

1.1 ВИД – эволюционно сложившаяся совокупность особей, имеющая единый генотип, проявляющийся сходными фенотипическими признаками. ВАРИАНТ (ВАР) - особи одного вида, различающиеся по разным признакам (серовары, хемовары, культивары, морфовары, фаговары). ПОПУЛЯЦИЯ - совокупность особей одного вида, относительно длительно обитающих на определенной территории. КУЛЬТУРА - совокупность бактерий одного вида (чистая) или нескольких видов (смешанная), выращенная на питательной среде (жидкой или плотной). КОЛОНИЯ – видимое скопление бактерий одного вида на поверхности или в глубине плотной питательной среды. ШТАММ - чистая культура одного вида бактерий, выделенная в определенное время из одного источника. КЛОН - культура клеток, выращенная из одного микроорганизма методом клонирования. |
1. 2 R-колонии - характеризуется неровными краями и шероховатой поверхностью, второй тип - S-колоний- имеет круглую форму, гладкую поверхность. Процесс расщепления бактериальных клеток протекает в одном направлении: от S- к R-форме. Обратный переход R- в S-форму наблюдается реже. Для большинства вирулентных бактерий характерен рост в виде S-формы колоний. Биологическое значение S-R-диссоциации состоит в приобретении бактериями определенных селективных преимуществ, обеспечивающих их существование в организме человека или во внешней среде. К ним относится более высокая устойчивость S-форм к фагоцитозу макрофагами, бактерицидному действию сыворотки крови. R-формы обладают большей устойчивостью к факторам окружающей среды. Они более длительное время сохраняются в воде, молоке. Вместе с тем S-R-диссоциация во многих случаях усложняет бактериологическую диагностику ряда инфекционных заболеваний, например дизентерии Зонне, эшерихиоза.
|
2.2 1) механического разделения микроорганизмов: а) механическое разобщение бактериальных клеток на поверхности плотных питательных сред шпателем (по Дригальскому), рассев штрихами калиброванной петлей (по Гольду) или тампоном после забора им исследуемого материала; б) метод фильтрации, основан на пропускании исследуемого материала через специальные фильтры с определенным диаметром пор и разделение содержащихся микроорганизмов по величине. Этот метод применяется главным образом для очистки вирусов от бактерий, а также при получении фагов и токсинов (в фильтрате - чистый фаг, очищенный токсин). 2) Метод, основанный на получении серийных разведении в жидкой среде с последующим высевом на чашки с агаром (метод пластинчатых разводок по Коху). 3) Методы, основанные на биологических свойствах микроорганизмов: а) избирательном подавлении размножения сопутствующей микрофлоры при низкой температуре для получения культур психрофильных бактерий; б) метод Шукевича: клетки бактерий (протея), характеризующиеся скользящим типом движения, засеянные в конденсационную жидкость свежеприготовленного скошенного агара, поднимаются («всползают») по поверхности среды и разрастаются далеко за зону внесения посевного материала; в) метод прогревания: позволяет отделить спорообразующие бациллы от неспоровых форм. Прогревают исследуемый материал на водяной бане при 80°С 10-15 мин.); г) бактериостатический метод: основан на различном действии некоторых химических веществ и антибиотиков на микроорганизмы - определенные вещества угнетают рост одних микроорганизмов и не оказывают влияния на другие, д) метод заражения лабораторных животных или растений: основан на способности некоторых бактерий быстро размножаться в организме чувствительных к ним лабораторных животных (биопроба); При выделении и изучении облигатных паразитов этот метод является основным и единственным.
|
|
Метод Нейссера используется для выявления зерен волютина. Мазок окрашивают уксуснокислым метиленовым синим 2-3мин Промывают водой В течение 1 мин докрашивают везувином. Грам 1. Фиксированный мазок окрашивают карболовым раствором генцианового фиолетового в течение 1-2 минут. 2. В течение 1 минуты обрабатывают мазок раствором Люголя.3. Обесцвечивают спиртом 10-20 сек. 4. Промывают водой. 5. Докрашивают мазок водным раствором фуксина 1-2 минуты. Циль- окрашивают карболовым фуксином Циля (основной краситель) при нагревании 3-5 мин. 2. Обесцвечивают раствором серной кислоты (дифференцирующее вещество) в течение 1-2 мин. 3. Промывают водой. 4. Докрашивают 3-5 мин метиленовым синим (дополнительный краситель). Бури - Черную тушь смешивают с культурой и высушивают. 2. Проводят фиксацию в пламени горелки. 3. Окрашивают тела микробных клеток по Гинсу - водным фуксином в течение 1 минуты 4. Промывают водой |
|
3.3 а) получения генетического материала (искусственный синтез или выделения природных генов), б) включение этих генов в генетическую структуру, которая реплицируется автономно (векторную молекулу ДНК), то есть создание рекомбинантной молекулы ДНК, в) введение векторной молекулы (с включенным в нее геном) в клетку-реципиент, где она монтируется в хромосомный аппарат.
