

Контакты сцепляющего типа:
десмосомы
адгезивный поясок
Контакты запирающего типа:
плотное соединение (запирающая зона)
Контакты коммуникационного типа:
щелевидные соединения (нексусы)
синапсы
Простое межклеточное соединение осуществляется путем сближения плазмолемм клеток до расстояния 15-20нм и взаимодействия белков плазматических мембран – кадгеринов. Имеются разнообразные семейства кадгеринов, характерные для той или иной ткани. Благодаря кадгерину клетки в процессе гистогенеза и органогенеза узнают друг друга и объединяются в единую структуру, например, эпителиальный пласт. (Раковые клетки не узнают друг друга). Пальцевидные соединения (интердигитации) образуются за счет взаимной инвагинации (впячивания) обеих плазмолемм в начале в одном, а затем в другом. Это один из трех видов контактов между кардиомиоцитами. Десмосома представляет небольшое округлое образование, построенное с участием плазмолемм соседних клеток. Десмосомы построены из белка десмоплакина, который образует слой на внутренней стороне каждой мембраны. К слою десмоплакина присоединяются пучки промежуточных филаментов. Промежуточные филаменты в разных тканях представлены разными белками, например, в эпителии – кератином, в мышечной – десмином. С наружной стороны мембраны пространство между десмосомами заполнено утолщенным слоем гликокаликса. Гликокаликс десмосом пронизан склеивающим (адгезивным) белком – десмоглеином. Адгезивный поясок встречается в однослойных эпителиях, имеет вид двойных лент. По структуре адгезивный поясок похож на десмосому, но образован другими белками. Плотное соединение образуется с помощью интегральных адгезивных белков. В таких контактах плазмолеммы плотно прилегают друг к другу. Плотные соединения также имеют лентовидную форму. Однако ленты имеют вид ячеистой сети. Плотные контакты надежно разграничивают компартменты, находящиеся с базальной и апикальной (верхушечной) сторон однослойного эпителия. Контакты в виде плотных соединений имеются вэндотелии сосудов. 9) Включения — это непостоянные компоненты цитоплазмы, содержание которых меняется в зависимости от функционального состояния клетки. Различают трофические, секреторные и экскреторные включения. Трофические включения (trophe — пища) — наиболее значительная группа, это запасные питательные вещества, за счет которых клетки живут и строят свое тело. Однако они имеют значение не только для данной клетки, но и для других клеток и тканей организма. Среди трофических включений особенно важную роль играют жиры и углеводы. В виде жира запасный питательный материал откладывается в основном в клетках жировой ткани. Внешний вид этих клеток сильно изменяется в зависимости от степени отложения жира. Последний в цитоплазме появляется в виде мелких капелек, затем они увеличиваются и сливаются в одну огромную каплю, занимающую центральное положение в клетке, цитоплазма и ядро при этом оттесняются к периферии. Трофические включения в виде жира характерны также для эпителиальных клеток кишечника и др. При голодании животного запасной жир расходуется прежде всего. Включения типа углеводов очень распространены в клетках растений (крахмал и сахар). В организме животных углеводы отлагаются в форме гликогена (животный крахмал). Особенно богаты им клетки печени и мышечная ткань. Белки в виде запасных питательных веществ у животных встречаются очень редко. Их находят в клетках печени в виде разнообразной формы и величины зернышек и в женских половых клетках в виде желточных зерен. Некоторые витамины находятся в клетке в виде микроскопически видимых включений. Секреторные включения являются продуктами жизнедеятельности клеток желез внешней и внутренней секреции. К ним относятся ферменты, гормоны, слизь и другие вещества, подлежащие выведению из клетки. Экскреторные включения представляют собой продукты обмена веществ в растительных и животных клетках (кристаллы щавелевой кислоты, щавелевокислого кальция и др.). Пигментные включения придают клеткам и тканям определенную окраску. В клетках у млекопитающих особенно распространены пигменты —меланины, обусловливающие черную и коричневую окраску, липохромы, имеющие желтую или красную окраску, и