

Покриви тіла Здатність живих істот до життя в різних умовах є результатом еволюції не тільки їх самих, але і їх систем органів. Нижче ми розглянемо цей висновок на прикладах еволюції систем органів тварин, починаючи з найпростіших і закінчуючи ссавцями. Зовнішні покриви тварин (шкіра або интегумент) виконують захисну функцію, захищають тіло від зайвої втрати води і допомагають регулювати температуру тіла. Крім того, у ряду організмів є додатковим органом, який приймає участь в обміні речовин (диханні і виділення). У безхребетних покриви тіла не досягають значного розвитку, будучи представленими ектодерми і її похідними. Еволюція покривів у них йшла у напрямку розвитку миготливого епітелію в плоский епітелій, що відзначається у турбеллярий та інших плоских хробаків, а також у круглих і кільчастих хробаків. Миготливий епітелій служить примітивним органом руху, тоді як плоскі цією здатністю не володіють. У членистоногих поверхневий шар епітелію перетворюється в хітинізовані кутикулу, яка розвивається в панцир у ракоподібних. У молюсків поверхневий шар епітелію розвивається в раковину, що містить вапно. У тварин окремих груп (кільчасті черви, членистоногі та молюски) в епідермісі локалізовані одно-або багатоклітинні залози (слинні, павутинні, отруйні та інші). У хордових еволюція шкірних покривів йшла в напрямку заміни одношарового епітелію багатошаровим, зокрема, у формуванні двох шарів і в превалирующем розвитку власне шкіри (коріумі). Шкіра хордових складається з поверхневого шару ектодермального походження, або епідермісу, іноді званого кутикулою, і нижнього шару мезодермального походження - власне шкіри (коріумі), або дерми. У тунікат (асцидій, піросів та інших морських тварин) наружний покрив є драглисту або хрящову оболонку, що є продуктом ектодермального епітелію і пронизану мезодермальних клітинами і судинами. Покрови ланцетника мають Двошарове будову. Перший шар представлений циліндричним одношаровим епітелієм ектодермального походження, другий - драглистої сполучною тканиною, яка виконує роль коріумі. Покрови у хребетних досягли великого розвитку і являють собою шкіру, яка побудована з двох різних шарів. Один з них є поверховим і складається з багатошарового епітеліального епідермісу ектодермального походження, другий - нижнім, власне шкірою, яка прилягає до підшкірної тканини. Остання, у свою чергу, прилягає до м'язів і кісток. Оскільки у земноводних шкіра використовується в якості додаткового органу дихання, але при цьому необхідна водна плівка, то це призвело до розвитку в шкірі цих тварин багатошарових залоз, які продукують слиз і зволожуючих її. Шкіра земноводних містить також отруйні залози, що виконують захисну функцію. У ссавців розвинулися похідні шкіри (поверхневий каратин, волосся, копита і роги), які виконують важливі функції в житті організмів. Велике значення в терморегуляції мають шкірні потові залози.