|
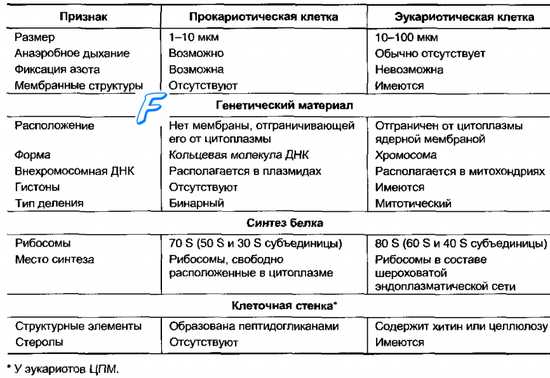
4.1 Клеточная стенка - обязательный структурный элемент. На долю клеточной стенки приходится от 5 до 50% сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. По строению и химическому составу клеточная стенка прокариот резко отличается от таковой клеточной стенки эукариотных организмов . В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Клетки большинства бактерий покрыты оболочкой - полимерной субстанцией, которая имеет множество свойств и функций. Эта оболочка, или капсула, отлична от двухслойной мембраны и располагается над ней. У бактерий термин "капсула" используется для определения высокомолекулярных полимеров, которые "прикрепляются" к поверхности бактерий. С наличием капсул связаны некоторые патогенные свойства бактерий. Капсулярные полисахариды бактерий могут необратимо связываться с отрицательно заряженными поверхностями. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В состав клеточной стенки эубактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных эубактерий он составляет основную массу вещества клеточной стенки (от 40 до 90%), у грамотрицательных - содержание пептидогликана значительно меньше (1-10%). Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2-3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8-10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны. Клеточная стенка грамположительных эубактерий плотно прилегает к ЦПМ в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм.
|
5.1 Внешний липопротеидный слой протопласта бактерий, обладающий особыми физическими и химическими свойствами, называется цитоплазматической мембраной. Цитоплазматическая мембрана выполняет очень важную роль — регулирует поступление веществ в клетку и выделение наружу продуктов обмена. Через мембрану питательные вещества могут поступать в клетку в результате активного биохимического процесса с участием ферментов. Кроме того, в мембране происходит синтез некоторых составных частей клетки, в основном компонентов клеточной стенки и капсулы. Наконец, в цитоплазматической мембране находятся важнейшие ферменты (биологические катализаторы). Упорядоченное расположение ферментов на мембранах позволяет регулировать их активность и предотвращать разрушение одних ферментов другими. С мембраной связаны рибосомы — структурные частицы, на которых синтезируется белок. Мембрана состоит из липопротеидов. Она достаточно прочна и может обеспечить временное существование клетки без оболочки. Цитоплазматическая мембрана составляет до 20% сухой массы клетки. Между плазматической мембраной и клеточной стенкой имеется связь в виде десмозов — мостиков. Цитоплазматическая мембрана часто дает инвагинации — впячивания внутрь клетки. Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. 5. 2 Репарация — клеточный механизм коррекции повреждённой последовательности ДНК. Одна из систем, восстанавливающая повреждения ДНК, вызванные УФ-лучами, названа системой фотореактивации. Ферменты, обеспечивающие фотореактивацию, действуют в присутствии видимого света и осуществляют расщепление тиминовых димеров, превращая их в мономерные формы. Активность другой системы, выполняющей эти же функции, обеспечивается ферментами действующими в отсутствии видимого света. Она названа системой темновой репарации, которую условно подразделяют на дорепликативную и пострепликативную. Состоит из 5 этапов, контролируемых множеством генов, кодирующих участвующие в репарации ферменты (эндонуклеазы, экзонуклеазы, ДНК-полимераза, ДНК-лигаза):
|
4.2 Трансформация - непосредственная передача генетического материала (фрагмента ДНК) донора реципиентной клетке. С донорной ДНК в реципиентную клетку обычно передается только один ген. Это связано с протяженностью трансформирующего фрагмента ДНК, который может проникнуть в реципиентную клетку. Обычно он не превышает 1/100 длины бактериальной хромосомы, т.е. включает один или несколько сцепленных генов. Эффективно трансформация происходит в опытах с бактериями одного и того же вида, имеющих разный генотип. Так, например, в опытах трансформации можно заместить гены «дикого» на мутировавшие или произвести замену обратного порядка. Трансформирующему воздействию ДНК поддаются не все, а только часть клеток бактериальной популяции. Клетки, способные воспринимать донорскую ДНК, называются компетентными. Состояние компетентности непродолжительно. Оно возникает в определенный период роста бактериальной культуры, чаще всего совпадающий с концом логарифмической фазы. В состоянии компетентности клеточная стенка бактерий становится проницаемой для высокополимерных фрагментов ДНК. По-видимому, это связано с тем, что трансформируемый фрагмент ДНК связывается с белком, образуя трансформасому, в которой он переносится в бактериальную клетку. Вместе с тем в трансформосоме он защищен от клеточных нуклеаз. Процесс трансформации бактерий можно подразделить на несколько фаз:
После проникновения внутрь клетки трансформирующая ДНК деспирализуется. Затем происходит физическое включение любой из двух нитей ДНК донора в геном реципиента.
Эффективность спаривания трансформирующей ДНК с соответствующим участком хромосомы реципиента зависит от степени гомологичности ДНК донора и реципиента. Чем выше гомологичность, тем эффективнее спаривание, что определяет конечный результат трансформации, т.е. количество формирующихся рекомбинантов (трансформантов). Отсюда ясно, почему межвидовая трансформация происходит гораздо реже, чем внутривидовая
|
6.1 Капсула. Клеточная стенка многих бактерий сверху окружена слоем слизистого материала — капсулой Толщина капсулы может во много раз превосходить диаметр самой клетки, а иногда она настолько тонкая, что ее можно увидеть лишь через электронный микроскоп, — микрокапсула. Капсула не является обязательной частью клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она служит защитным покровом клетки и участвует в водном обмене, предохраняя клетку от высыхания. По химическому составу капсулы чаще всего представляют собой полисахариды. Иногда они состоят изгликопротеидов (сложные комплексы сахаров и белков) и полипептидов (род Bacillus), в редких случаях — из клетчатки (род Acetobacter). Слизистые вещества, выделяемые в субстрат некоторыми бактериями, обусловливают, например, слизисто-тягучую консистенцию испорченного молока и пива.
|
6.2 Трансдукция – это передача генетической информации между бактериальными клетками с помощью умеренных трансдуцирующих фагов. Трансдуцирующие фаги могут переносить один ген или более. Этот вид генетического обмена открыт Н. Циндером и Дж. Ледербергом в 1951 г. Различают три типа трансдукции:
Неспецифическая трансдукция. В процессе репродукции фага в момент сборки фаговых частиц в их головку вместе с фаговой ДНК может проникнуть какой-либо фрагмент ДНК бактерии-донора. При этом фаг может утратить часть своего генома и стать дефектным. Такие дефектные трансдуцирующие фаги составляют примерно 0,3% всего потомства. При неспецифической трансдукции в клетки реципиентного штамма вместе с фаговой ДНК могут быть перенесены любые гены донора, например гены, контролирующие способность синтезировать аминокислоты, пурины, пиримидины, гены резистентности к антибиотикам. Принесенный фагом фрагмент ДНК бактерии-донора способен включаться в гомологическую область ДНК клетки-реципиента путем рекомбинации. Таким образом, при неспецифической трансдукции трансдуцирующие фаги являются только переносчиком генетического материала от одних бактерий к другим, поскольку сама фаговая ДНК не участвует в образовании рекомбинантов (трансдуктантов). Специфическая трансдукция характеризуется способностью фага переносить определенные гены от бактерии-донора к бактерии-реципиенту. Это связано с тем, что образование трансдуцирующего фага происходит путем выщепления профага из бактериальной хромосомы вместе с генами, расположенными на хромосоме клетки-донора рядом с профагом. Например, трансдуцирующий фаг лямбда (λ) переносит ген gal, контролирующий ферментацию галактозы, или ген bio, ответственный за синтез биотина. При выщеплении бактериальных генов вместе с ДНК профага из состава хромосомы часть фаговых генов утрачивается, в результате чего формируется дефектный фаг d, несущий ген gal клетки-донора. Он обозначается как dgal. В том случае, когда он несет ген bio, его обозначают как dbio. При взаимодействии трансдуцирующих фагов с клетками реципиентного штамма происходит включение гена бактерии-донора вместе с ДНК дефектного фага в хромосому бактерии-реципиента. Бактерии, лизогенированные дефектным фагом, невосприимчивы, как и все лизогенные клетки, к последующему заражению гомологичным вирулентным фагом. Абортивная трансдукция. При абортивной трансдукции принесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому бактерии-реципиента, а располагается в ее цитоплазме и может в таком виде функционировать. Во время деления бактериальной клетки трансдуцированный фрагмент ДНК-донора может передаваться только одной из двух дочерних клеток, т.е. наследоваться однолинейно и в конечном итоге утрачиваться в потомстве.