др. Клетки с боль-шим количеством этих включений называют пигментными. Они разбросаны поодиночке в отдельных местах организма или, скопляясь в больших количествах, образуют пигментную ткань. Иногда пигментные включения появляются в нервных клетках. Секреты и инкреты — включения многих железистых клеток. Секреты —это слюна, желчь и т. д. К инкретам относят физиологически активные вещества —гормоны, выделяемые органами внутренней секреции: щитовидной и зобной железами, надпочечниками, гипофизом и др. Одни гормоны влияют на рост организма, другие — на обмен веществ, третьи — на работу половых желез и т. д. Экскреты — конечные продукты жизнедеятельности клеток. Они обычно выводятся из клетки, но могут иногда задерживаться в ней довольно длительное время. 10) Эндоцито́з (англ. endocytosis) — процесс захвата (интернализации) внешнего материала клеткой, осуществляемый путём образования мембранных везикул. В результате эндоцитоза клетка получает для своей жизнедеятельности гидрофильный материал, который иначе не проникает через липидный бислой клеточной мембраны. Различают фагоцитоз, пиноцитоз и рецептор-опосредованный эндоцитоз. Термин был предложен в 1963 году бельгийским цитологом Кристианом де Дювом для описания множества процессов интернализации, развившихся в клетке млекопитающих.[1] ТИПЫ Фагоцитоз (поедание клеткой) — процесс поглощения клеткой твёрдых объектов, таких как клетки эукариот, бактерии, вирусы, остатки мёртвых клеток и т. п. Вокруг поглощаемого объекта образуется большая внутриклеточная вакуоль (фагосома). Размер фагосом — от 250 нм и больше. Путем слияния фагосомы с первичной лизосомой образуется вторичная лизосома. В кислой среде гидролитическиеферменты расщепляют макромолекулы, оказавшиеся во вторичной лизосоме. Продукты расщепления (аминокислоты, моносахариды и прочие полезные вещества) транспортируются затем через лизосомную мембрану в цитоплазму клетки. Фагоцитоз распространен очень широко. У высокоорганизованных животных и человека процесс фагоцитоза играет защитную роль. Фагоцитарная деятельность лейкоцитови макрофагов имеет огромное значение в защите организма от попадающих в него патогенных микробов и других нежелательных частиц. Фагоцитоз впервые описал русский ученый И. И. Мечников.
Пиноцитоз (питьё клеткой) — процесс поглощения клеткой жидкой фазы из окружающей среды, содержащей растворимые вещества, включая крупные молекулы (белки, полисахариды и др.). При пиноцитозе от мембраны отшнуровываются внутрь клетки небольшие пузырьки — эндосомы. Они меньше фагосом (их размер до 150 нм) и обычно не содержат крупных частиц. После образования эндосомы к ней подходит первичная лизосома, и эти два мембранных пузырька сливаются. Образовавшаяся органелла носит название вторичной лизосомы. Процесс пиноцитоза постоянно осуществляют все эукариотическме клетки.
Рецептор-опосредованный эндоцитоз — активный специфический процесс, при котором клеточная мембрана выпучивается внутрь клетки, формируя окаймлённые ямки. Внутриклеточная сторона окаймлённой ямки содержит набор адаптивных белков (адаптин, клатрин, обуславливающий необходимую кривизну выпучивания, и др. белки). Макромолекулы, связывающиеся со специфическими рецепторами на поверхности клетки, проходят внутрь со значительно большей скоростью, чем вещества, поступающие в клетки за счет пиноцитоза. Внешняя сторона мембраны при этом включает специфические рецепторы (например, ЛПНП-рецептор). При связывании лиганда из окружающей клетку среды окаймлённые ямки формируют внутриклеточные везикулы (окаймлённые пузырьки). Рецептор-опосредованный эндоцитоз включается для быстрого и контролируемого поглощения клеткой соответствующего лиганда (например, ЛПНП). Эти пузырьки быстро теряют свою кайму и сливаются между собой, образуя более крупные пузырьки — эндосомы. После чего эндосомы сливаются с первичными лизосомами, в результате чего формируются вторичные лизосомы. Например, когда животной клетке необходим холестерин для синтеза мембраны, она экспрессирует ЛПНП-рецепторы на плазматической мембране. Богатые холестерином и эфирами холестерина ЛПНП, связавшиеся с ЛПНП-рецепторами, быстро доставляют холестерин в клетку.