Скелет
Скелет виконує опорну і захисну функції. У безхребетних скелетом є ригідні структури, які мають найрізноманітніший характер і підрозділяються на ендоскелет і екзоскелет Ендоскелет побудований з органічних фібрил або кристалів неорганічних сполук. У найпростіших ендоскелетной структурою є аксостіль ряду джгутикових, який має фиб-ріллярную структуру, у радіолярій і феромініфер ендоскелет представлений кристалами неорганічного походження, у губок - спікулами (голками, що складаються з відкладень карбонату кальцію або силікону навколо клітинних фібрил), у морських зірок - різними хрящеподобной тканинами. Деякі членистоногі мають добре розвинений ендоскелет, до якого прикріплюються м'язи. Вапняні платівки та голки иг-локожіх мають мезодермаль-ное походження. Екзоскелет є у багатьох безхребетних організмів і характеризується надзвичайною різноманітністю, починаючи від пеллікули (потовщеною клітинної стінки) і закінчуючи такими утвореннями, як крила метеликів. Імпрегнація ек-зоскелета неорганічними солями підвищує його ригідність. Гнучкість екзоскелета, яка необхідна для його мобільності, досягається у багатьох нематод внаслідок гнучкості кутикули, а у коловерток і особливо у членистоногих завдяки наявності гнучких районів між ригідними ділянками кутикули. У членистоногих кутикула є результатом секреції клітинного епідермісу і складається з тонкої зовнішньої епікутикули і товстої ендокутикулу, до складу якої входять хітин і білок. У нижчих хордових розвинений осьовий скелет, яким є хорда. У безчерепних ендоскелет є хорда, а екзоскелетів - стержневідние освіти, що складаються з щільного драглистого речовини (непарні плавці й опора зябрового апарату). В круглоротих хорда також зберігається впродовж всього онтогенезу. Однак у них під хордою вже з'являються парні закладки хребців у вигляді хрящоподібних утворень (верхні дуги). У безхребетних кальцієвий скелет представлений кальцитом. У хребетних ендоскелет досягає високого розвитку, будучи хрящовим і (або) кістковим скелетом. У цих організмів розрізняють осьовий скелет, скелет голови (черепна коробка і вісцеральний скелет) і скелет кінцівок. Екзоскелет представлений волоссям, пір'ям, лускою, рогами, відростками рогів, копитами, кігтями, кісточками в шкірі. У примітивних риб поряд з верхніми дугами з'являються закладки нижчих дуг. У вищих риб з тканин, які оточують хорду, і підстав верхніх і нижчих дуг, розвиваються хребці. Хребетний стовп складається з туловищного і хвостового відділів. Завдяки зрощенню решт верхніх дуг утворюється канал для спинного мозку. До нижчих дугам прикріплюються ребра. У земноводних процес заміни хорди хребцями поглиблюється ще більшою мірою і відбувається в ранні періоди розвитку. Розвиваються чотири відділи хребта - шийний, грудний, крижовий і хвостовий. Шийний відділ дуже короткий (один хребець), грудний відділ налічує п'ять хребців, до яких прикріплюються вільно закінчуються ребра (не доходять до грудини), крижовий відділ також складається з одного хребця, до якого прикріплюються кістки тазу. Хвіст хвостатих амфібій складається з декількох хребців, безхвостих - з одного хребця, званого уростілем. У амніот (рептилій, птахів і ссавців) розрізняють осьовий скелет, скелет голови (череп) і скелет кінцівок. Осьовий скелет складається з шийного, грудного, поперекового, крижового і хвостового відділів. Кількість шийних хребців варіює від 1 (у сучасних земноводних) до 76 (у викопних форм - морських плезіозаврів). У мніот і лабірінтодонтов перші два шийні хребці спеціалізовані, забезпечуючи підтримку і рух голови. Кількість шийних хребців залежить від довжини шиї. У деяких рептилій їх кількість досягає 8, у птахів - до 25. Сучасні ссавці мають 7 шийних хребців. Кількість грудних хребців різна в різних груп хребетних. У черепах 10 грудних хребців злиті зі зв'язками і кістками і формують панцир, у плазунів до грудних хребців прикріплюються ребра, що формує грудну клітку. Птахи мають по 4-6 грудних хребців. У ссавців частіше розвивається 10-20 грудних хребців. У людини грудну клітку формують також ребра (12 пар) і грудина. Поперековий відділ розвинений по-різному. У рептилій поперековий відділ виражений погано. У птахів поперекові хребці зазвичай злиті з грудними, крижовими і урокрестцовимі, формуючи складний криж. У ссавців кількість поперекових хребців становить від 3 до 24 в залежності від виду. Кількість хвостових хребців залежить від довжини хвоста. У жаб, наприклад, хвостові хребці зливаються в одиночну кістка (уростіль), у птахів є 4-6 хвостових хребців, які зливаються також у уростіль. Деякі викопні птиці (археоптерикс) мали довгий хвіст, що складається з 20 хребців. У ссавців кількість хребців у хвості залежить від виду. Прямоходіння людини вело до викривлення хребта, тобто це виявилося «платою» за Прямоходіння Скелет голови складається з черепа і вісцерального (лицьового) скелета. Черепна коробка розрізняється не тільки за формою, а й за кількістю вхідних в неї кісток, які дуже коливаються в залежності від приналежності тварин до того або іншого класу. У кісткових риб, наприклад, кількість кісток черепа перевищує 100, у той час як у земноводних воно дорівнює 15-19, у плазунів - від 40 до 70, у птахів - 20, у примітивних ссавців (Didelphis) - 48, у шимпанзе - 22 , у людини - 27 Зниження кількості кісток черепа в еволюції від риб до ссавців є результатом втрати індивідуальних кісток або їх злиття з іншими кістками. Еволюція вісцерального скелета йшла в напрямку його редукції. Він дуже зменшений у чотириногих хребетних, дихають легенями. Первісна функція вісцеральних дуг полягала у підтримці жабер. Це підтверджується і наявністю зябрових щілин у нижчих риб (кроссоптеріг, осетрових та ін.) У рептилій і птахів перша зяброва дуга розвинулася в скелет гортані. Зяброва кришка, яка представляла собою кістковий клапан, що покриває зябра, і більшість жабер були загублені наземними хребетними. Багато кісток мозкового черепа, які розділені у нижчих чотириногих, злилися у ссавців. Ссавці відрізняються від інших тварин наявністю трьох невеликих кісток в порожнині середнього вуха. Одна з цих кісток - стремінце - гомологічна другий вісцеральної дузі риб, інша - ковадло - гомологічна кістки нижньої щелепи інших хребетних, а третя - молоточок - гомологічна кістки верхньої щелепи інших хребетних. Скелет кінцівок має особливості, причому ключовим є питання про перехід плавців у кінцівки. Кінцівки класифікують на непарні і парні. Прикладом непарних кінцівок є спинний і хвостовий плавці у риб, парних - вільні кінцівки, роль яких виконують плавці у риб і п'ятипалі кінцівки у наземних тварин. Парні плавці кистеперих риб містять гомологи плечової, ліктьової і променевої кісток пятипалой кінцівки наземних хребетних, що свідчить про походження пятипалой кінцівки від плавця кистеперих риб. Вільні кінцівки чотириногих зазвичай складаються з трьох сегментів, які організовані по-різному у різних тварин. Головні варіації відзначаються в будові кісток плеча і стегна. Зокрема, плечова і стегнова кістки вкорочені у дельфінів і деяких копалин форм (іхтіозаврів). Істотні коливання відзначаються в кількості кісток зап'ястя і плесна. Деякі з примітивних копалин земноводних мали від 20 до 30 кісток у цих частинах скелета, але сучасні земноводні мають лише від 3 до 9 в зап'ястя і від 3 до 12 в плюсне. Число кісток в кожному пальці також коливається в різних хребетних. Фалангових формула (кількість кісток від першого пальця до останнього) у сучасних амфібій, у яких є зазвичай чотири пальці, складає 2-2-3-2 (саламандри), 2-2-3-3 (жаби). Іхтіозаври мали від трьох до восьми пальців. Крила сучасних птахів мають по три пальці (знижений число фаланг). Загальна кількість кісток у людини становить 206. У ході еволюції сформувався механізм остеофікаціі скелета ембріонів, в якому істотне значення мають кальцій,фосфор, вітаміни Д і ряд ферментів. У хребетних кальцієвий скелет представлений не кальцитом, а гідроксиапатиту. У ході еволюції істотний розвиток придбала і скелетна мускулатура. Виробився також механізм використання хімічної енергії їжі для скорочення м'язів (20-35% від загальної кількості енергії) і для використання решті енергії як тепла, що забезпечує тепловий режим організму.