|
7.1 Их группируют вместе из-за сходства с микробами, поскольку они тоже размножаются делением, чувствительны к антибактериальным препаратам, однако не имеют некоторых структур (например, у микоплазм отсутствует клеточная стенка) либо некоторых особенностей метаболизма (например, у хламидии нет синтеза АТФ). Хламидии часто служат причиной мочеполовых и легочных инфекций, а также конъюнктивитов у новорожденных. Большинство риккетсии передается насекомыми, в частности вшами, вызывающими эпидемический (сыпной) тиф, а также клещами, индуцирующими лихорадку Скалистых гор, и водяными клещами, которые вызывают цуцугамуши (японскую речную лихорадку, акамуши). Риккетсии — облигатные внутриклеточные паразиты, размножающиеся в цитоплазме эндотелиальных клеток. Микоплазма и близкородственная ей уреаплазма являются после вирусов самыми мелкими из свободноживущих микроорганизмов. 8.1 По усвоению углерода бактерии можно разделить на два типа:
Аутотрофы способны получать углерод из неорганических соединений и даже из углекислоты. Энергию, необходимую для синтеза органических веществ, аутотрофы получают при окислении минеральных соединений. К аутотрофным бактериям относятся нитрифицирующие, железобактерии Гетеротрофы используют в качестве источника углерода органические соединения. Универсальным источником углерода служат различные углеводы (их часто добавляют в питательные среды), белки и др. Гетеротрофы играют значительную роль в уничтожении различных мертвых органических остатков. Такие бактерии называются сапрофитами (от sapros — гнилой, phyton — растение). Микробы, способные существовать за счет органических соединений организма животных и в клетках растений, получили название паразитических. Среди патогенных микроорганизмов выделяют так называемые облигатные паразиты, которые способны жить только в живых клетках или тканях. К таким микробам относятся риккетсии, вирусы и некоторые простейшие. По способности усваивать азот бактерии делятся также на две группы:
Аминоаутотрофы используют молекулярный азот воздуха. Бактерии этой группы — азотфиксирующие почвенные и клубеньковые бактерии — единственные живые существа, усваивающие свободный азот, принимают активное участие в круговороте азота в природе. Аминогетеротрофы получают азот из органических соединений — сложных белков. К аминогетеротрофам относятся все патогенные микроорганизмы и большинство сапрофитов. В настоящее время для аутотрофов применяется также название литотрофы, а для гетеротрофов — органотрофы.
По источникам энергии различают
В зависимости от окисляемого субстрата среди хемотрофных организмов выделяют хемолитотрофы и хемоорганотрофы .
|
Для культивирования бактерий используют питательные среды, к которым предъявляется ряд требований. 1. Питательность. Бактерии должны содержать все необходимые питательные вещества. 2. Изотоничность. Бактерии должны содержать набор солей для поддержания осмотического давления, определенную концентрацию хлорида натрия.3. Оптимальный рН (кислотность) среды. Кислотность среды обеспечивает функционирование ферментов бактерий; для большинства бактерий составляет 7,2–7,6.4. Оптимальный электронный потенциал, свидетельствующий о содержании в среде растворенного кислорода. Он должен быть высоким для аэробов и низким для анаэробов.5. Прозрачность (чтобы был виден рост бактерий, особенно для жидких сред). 6. Стерильность.Классификация питательных сред.1. По происхождению:1) естественные (молоко, желатин, картофель и др.);2) искусственные – среды, приготовленные из специально подготовленных природных компонентов (пептона, аминопептида, дрожжевого экстракта и т. п.);3) синтетические – среды известного состава, приготовленные из химически чистых неорганических и органических соединений.2. По составу:1) простые – мясопептонный агар, мясопептонный бульон;2) сложные – это простые с добавлением дополнительного питательного компонента (кровяного, шоколадного агара): сахарный бульон, желчный бульон, сывороточный агар, желточно-солевой агар, среда Китта—Тароцци.3. По консистенции:1) твердые (содержат 3–5 % агар-агара);2) полужидкие (0,15—0,7 % агар-агара);3) жидкие (не содержат агар-агара).4. По назначению:1) общего назначения – для культивирования большинства бактерий (мясопептонный агар, мясопептонный бульон, кровяной агар);2) специального назначения:а) элективные – среды, на которых растут бактерии только одного вида (рода), а род других подавляется (щелочной бульон, 1 %-ная пептонная вода, желточно-солевой агар, казеиново-угольный агар и др.);б) дифференциально-диагностические – среды, на которых рост одних видов бактерий отличается от роста других видов по тем или иным свойствам, чаще биохимическим (среда Эндо, Левина, Гиса, Плоскирева и др.);в) среды обогащения – среды, в которых происходит размножение и накопление бактерий-возбудителей какого-либо рода или вида (селенитовый бульон).