Распространенность
Типичный эндоцитоз встречается у эукариот, лишенных клеточной стенки — животных и многих протистов. Долгое время считалось. что прокариоты полностью лишены способности к эндоцитозу. Однако в 2010 году эндоцитоз был описан у бактерий рода Gemmata[2]
Экзоцитоз (от греч. Έξω — внешний и κύτος — клетка) — у эукариот клеточный процесс, при котором внутриклеточные везикулы (мембранные пузырьки) сливаются с внешней клеточной мембраной. При экзоцитозе содержимое секреторных везикул (экзоцитозных пузырьков) выделяется наружу, а их мембрана сливается с клеточной мембраной. Практически все макромолекулярные соединения (белки, пептидные гормоны и др.) выделяются из клетки этим способом.
У прокариот везикулярный механизм экзоцитоза не встречается, у них экзоцитозом называют встраивание белков в клеточную мембрану (или в наружную мембрану у грамотрицательных бактерий), выделение белков из клетки во внешнюю среду или в периплазматическое пространство.
Экзоцитоз может выполнять три основные задачи:
доставка на клеточную мембрану липидов, необходимого для роста клетки;
высвобождение различных соединений из клетки, например, токсичных продуктов метаболизма или сигнальных молекул (гормонов или нейромедиаторов);
доставка на клеточную мембрану функциональных мембранных белков, таких как рецепторы или белки-транспортёры. При этом часть белка, которая была направлена внутрь секреторной везикулы, оказывается выступающей на наружной поверхности клетки.
Этапы
Различают следующие этапы экзоцитоза:
Транспортировка везикулы от места синтеза и формирования (аппарат Гольджи) до места доставки осуществляется моторными белками вдоль актиновыхфиламентов либо микротрубочек цитоскелета. Этот этап может потребовать перемещения секретируемого материала на значительное расстояние, как, например, внейроне. Когда везикула достигает места секреции, она входит в контакт со специфическими удерживающими факторами клеточной мембраны.
Удержание доставленной везикулы обеспечивается относительно слабыми связями на расстоянии более 25 нм и может служить, например, для концентрирования синаптических везикул около пресинаптической мембраны.
Стыковка везикулы с мембраной является непосредственным продолжением первой фазы доставки, когда мембрана везикулы входит в близкий контакт с мембраной клетки (5-10 нм). Это включает прочное соединение белковых компонентов обеих мембран, вызванным внутримолекулярными перестановками, и предваряет формирования SNARE комплекса.
Стимуляция (прайминг) везикулы фактически соответствует образованию особого SNARE комплекса между двумя мембранами и осуществляется только в случае нейронального экзоцитоза. Этот этап включает процессы молекулярных перестановок и АТФ-зависимые модификации белков и липидов, происходящие непосредственно до слияния мембран в ответ на подъём уровня свободного кальция. Этот кальций-зависимый процесс необходим для контролируемого быстрого выброса нейромедиатора и отсутствует в случае конститутивного экзоцитоза.
Слияние мембраны везикулы с мембраной клетки приводит к высвобождению, или выбросу, содержания секретируемой везикулы во внеклеточное пространство и объединению липидного бислоя везикулы с внешней мембраной. В случае синаптического выброса процесс слияния, так же как и стимуляция, осуществляется SNARE комплексом. 11)
Клеточный цикл — это период существования клетки от момента её образования путем деления материнской клетки до собственного деления или гибели Длительность клеточного цикла эукариот
Длительность клеточного цикла у разных клеток варьируется. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные или базальные клетки эпидермиса и тонкой кишки, могут входить в клеточный цикл каждые 12—36 ч. Короткие клеточные циклы (около 30 мин) наблюдаются при быстром дроблении яициглокожих, земноводных и других животных. В экспериментальных условиях короткий клеточный цикл (около 20 ч) имеют многие линии клеточных культур. У большинства активно делящихся клеток длительность периода между митозами составляет примерно 10—24 ч.
