

Механізм імунного захисту організму
Незважаючи на могутній механізм неспецифічного захисту організму, все ж таки вирішальне значення в підтримці гомеостазу має імунна система, яка ініціює специфічні реакції захисту. Вони в свою чергу поділяються на гуморальні і клітинні.
До гуморальних чинників імунного захисту відносяться імуноглобуліни , яких існує 5 класів (IgM, IgG, IgA, IgE, IgD), і різноманітні цитокіни, що виділяються клітинами організму. Основними чинниками клітинного захисту є Т-ефектори і Т-хелпери різних субпопуляцій. Яким чином розвивається імунна відповідь організму, як реагує він на проникнення збудника? Залежно від того, яким чином антиген проник в організм, залежить його місце перебування в лімфоїдній системі. Проникнувши у тканини, антиген затримується у регіонарному лімфатич ному вузлі. У той же час антигени, що проникли на слизові оболонки дихальних шляхів або кишечника попадають в лімфоїдну тканину слизових оболонок, а антиген, який безпосередньо проникає в кров, затримується у селезінці.
Виділяють такі стадії розвитку імунної відповіді:
Проникнення антигена через бар'єрні тканини.
Зв'язування частини антигена антигенпрезентуючими клітинами; стимуляція антигеном продукції цитокінів клітинами бар`єрних органів, які готують судини і лімфоцити до активації.
Розпізнавання лімфоцитами антигена своїми специфічними рецепторами TcR i BcR.
Взаємодія і проліферація Т- і В-лімфоцитів.
Диференціація (імунопоез) лімфоцитів.
Організація лімфоцитами знищення антигена макрофагами, еозинофілами, базофілами, тучними клітинами, нейтрофілами. Самостійно знищують антиген Т-кілери.
Виведення зруйнованого антигена з організму.
Детальніше цей процес виглядає таким чином. Антиген зв'язують дендритні клітини бар'єрної тканини, наприклад, клітини Лангерганса, і прямують у регіонарний лімфатичний вузол. За час такого переміщення вони частково переробяють антиген до пептидних фрагментів і зв'язують їх з своїми молекулами ГКГ ІІ. Одночасно антиген безпосередньо стимулює інші клітини (кератиноцити, макрофаги), і вони у незначних кількостях починають виділяти хемокіни, фактор некрозу пухлин, які активують ендотелій у місці попадання антигена. На ендотелії появляються молекули адгезії для лімфоцитів і лейкоцитів.
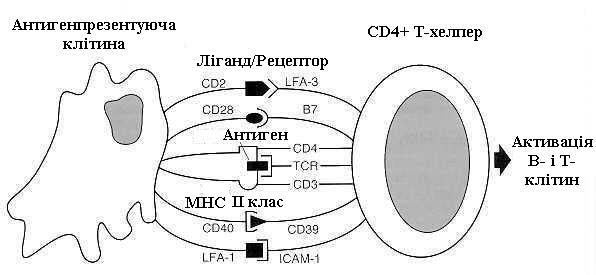
У лімфатичному вузлі дендритні клітини подають антиген для розпізнання Т- і В-лімфоцитам (рис. 7.23). Процес розпізнавання здійснюється за рахунок взаємодії між молекулами мембрани лімфоцита і молекулами мембрани антигенпрезентуючої клітини: рецептор Т-лімфоцита TcR реагує з антигеном; CD4 Т-хелпера з антигеном ГКГ ІІ класу або CD8 Т-ефектора зв'язується з антигеном ГКГ І класу; має місце взаємодія і інших важливих молекул. Тільки після цього може розпочатись проліферація Т-лімфоцитів і секреція ними проліферативних цитокінів (рис. 7.24).
|
Рис. 7.23. Презентація антигену Т-хелперу. |
|
Рис. 7.24. Схема розпізнавання антигену: CD2, 3, 4,28,39,40 B7, TCR, LFA-1, LFA-3, ICAM - рецептори на поверхні клітини. |
Клони Т- і В-лімфоцитів, які зв'язали антиген, починають взаємодіяти між собою, і Т-хелпери синтезують власні фактори росту, наприклад, ІЛ-2. Щоб Т-лімфоцит почав продукувати цито кіни, які зумовлюють диференціацію В-лімфоцитів, необхідна взаємодія мембранної молекули Т-хелпера CD40L з молекулою CD40 на мембрані В-лімфоцита, який теж розпізнав цей антиген. Після цього В-лімфоцити швидко проліферують . Проліферація є характерною ознакою імунної відповіді. У результаті проліферації розростаються тільки специфічні клони лімфоцитів, що зумовлюють багатократне збільшення клітин, спрямованих своєю активністю на антигени.
Лімфоцити антигенрозпізнавальних клонів після проліферації диференцюються згідно спеціалізації, набутої ще в лімфопоезі. У результаті диференціації Т-хелпери, Т-ефектори, В-лімфоцити у залежності від наявності на їх мембрані певних homing-рецепторів мігрують із лімфатичного вузла в ті місця тканини, де перебуває антиген. Плазмоцити (диференційовані В-лімфоцити) починають секретувати у кров імуноглобуліни, які необхідні для зв`язування антигена, який там перебуває. У місцях найбільшого скупчення антигена активовані лімфоцити залучають своїми цитокінами нейтрофіли, макрофаги, еозинофіли, тучні клітини, базофіли, які здатні переробити, зруйнувати антиген і вивести його з організма.
