

Транспорт электронов и фотофосфорилирование
В фотосистемах I и II поглощение двух световых квантов (фотонов) хлорофиллом и другими пигментами приводит к возбуждению электрона в реакционном центре (например, Р700 в случае фотосистемы I; см. рисунок). Обладающий достаточно высокой энергией электрон включается в цепь электронного транспорта. Затем электрон «падает» на низший энергетический уровень (этот процесс изображен на рисунке как повышение значения стандартного электродного потенциала Е°'). В фотосистеме I электрон в конечном счете используется для восстановления NADP+, а возбужденные электроны фотосистемы II передаются фотосистеме I, причем в процессе их транспорта каждая пара возбужденных электронов участвует в фосфорилировании одной молекулы ADP. Таким образом, общая стехиометрия реакций, индуцируемых фотонами в фотосинтезирующих эукариотах, выглядит следующим образом:
H 2O + 4hν + NADP+ + ADP + Pi NADPH + Н+ + ½О2 + (ATP+H2О)

Рисунок - Инициированное поглощением света возбуждение электронов
(вертикальные стрелки) и их транспорт в фотосинтезирующих системах.
Поглощенная световая энергия может участвовать в регенерации дополнительных количеств АТР по пути циклического фотофосфорилирования, изображенному на рисунке штриховой линией. В этом случае возбужденные электроны транспортируются от Р700 к Р430, затем к цитохрому b563 и снова к Р700; на последнем этапе этого пути электроны принимают участие в фосфорилировании ADP по следующему простому уравнению:
3 hν
+ ADP
+ Pi
АТР + Н2О
hν
+ ADP
+ Pi
АТР + Н2О
Изменение свободной энергии в реакции фотосинтеза равно +686 ккал/моль. Поскольку в любой из фотосистем для возбуждения одного электрона необходимы два фотона и поскольку генерирование одной молекулы О2 сопровождается переносом четырех электронов, то образование 6О2 требует поглощения 48 квантов света. Согласно уравнению для расчета Ер, энергия фотонов зависит от длины их волны. Если принять λ = 700 нм, то минимальное количество световой энергии, необходимой для инициирования реакции фотосинтеза с помощью фотосинтезирующей системы растений, составит 1968 ккал/моль глюкозы. Следовательно, в первом приближении эффективность утилизации поглощенного света будет равна 686 : 1968 = 0,35 = 35%.
Темновые реакции фотосинтеза
В зеленых растениях и фотосинтезирующих микроорганизмах существует фермент, катализирующий включение СО2 в карбоксильную группу 3-фосфоглицериновой кислоты, которая затем по пути глюконеогенеза преобразуется глюкозо-6-фосфат, а затем в глюкозу. Этот фермент, названный дифосфорибулозокарбоксилазой или рибулозодифосфаткарбоксидисмутазой, катализирует карбоксилирование и гидролитическое расщепление рибулозо-1,5-дифосфата с образованием двух молекул 3-фосфоглицериновой кислоты.
Однако участием одной только этой последовательности реакций нельзя было объяснить тот факт, что все шесть атомов углерода гексозы при фотосинтезе образуются в конечном счете из С02. Кальвин предложил циклический механизм синтеза гексоз, при котором одна молекула рибулозо-1,5-дифосфата регенерируется при восстановлении одной молекулы СО2.
Суммарное уравнение этого сложного цикла имеет следующий вид:
6 Рибулозо-1,5-аифосфат + 6С02+18АТФ + 12НАДФ-Н+12Н+
6 Рибулозо-1,5-дифосфат + Гексоза + 18ФН + 18АДФ + 12НАДФ+.