Фазы клеточного цикла эукариот
Клеточный цикл эукариот состоит из двух периодов:
Период клеточного роста, называемый «интерфаза», во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
Периода клеточного деления, называемый «фаза М» (от слова mitosis — митоз).
Интерфаза состоит из нескольких периодов:
G1-фазы (от англ. gap — промежуток), или фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов;
S-фазы (от англ. synthesis — синтез), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они, конечно, есть).
G2-фазы, во время которой идет подготовка к митозу.
У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G1 фаза. Такие клетки находятся в фазе покоя G0.
Период клеточного деления (фаза М) включает две стадии:
кариокинез (деление клеточного ядра);
цитокинез (деление цитоплазмы).
В свою очередь, митоз делится на пять стадий.
Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопиификсированных и окрашенных клеток.
Регуляция клеточного цикла
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, как циклин-зависимые киназы и циклины.Клетки, находящиеся в G0 фазе, могут вступать в клеточный цикл при действии на них факторов роста. Разные факторы роста, такие как тромбоцитарный, эпидермальный, фактор роста нервов, связываясь со своими рецепторами, запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции геновциклинов и циклин-зависимых киназ. Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами. Содержание различныхциклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса. Киназы не активны без циклинов. На разных стадиях клеточного цикла синтезируются разные циклины. Так, содержание циклина B в ооцитах лягушки достигает максимума к моменту митоза, когда запускается весь каскад реакций фосфорилирования, катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.
Контрольные точки клеточного цикла
Для определения завершения каждой фазы клеточного цикла необходимо наличие в нем контрольных точек. Если клетка «проходит» контрольную точку, то она продолжается «двигаться» по клеточному циклу. Если же какие-либо обстоятельства, например повреждение ДНК, мешают клетке пройти через контрольную точку, которую можно сравнить со своего рода контрольным пунктом, то клетка останавливается и другой фазы клеточного цикла не наступает по крайней мере до тех пор, пока не будут устранены препятствия, не позволявшие клетке пройти через контрольный пункт. Существует как минимум четыре контрольных точки клеточного цикла: точка в G1, где проверяется интактность ДНК, перед вхождением в S-фазу, сверочная точка в S-фазе, в которой проверяется правильность репликации ДНК, сверочная точка в G2, в которой проверяются повреждения, пропущенные при прохождении предыдущих сверочных точек, либо полученные на последующих стадиях клеточного цикла. В G2 фазе детектируется полнота репликации ДНК, и клетки, в которых ДНК недореплицирована, не входят в митоз. В контрольной точке сборки веретена деления проверяется, все ли кинетохоры прикреплены к микротрубочкам
12) Клеточная теория — одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений, животных и остальных живых организмов с клеточным строением, в котором клетка рассматривается в качестве единого структурного элемента живых организмов. Общие сведения
Клеточная теория — основополагающая для биологии теория, сформулированная в середине XIX века, предоставившая базу для понимания закономерностей живого мира и для развития эволюционного учения. Маттиас Шлейден и Теодор Шванн сформулировали клеточную теорию, основываясь на множестве исследований о клетке(1838). Рудольф Вирхов позднее (1858) дополнил её важнейшим положением (всякая клетка происходит от другой клетки).
Шлейден и Шванн, обобщив имеющиеся знания о клетке, доказали, что клетка является основной единицей любого организма. Клетки животных, растений и бактерииимеют схожее строение. Позднее эти заключения стали основой для доказательства единства организмов. Т. Шванн и М. Шлейден ввели в науку основополагающее представление о клетке: вне клеток нет жизни. Клеточная теория дополнялась и редактировалась с каждым разом.
Положения клеточной теории Шлейдена-Шванна
Все животные и растения состоят из клеток.
Растут и развиваются растения и животные путём возникновения новых клеток.
Клетка является самой маленькой единицей живого, а целый организм — это совокупность клеток.
Основные положения современной клеточной теории
Клетка - это элементарная, функциональная единица строения всего живого. (Кроме вирусов, которые не имеют клеточного строения)
Клетка - единая система, она включает множество закономерно связанных между собой элементов, представляющих целостное образование, состоящее из сопряжённых функциональных единиц - органоидов.
Клетки всех организмов гомологичны.