|
Ферменты Так, в цитоплазматической мембране находятся окислительно-восстановительные ферменты, которым принадлежит основная роль в дыхании клетки, ферменты, обеспечивающие доставку питательных веществ, и др. Ферменты, связанные с делением клетки, обнаруживаются в мезосомах, клеточной стенке, в месте образования перегородки.
У бактерий по характеру вызываемых ими превращений обнаруживаются следующие основные группы ферментов:
г и д р о л а з ы, вызывающие расщепление протеинов, углеводов, липидов путем присоединения молекул воды;
оксидоредуктазы, катализирующие окислительно-восстановительные реакции;
трансфера з ы, осуществляющие перенос отдельных атомов, от молекулы к молекуле;
л и а з ы, отщепляющие химические группы негидролитическим путем;
изомеразы, участвующие в углеводном обмене;
л и г а з ы, способствующие биосинтетическим реакциям клетки.
Ферменты бактерий классифицируются на экзоферменты и эндоферменты. Экзоферменты выделяются бактериальной клеткой в окружающую среду для внеклеточного переваривания. Этот процесс осуществляется с помощью гидролаз, которые расщепляют макромолекулы питательных веществ до простых соединений — глюкозы, аминокислот, жирных кислот. Такие соединения могут свободно проходить через оболочку клетки и с помощью пермеаз передаваться в цитоплазму клетки для участия в метаболизме, являясь источниками углерода и энергии. Некоторые экзоферменты выполняют защитную функцию, например, пенициллиназа, выделяемая многими бактериями, делает клетку недосягаемой для антибиотика — пенициллина.
Эндоферменты катализируют метаболические реакции, происходящие внутри клетки.
Ферменты бактерий классифицируются также на конститутивные и индуцибельные. Конститутивными называются такие ферменты, которые синтезируются клеткой независимо от наличия субстрата в среде, индуцибельные ферменты образуются бактериями только при наличии в среде соответствующего индуцирующего соединения, т. е. субстрата данного фермента. Например, в геноме кишечной палочки заложена способность разлагать лактозу, но только при наличии в среде лактозы клеткой синтезируется фермент, катализирующий ее гидролиз.
Известны также ферменты, которые получили название аллостерических . Кроме активного центра у них имеется регуляторный или аллостерический центр, который в молекуле фермента пространственно разделен с активным центром. Аллостерическим (от греч. allos - иной, чужой) он называется потому, что молекулы, связывающиеся с этим центром, по строению ( стерически ) не похожи на субстрат, но оказывают влияние на связывание и превращение субстрата в активном центре, изменяя его конфигурацию. Молекула фермента может иметь несколько аллостерических центров. Вещества, связывающиеся с аллостерическим центром, называют аллостерическими эффекторами. Они влияют через аллостерический центр на функцию активного центра: или облегчают ее, или затрудняют. Соответственно аллостерические эффекторы называются положительными (активаторы) или отрицательными (ингибиторы). Аллостерические ферменты играют важную роль в тонкой регуляции метаболизма бактерий. Поскольку практически все реакции в клетке катализируются ферментами, регуляция метаболизма сводится к регуляции интенсивности ферментативных реакций.
Патогенные бактерии обладают наряду с ферментами обмена также ферментами агрессии, являющимися факторами вирулентности. К таким ферментам относятся
гиалуронидаза,
дезоксирибонуклеаза,
коллагеназа,
н е й р а м и и и д аза, и др.
Гиалуронидаза стрептококков, например, расщепляет гиалуроновую кислоту в мембранах клеток соединительных тканей макроорганизма , что способствует распространению возбудителей и их токсинов в организме, обуславливая высокую инвазивность этих бактерий. Плазмокоагулаза является главным фактором патогенности стафилококков, так как участвует в превращении протромбина в тромбин, который вызывает образование фибриногена, в результате чего каждая бактерия покрывается пленкой, предохраняющей ее от фагоцитоза.Ферменты бактерий обладают высокой специфичностью, и именно это свойство широко используется при идентификации и определении видов микроорганизмов. Наибольшее значение имеет определение сахаролитических (ферментация сахаров) и протеолитических (разложение белков) свойств.