Травна система
У найпростіших органами травлення є травні вакуолі. Для цих тварин характерно внутрішньоклітинне травлення, яке є найдавнішим. У кишковопорожнинних і черв'яків з ембріонального бластопора розвивається ротовий отвір. У кишковопорожнинних рот відкривається в гастральную порожнину, яка є порожниною первинної кишки і закінчується сліпо (анального отвору немає). У цій порожнині поряд з внутрішньоклітинним травленнямз'являється позаклітинне. На більш високих рівнях еволюції внутрішньоклітинне травлення втрачається повністю, поступаючись місцем позаклітинного (порожнинної) як більш прогресивному способу. Однак внутрішньоклітинне травлення все ж таки зберігається в організмів на всіх щаблях розвитку, але лише у вигляді фагоцитозу, що виконує захисну роль. У війчастих і плоских хробаків будова травної системи ще зберігає схожість з травною системою кишечнику-нополостних. Анального отвору немає, тому викид травних залишків відбувається через рот. Але у круглих хробаків травна система вже дещо ускладнюється, оскільки травна трубка помітно підрозділяється на передній, середній і задній відділи, з яких передній і задній відділи мають ектодермальне походження, середній - ентодермальноє. Вперше розвивається анальний отвір за допомогою впячивания ектодерми в каудальную частина тіла і з'єднання її з порожниною первинної кишки. У кільчастих хробаків травний тракт вже представлений глоткою, стравоходом, зобом, шлунком, середньою та задньою кишками. Подальше ускладнення травного тракту триває у членистоногих та молюсків, у яких розвиваються ротові придатки навколо рота, які служать для подрібнення їжі, в кишечнику розвиваються відростки, що збільшує його поверхню. Розвиваються травні залози, які є аналогом печінки хребетних. У хордових в ході еволюції триває диференціація кишкової трубки, причому більш глибока, в результаті чого відбувається збільшення кишкової поверхні для всмоктування продуктів травлення. Виникають залози, що беруть участь у травленні. Травна система всіх хордових в основному має ентодермальноє походження (крім ротового відділу і заднього кишечника, мають ектодермальне походження). У хребетних кишкова трубка диференціюється на ротову порожнину, глотку, стравохід, шлунок, тонкий і товстий кишечники, які, проте, не у всіх організмів (класів) диференційовані повністю. Починаючи з круглоротих, у хребетних розвивається печінка, а починаючи з риб - підшлункова залоза. У риб розвиваються кісткові пластинки і зуби. У земноводних вперше з'являються слинні залози, закінчується відокремлення шлунка, тонкої і товстої кишок, клоаки, розвиваються однорядні зуби. Дуже інтенсивно розвивається мова. У рептилій намічається деяка диференціювання зубів, частина слинних залоз перетворюється на отруйні, поглиблюється відокремлення шлунка, в кишечнику розвивається зачаток сліпої кишки. У птахів здатність до польоту супроводжується змінами в травній системі. Замість щелеп і зубів у них з'являється дзьоб, а в стравоході утворюється еластичне розширення, зване зобом. Шлунок розділений на залозистий і м'язовий відділи, в яких відбувається хжліческая і механічна переробка їжі (відповідно). Збільшується в довжину товстий відділ кишечнику, але редукується мову. У ссавців розвиток травної системи досягає верхньої межі. Маючи значну довжину, вона характеризується більшою розчленованістю. Диференціюються зуби, розвивається стравохід. Шлунок, складається з декількох шарів і містить багато травних залоз. У деяких травоїдних (жуйних копитних) шлунок є багатокамерним. Кишечник диференційований на тонкий, товстий і прямий відділи. Значною диференціювання досягають печінка, підшлункова залоза, їх протоки впадають в передній відділ тонкого кишечника. Відбувається подовження товстого кишечника, з'являються сліпа кишка, і апендикс. У багатьох ссавців отримала значний розвиток сліпа кишка, досягаючи '/ g довжини всього кишечника. Товста кишка, закінчується анусом, відокремленим від сечостатевого отвори промежиною. У яйцекладущіх товста кишка закінчується клоакою. У людини до складу травної системи входить рот, глотка, стравохід, шлунок, кишечник, слинні залози, печінка, жовчний міхур і панкреатична заліза Всі відділи шлунково-кишкового тракту побудовані з чотирьох шарів. Людина є всеїдним істотою, оскільки його травна система забезпечує метаболізірованіе як рослинною, так і тваринної їжі (1).
Головними вуглеводами, що містяться в рослинній їжі, є крохмаль і целюлоза, а в тваринної їжі-глікоген.
