

Класифікація інтерферонів
Інтерферони (IFNs) ділять на 3 групи залежно від того, з яким рецептором вони зв’язуються:
IFNs І типу (зв’язуються з рецептором IFNs І типу): IFNα, β, δ, ε, κ, і ω.
IFNs ІІ типу (зв’язуються з рецептором IFNs ІІ типу): IFNγ.
IFNs ІІІ типу (використовується специфічний IFNλ рецептор): IFNλ.
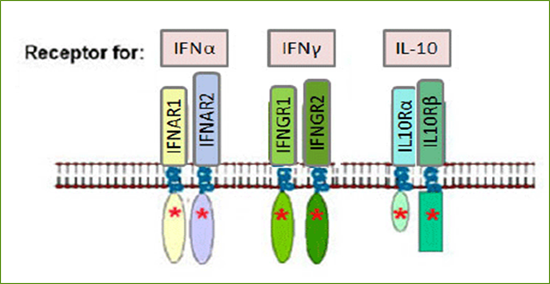
Рис.2. Структура рецепторів різних типів інтерферонів.
Типи інтерферонів відрізняються не лише за рецепторами, з якими вони взаємодіють, але й клітинами-продуцентами і генетичною мінливістю. У той час як IFN-γ кодується одним геном, IFN-λ має чотири підтипи,а IFN-α має до 14. Інтерферони I типу є найбільш поширеними і можуть продукуватися практично будь-якою клітиною в організмі, хоча IFN-α синтезується в основному макрофагами і дендритними клітинами, а IFN-β продукується переважно фібробластами. Тип II і тип III інтерферонів є набагато більш обмеженими у типах клітин, які можуть їх виробляти і реагувати на них.
4. Рецептори інтерферонів і шляхи передачі сигналів
4. 1. IFN-α/β рецептор
IFN-α/β рецептор локалізований на багатьох типах клітин. Рецептор є гетеродимером (Рис.3), що складається з двох субодиниць: IFNAR1 (550 а.з. інтегрального мембранного білка) і IFNAR2 (487 а.з. інтегрального мембранного білка). Зовнішньоклітинні домени субодиниць рецептора утворені кількома фібронектин ІІІ-подібними повторами (IFNAR1 має 4 повтори, IFNAR2 має 2 повтори). У присутності IFN-β дві субодиниці з’єднюються, утворюючи функціональний рецепторний комплекс.

Рис.3. Будова IFN-α/β рецептора.
Внутрішньоклітинні домени IFNAR1 і IFNAR2 асоційовані з двома тирозиновими кіназами родини Януса - Tyk2 і JAK1 (Рис.4), які після утворення функціонального рецептора трансфосфорилюють одна одну і фосфорилюють рецептор. Далі кінази фосфорилюють молекули білків STAT1 і STAT2 ((signal transducer and activator of transcription 1 and 2), після чого до утвореного гетеродимеру приєднюється інтерферон регулюючий фактор 9 (IRF-9). Утворений комплекс має назву ISGF3-комплекс. Він проникає у ядро і стимулює транскрипцію ISREs (IFN-stimulated response elements).

Рис.4. Шлях передачі сигналу через IFN-α/β рецептор.
2. IFN-γ рецептор
IFN-γ рецептор експресується майже на всіх ядровмісних клітинах. Функціональний IFN-γ рецептор складається з двох субодиниць: IFNgR1 (90 кДа кожна) і двох субодиниць IFNgR2 (62 кДа кожна) (Рис.5). Людський ген IFNGR1 містить 7 екзонів і знаходиться у 6 хромосомі. Позаклітинна частина IFNgR1 містить IFN-γ –зв’язуючий домен; внутрішньоклітинна частина необхідна для трансдукції сигналу і рециклювання рецептора. Ген IFNGR2 теж має 7 екзонів і знаходиться в 21 хромосомі. Внутрішньоклітинна частина IFNgR2 необхідна для передачі сигналу. Зовнішньоклітинна частина взаємодіє з IFNgR1/IFN-γ комплексом і не відіграє важливої ролі у зв’язуванні ліганду.

Рис.5.Будова IFN-γ рецептора.
Шлях передачі сигналу (Рис.6):

Рис.6. Шлях передачі сигналу через IFN-γ рецептор.
Збирання рецептора та активація тирозинових кіназ JAK1і JAK2.
Фосфорилювання внутрішньоклітинної частини IFNgR1.
Фосфорилювання STAT1 й утворення гомодимеру.
Транслокація утвореного гомодимеру в ядро та активація транскрипції IFN-γ-індуцибельних генів.
3. IFN-λ рецептор
IFN-λ рецептор складається з 2 субодиниць (Рис.7): IFN-λR1 і IL-10R2 . IFN-λR1 використовується лише IFN ІІІ типу, тоді як IL-10R2 використовується також IL-10, IL-22 і IL-26.
Обидві субодиниці мають позаклітинну частину, утворену 2 фібронектин-ІІІ-подібними повторами. Внутрішньоклітинна частина IFN-λR1 містить 270 а. з.,тоді як субодиниця IL-10R2 – 82 а.з. Невідомо, чи рецептор знаходиться
на поверхні клітини у «готовому» вигляді чи стає активним після зв’язування ліганду.

Рис.7. Будова IFN-λ рецептора.
Шлях передачі сигналу через IFN-λ рецептор аналогічний шляху передачі сигналу через IFN-α/β рецептор (Рис.8).
Головна відмінність між інтерферонами І і ІІІ типу є поширення рецепторів. IFN-α/β рецептор є «повсюдно» експресований, тоді як IFN-λ рецептор присутній на окремих типах клітин (частково клітини епітеліального походження, кератиноцити).

Рис.8. Шлях передачі сигналу через IFN-λ рецептор.