Після виведення антигена з організму імунна відповідь на нього припиняється. Цьому сприяє відсутність антигена; із активованих CD4+Т-лімфоцитів розвивається субпопуляція Th3, яка виділяє, серед інших цитокінів, велику кількість супресорного цитокіна TGFb, який припиняє проліферацію лімфоцитів і продукцію запальних цитокінів; активовані лімфоцити гинуть за механізмом цитоптоза.
Частина лімфоцитів активно захищається від апоптозу, у них припиняється продукція цитокінів, і вони здатні тривалий час циркулювати в організмі. Це - клітини імунологічної пам'яті. При повторному попаданні антигена в організм його зустріне значна кількість вже диференційованих клітин антигенспецифічного клону. На цьому принципі і базується сучасна вакцинація населення.
На початку нашого викладу йшлось про те, що в імунному захисті велика роль належить імуноглобулінам і цитокінам. Імуноглобуліни ми розглядали у першій лекції. Тепер потрібно ознайомитись з основними цитокінами. З допомогою цитокінів лімфоцити взаємодіють між собою і одночасно з клітинами інших тканин організму, які теж здатні ви діляти значну кількість цих біологічно активних речовин і у свою чергу впливають на лімфоцити. Цитокіни є тим містком, який зв`язує систему імунітету із цілим організмом.
На даний час за своєю функцією цитокіни ділять на 5 груп. 1.Цитокіни, які зумовлюють природний захист. Вони виробляються макрофагами, кератиноцитами та іншими клітинами у результаті безпосереднього контакту з мікроорганізмами. До них відносять: a- і b-інтерферони, ІЛ-1, ІЛ-6, хемокіни, фактор некрозу пухлин. 2. Цитокіни, які регулюють ріст, активацію і диференціацію лімфоцитів. (ІЛ-2, ІЛ-4, TGFb). Їх головними продуцентами є Т-лімфоцити. Цитокіни починають синтезуватись після того, як рецептор Т-лімфоцита TcR прореагує з антигеном.
3. Цитокіни, які активують клітини запального інфільтрату. Найчастіше їх продукують імунні лімфоцити з метою залучення у вогнище з антигеном клітин запалення - нейтрофілів, макрофагів, еозинофілів, які повинні знищити і видалити цей антиген. У цю групу входять: g-інтерферон, ІЛ-5, ІЛ-10, ІЛ-12, лімфотоксин. g-інтерферон найактивніший стимулятор макрофагів, що активує ПК (природні кілери), нейтрофіли, ендотелій судин, запускає диференціацію CD4+Т-лімфоцитів у Th1 і CD8+ у Т-ефектори; у В-лімфоцитах пригнічує перемикання синтезу імуноглобулінів на IgG1 i IgE.
4. Цитокіни - фактори росту. Вони стимулюють проліферацію і диференціацію клітин попередників лейкоцитів у кістковому мозку (ІЛ-3, ІЛ-7, ІЛ-9, ІЛ-11,G-CSF та інші ростові фактори).
5. Мембраноасоційовані цитокіни - стимулюють ангіогенез. Як правило цитокінів не можна виявити за виключенням важких септичних станів, коли там можуть появитись ІЛ-1, ІЛ-6, фактор некрозу пухлин. Інші цитокіни ніколи у кров не попадають, а завжди діють локально у місці їх виділення. Цитокіни синтезуються оперативно, коли клітини отримують відповідний сигнал. Зрозуміло, що вони діють тільки на ті клітини, на мембранах яких розташований рецептор, що може зв`язати цей цитокін.
Серед цитокінів є значна кількість інтерлейкінів. Основні з вивчених інтерлейкінів подані нижче.
Таблиця 6
Імунологічна дія основних інтерлейкінів.
