигібридне схрещування — це, по суті, два незалежних моногібридних, які ніби накладаються одне на одного.
Застосовуючи закони Г. Менделя, можна спрогнозувати розщеплення і для більш складних схрещувань: три-, тетрагібридного і т. д. В основі завжди лежатиме розщеплення при моногібридному схрещуванні (табл. 3.1).
Таблиця 3.1. Характеристика типів схрещування
Параметри |
Тип схрещування |
||
Моногібридне |
Дигібридне |
Полігібридне |
|
Кількість типів гамет (ТУ), утворених гібридом |
2і |
22 |
2" |
Кількість комбінацій гамет при утворенні |
4> |
42 |
4" |
Розщеплення за фенотипом у б, |
(3:1)' |
(3:1 )2 |
(3:1)" |
Розщеплення за генотипом у Р2 |
(1:2:1)' |
(1:2:1)2 |
(1:2:1)" |
Кількість різних класів за фенотипом |
2'=2 |
22=4 |
2" |
Кількість різних класів за генотипом |
3'=3 |
32=9 |
3" |
Із кількісних співвідношень гаметоутворення випливає формула:
N = 2",
де N — кількість типів гамет; п — число гетерозиготних алелів генотипу.
Якщо викладені закономірності гаметоутворення адаптувати до дигібридного схрещування, то для можливих класів генотипів отримаємо результати:
Гомозиготні генотипи |
ААВВ |
ААвв |
ааВВ |
аавв |
Гамети (п = 0, N = 1) |
АВ |
Ав |
аВ |
ав |
Моногетерозиготні генотипи |
ААВв |
ааВв |
АаВВ |
Аавв |
Гамети (п = 1, N = 2) |
АВ, Ав |
аВ, ав |
АВ, аВ |
Ав, ав |
Дигетерозиготний генотип |
АаВв |
|||
Гамети (п = 2, N = 4) |
АВ, Ав, аВ, ав |
|||
А
— ген нормального слуху а — ген
глухонімоти В — ген нормального слуху
в — ген глухонімоти 9
—
ааВв в
— Аавв
Р
Гамети
Г,
Аавв
Ав,
ааВв,
глухонімота
75 %
Задача
1. У
людини є дві форми глухонімоти, які
зумовлюють рецесивні алелі різних
генів а і в. Визначити вірогідність
народження нормальної дитини в сім’ї,
де обоє батьків мають різні форми
глухонімоти, а за другою парою алелів
вони гетерозиготні. Розв'язання.
$
ааВв аВ, ав АаВв,
норма
25 %
Аавв,
7сЬІ
—
нормальна дитина/
Відповідь.
Вірогідність народження нормальної
дитини становить 25 %.
Задача
2. Скільки
типів гамет і які саме утворюють особини
з генотипами: 1)
АА;
2) Аа; 3) АаВВ; 4) АаВв; 5) АаввСсОЭ;
6)
ааВВ; 7) АаВвСс?
Розв'язання.
Кількість
типів гамет визначаєм за формулою N =
2П,
де N — кількість гамет, п — число
гетерозиготних алелей:
для
генотипу АА п = 0,
N
=
2°= 1 — один тип гамет: А;
для генотипу Аап = 1, N = 2і = 2 — два типи гамет: А, а;
для генотипу АаВВ п = 1, N = 2'= 2 — два типи гамет: АВ, аВ;
для генотипу АаВв п = 2, N = 22=4 — чотири типи гамет: АВ, Ав, аВ, ав;
для генотипу АаввСсОО п = 2, N = 22 = 4 — чотири типи гамет: АвСО, АвсО, авСО, авсО;
для генотипу ааВВ п = 0, N = 2° = 1 — один тип гамет: аВ;