Рибулозо-1,5-дифосфат фигурирует в обеих частях уравнения только для того, чтобы показать, что он является необходимым компонентом и регенерируется в конце цикла. Суммарное уравнение (после сокращения рибуло-зо-1,5-дифосфата) имеет следующий вид:
6 С02+18АТФ
+ 12НАДФ • Н + 12Н+
Гексоза
+ 18ФН
+ 18АДФ + 12НАДФ+
С02+18АТФ
+ 12НАДФ • Н + 12Н+
Гексоза
+ 18ФН
+ 18АДФ + 12НАДФ+
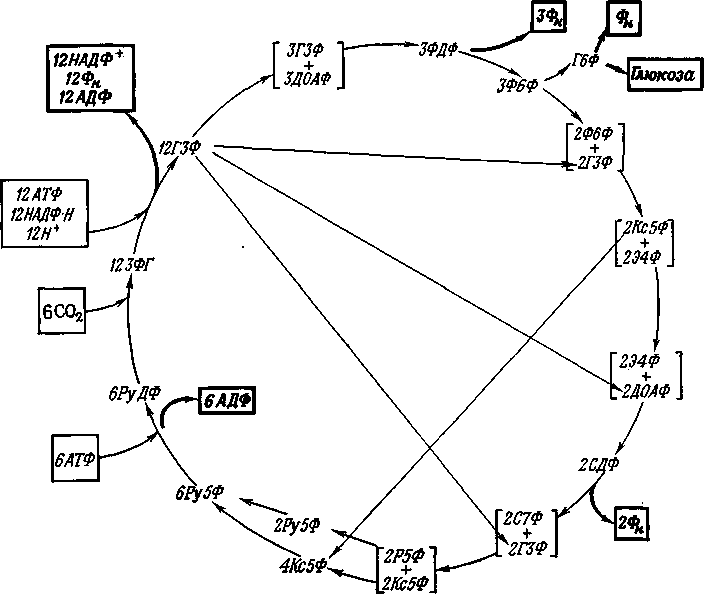
Последовательность реакций цикла Кальвина:
1 )
6СО2
+ 6
рибулозо-1,5-дифосфат
12
3-фосфоглицерат,
)
6СО2
+ 6
рибулозо-1,5-дифосфат
12
3-фосфоглицерат,
2 ) 12 3-фосфоглицерат + 12 АТФ 12 1,3-дифосфоглицерат
3) 12 1,3-дифосфоглицерат + 12НАДФ-Н + 12Н
12 глицеральдегид-3-фосфат + 12НАДФ+
4 ) 5 глицеральдегид-3-фосфат 5 диоксиацетонфосфат
5 )
3
глицеральдегид-3-фосфат + 3 диоксиацетонфосфат
)
3
глицеральдегид-3-фосфат + 3 диоксиацетонфосфат
3 фруктозо-1,6-дифосфат
6) 3 фруктозо-1,6-дифосфат 3 фруктозо-6-фосфат + 3Фн
7 ) Фруктозо-6-фосфат Глюкозо-6-фосфат
8 ) Глюкозо-6-фосфат Глюкоза + Фн
9 )
2
фруктозо-6-фосфат + 2
глицеральдегиц-З-фосфат транскетолаза
)
2
фруктозо-6-фосфат + 2
глицеральдегиц-З-фосфат транскетолаза
Ксилулозо-5-фосфат + 2 эритрозо-4-фосфат
1 0) 2 эритрозо-4-фосфат + 2 диоксиацетонфосфат альдолаза
2 седогептулозо-1,7-дифосфат
1 1) 2 седогептулозо-1,7-дифосфат фосфатаза
2 седогептулозо-7- фосфат + 2ФН
12) 2 седогептулозо-7-фосфат + 2 глицеральдегид-3- фосфат транскетолаза 2 рибозо-5-фосфат + 2 ксилулозо-5-фосфат
13) 2 рибозо-5-фосфат изомераза 2 рибулозо-5-фосфат
1 4)
4
ксилулозо-5-фосфат эпимераза
4
рибулозо-5-фосфат
4)
4
ксилулозо-5-фосфат эпимераза
4
рибулозо-5-фосфат
1 5)
6
рибулозо-5-фосфат + 6АТФ фосфорибулокиназа
6
рибулозо-1,5-дифосфат
+ 6АДФ
5)
6
рибулозо-5-фосфат + 6АТФ фосфорибулокиназа
6
рибулозо-1,5-дифосфат
+ 6АДФ
Реакции (1) — (8) - реакции образования глюкозы из СО2 и рибулозо-1,6-дифосфата. Реакции (9) — (15) связаны с регенерацией рибулозо-1,5-дифосфата.