Інтерлейкіни |
Походження |
Імунологічна дія |
ІЛ-1 |
Макрофаги, Моноцити |
Посилює диференціацію і дію В- і Т- лімфоцитів; стимулює ріст і активність природніх кілерів |
ІЛ-2 |
Активовані Т-хелпери |
Стимулює ріст Т- клітин; допомагає в дифереціації В- клітин; стимулює NK- клітини |
ІЛ-3 |
Т-хелпери |
Стимулює ріст і диференціацію макрофагів, гранулоцитів, базофілів і тучних клітин |
ІЛ-4 |
Т-хелпери, макрофаги |
Допомагає в рості В- клітин і у диференціації Т- хелперів; стимулює синтез антигенів ГКГ на В- клітинах і макрофагах; разом із ІЛ-3 стимулює алергоцити |
ІЛ- 5 |
Т-хелпери, алергоцити |
Допомагає в диференціації В клітин і стимулює ріст і диференціацію еозинофілів, активує їх дію і хемотаксис |
ІЛ- 6 |
Макрофаги, моноцити, Т- клітини |
Стимулює дозрівання мегакаріоцитів і підвищує кількість тромбоцитів, посилює ріст і диференціацію Т- і В- клітин і продукцію Ig |
ІЛ -7 |
Клітини кісткового мозку |
Допомагає в рості недозрілих В-лімфоцитів; підвищує розмноження Т- клітин; стимулює цитотоксичну дію Т- клітин |
ІЛ- 8 |
Мононуклеарні клітини, фібробласти, епітеліальні клітини |
Хемотаксичний ефект на гранулоцити; стимулює,. виділення лізосомних ензимів; обумовлює адгезію нейтрофілів до ендотелію |
ІЛ- 9 |
Активовані Т- клітини |
Допомагає в дозріванні клітин кісткового мозку; стимулює ріст алергоцитів |
ІЛ- 10 |
Т-хелпери, В- лімфоцити |
Сильно пригнічує активність макрофагів; гальмує продукцію |
ІЛ- 11 |
Фібробласти |
Разом із ІЛ- 3 стимулює ріст мегакаріоцитів |
ІЛ- 12 |
В-лімфоцити, макрофаги |
Сприяє диференціації Т- хелперів; стимулює ріст і функції NK і Т- клітин |
Але у специфічній імунній відповіді, особливо на внутріклі тинну (вірусну) інфекцію крім гуморального механізму захисту, спрацьовують клітинні механізми. Основними діючими особами в даному випадку являються Т-хелпери і цитотоксичні Т-лімфоцити. CD8+ Т-кілери розпізнають своїм TcR рецептором антигени вірусів, деяких бактерій і найпростіших, що знаходяться на уражених соматичних клітинах, а їх молекули CD8 реагують з антигеном ГКГ І класу цих клітин. Приєднуючись до інфікованих клітин, вони виділяють всередину клітини-мішені перфорин, який руйнує мембрану, що зумовлює лізис клітини разом з вірусами, які там знаходились.
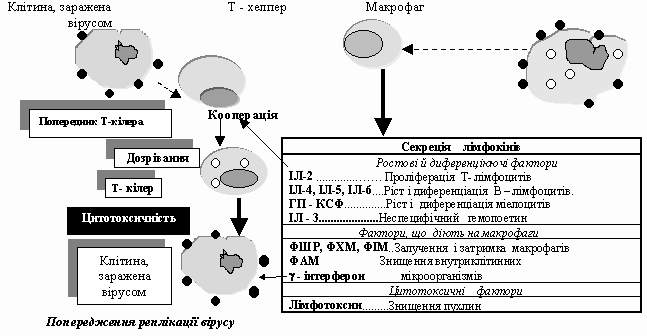 Схема
3.
Механізм противірусного захисту.
Схема
3.
Механізм противірусного захисту.
Тому, що CD8 молекула комплементарна антигену ГКГ І класу, то Т-кілер може знищити будь-яку клітину уражену збудником, адже всі клітини організму несуть на собі антигени ГКГ І класу.
Коли в клітинах знаходяться бактерії або найпростіші ситуація дещо інша. Лізис макрофагу, в якому знаходитиметься збудник, призведе до вивільнення мікроорганізму без його знищення. Тому в даному випадку підключаються інші Т- клітини - Th1 (Т-хелпери гіперчутливості сповільненого типу). Вони розпізнають інфікований макрофаг за присутністю на його поверхні мікробного антигену в комплексі з білком ГКГ ІІ класу. При цьому Т-клітини виділяють лімфокіни, які стимулюють внутриклітинне знищення збудника макрофагом. Таким чином, ми бачимо, що основною регулюючою клітиною, як гуморальної, так і клітинної імунної відповіді, є Т-лімфоцит хелпер. Без нього не можлива активація ні В-лімфоцитів, ні Т-кілерів. Якщо ж згадати, що при СНІДі в основному уражуються клітини, які мають СD4 молекули- антигени (Т-хелпери), то стає зрозумілим - страждає вся система імунного захисту організму. Організм стає обеззброєним, безпорадним проти бактерій і вірусів. Ось чому такою небезпечною є ця важка недуга. Клітинами, які володіють протилежною функцією в імунній відповіді в порівнянні з Т-хелперами, є супресорні клітини. Ви знаєте, що існують різні механізми супресії, пригнічення імунної відповіді. Це абсолютно природньо, адже, якщо природа створила клітини, що посилюють імунну відповідь, то вона повинна була створити і таку систему, яка би тримала в певних рамках Т-хелпери, щоб це посилення імунних реакцій не було безмежним. При значному розвитку імунної відповіді організму активуються супресорні механізми, які пригнічують Т-хелпери (Th0,Th1,Th2). Відповідно зменшується вплив останніх на В-лімфоцити і Т-ефектори- попередники Т-кілерів. Імунологічна відпо відь послаблюється. При певній межі такого пригнічення включається інша система клітин - система клітин контрсупресорів, які конкурують з супресорами і стримують їх вплив на Т-хелпери. І знову рівень імунної відповіді починає підвищуватись. Ось таким чином відбувається регуляція імунної відповіді.