Таблиця 1 Функції відділів травної системи людини
Відділи травної системи |
Ротова порожнина |
Стравохід |
Шлунок |
Тонкий кишечник |
Товстий кишечник |
Основні функції |
Надходження їжі, часткове перетравлення крохмалю |
Перехід їжі в шлунок |
Переварювання білків |
Перетравлювання і всмоктування |
Реадсорбції солей і води, формування калових мас |
Ферменти |
Амілаза |
|
Пепсин |
Трипсин, ліпаза |
|
Джерела |
Слинні залози |
|
Шлункові залози |
Панкреас, тонкий кишечник |
|
Утворені |
Складні цукру |
|
Крупномолекулярні фрагменти білків, деякіамінокислоти |
Жирні кислоти, глиця-рол, прості цукри, амінокислоти, фрагменти білків |
|
Перетравлювання вуглеводів починається
в порожнині рота, де слинна амілаза,
(птіалін) діє на крохмаль і глікоген,
перетворюючи їх у суміш Д-глюкози,
мальтози і олігосахаридів. Перетравлювання
вуглеводів завершується в тонкому
кишечнику, де переварений під дією
птіаліну крохмаль піддається подальшому
впливу панкреатичної амілази,
конвертується в мальтозу. У кінцевому
підсумку вся мальтоза під дією маль-тази
конвертується в глюкозу.
Всі
реакції відбуваються за участю води
(гідроліз) і ферментів:
Рот,
шлунок:
![]() Тонкий
кишечник:
Тонкий
кишечник:
![]()
![]()
![]()
![]() Неперетравлена
целюлоза сприяє нормальній
перистальтиці кишечника у
ссавців.
Переварювання жирів
починається в тонкому відділі кишечнику,
де панкреатична ліпаза за допомогою
жовчних кислот і білка коліпази
конвертує жири в
гліцерин і жирні кислоти. Одна молекула
жиру дає одну молекулу гліцеролу і три
молекули жирної кислоти:
Неперетравлена
целюлоза сприяє нормальній
перистальтиці кишечника у
ссавців.
Переварювання жирів
починається в тонкому відділі кишечнику,
де панкреатична ліпаза за допомогою
жовчних кислот і білка коліпази
конвертує жири в
гліцерин і жирні кислоти. Одна молекула
жиру дає одну молекулу гліцеролу і три
молекули жирної кислоти:
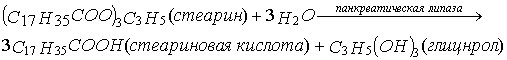 Переварюванню ліпідів в
тонкому відділі кишечнику
сприяють солі жовчних
кислот, які є похідними холевой кислоти.
Ці солі надходять з печінки в жовч, яка
потім виливається в передню частину
тонкого кишечника.
Білки починають
перетравлюватися (гідролізуються) у
шлунку під впливом пепсину і соляної
кислоти, виділення якої клітинами шлунка
відбувається під дією гормону - гастрит-ну.
Освічені продукти піддаються повторному
гідролізу трипсином і хемотрипсином,
попередники яких виробляються в клітинах
підшлункової залози, до поліпептидів,
після чого останні гідролізуються
пептідаза до амінокислот.
Однак
у шлунково-кишковому тракті людини не
всі білки перетравлюються
повністю. Наприклад, такий білок,
як кератин, перетравлюється лише
частково. Багато білків злакових також
перетравлюються не повністю, оскільки
вони покриті неперевариваемая целюлозою.
Переварюванню ліпідів в
тонкому відділі кишечнику
сприяють солі жовчних
кислот, які є похідними холевой кислоти.
Ці солі надходять з печінки в жовч, яка
потім виливається в передню частину
тонкого кишечника.
Білки починають
перетравлюватися (гідролізуються) у
шлунку під впливом пепсину і соляної
кислоти, виділення якої клітинами шлунка
відбувається під дією гормону - гастрит-ну.
Освічені продукти піддаються повторному
гідролізу трипсином і хемотрипсином,
попередники яких виробляються в клітинах
підшлункової залози, до поліпептидів,
після чого останні гідролізуються
пептідаза до амінокислот.
Однак
у шлунково-кишковому тракті людини не
всі білки перетравлюються
повністю. Наприклад, такий білок,
як кератин, перетравлюється лише
частково. Багато білків злакових також
перетравлюються не повністю, оскільки
вони покриті неперевариваемая целюлозою.