Клетка происходит только путём деления материнской клетки.
Многоклеточный организм представляет собой сложную систему из множества клеток, объединённых и интегрированных в системы тканей и органов, связанных друг с другом.
Клетки многоклеточных организмов тотипотентны.
Дополнительные положения клеточной теории
Для приведения клеточной теории в более полное соответствие с данными современной клеточной биологии список её положений часто дополняют и расширяют. Во многих источниках эти дополнительные положения различаются, их набор достаточно произволен.
Клетки прокариот и эукариот являются системами разного уровня сложности и не полностью гомологичны друг другу (см. ниже).
В основе деления клетки и размножения организмов лежит копирование наследственной информации — молекул нуклеиновых кислот («каждая молекула из молекулы»). Положения о генетической непрерывности относится не только к клетке в целом, но и к некоторым из её более мелких компонентов — к митохондриям,хлоропластам, генам и хромосомам.
Многоклеточный организм представляет собой новую систему, сложный ансамбль из множества клеток, объединённых и интегрированных в системе тканей и органов, связанных друг с другом с помощью химических факторов, гуморальных и нервных (молекулярная регуляция).
Клетки многоклеточных тотипотентны, то есть обладают генетическими потенциями всех клеток данного организма, равнозначны по генетической информации, но отличаются друг от друга разной экспрессией (работой) различных генов, что приводит к их морфологическому и функциональному разнообразию — к дифференцировке. 13)
Амито́з, или прямо́е деле́ние кле́тки (от др.-греч. ἀ- — частица отрицания и μίτος — «нить») — деление клеток простым разделением ядра надвое.
Впервые он описан немецким биологом Робертом Ремаком в 1841 году, термин предложен гистологом Вальтером Флеммингом в 1882 году. Амитоз — редкое, но иногда необходимое явление[1]. В большинстве случаев амитоз наблюдается в клетках со сниженной митотической активностью: это стареющие или патологически измененные клетки, часто обреченные на гибель (клетки зародышевых оболочек млекопитающих, опухолевые клетки и др.).
При амитозе морфологически сохраняется интерфазное состояние ядра, хорошо видны ядрышко и ядерная оболочка. Репликация ДНК отсутствует. Спирализацияхроматина не происходит, хромосомы не выявляются. Клетка сохраняет свойственную ей функциональную активность, которая почти полностью исчезает при митозе. При амитозе делится только ядро, причем без образования веретена деления, поэтому наследственный материал распределяется случайным образом. Отсутствие цитокинезаприводит к образованию двуядерных клеток, которые в дальнейшем не способны вступать в нормальный митотический цикл. При повторных амитозах могут образовываться многоядерные клетки.
Это понятие ещё фигурировало в некоторых учебниках до 1980-х гг. В настоящее время считается, что все явления, относимые к амитозу — результат неверной интерпретации недостаточно качественно приготовленных микроскопических препаратов, или интерпретации как деления клетки явлений, сопровождающих разрушение клеток или иные патологические процессы. В то же время некоторые варианты деления ядер эукариот нельзя назвать митозом или мейозом. Таково, например, делениемакронуклеусов многих инфузорий, где без образования веретена происходит сегрегация коротких фрагментов хром
14. Мито́з (др.-греч. μίτος — нить) — непрямое деление клетки, наиболее распространенный способ репродукцииэукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределениихромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений.
Митоз — один из фундаментальных процессов онтогенеза. Митотическое деление обеспечивает ростмногоклеточных эукариот за счёт увеличения популяций клеток тканей. В результате митотического деления клеток меристем увеличивается количество клеток тканей растений. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений.
На основании морфологических особенностей митоз условно подразделяется на стадии: профазу, прометафазу, метафазу, анафазу, телофазу. Первые описания фаз митоза и установление их последовательности были предприняты в 70—80-х годах XIX века. В конце 1870-х — начале 1880-х годов немецкий гистолог Вальтер Флемминг для обозначения процесса непрямого деления клетки ввёл термин «митоз».
Продолжительность митоза в среднем составляет 1—2 часа[. Митоз клеток животных, как правило, длится 30—60 минут, а растений — 2—3 часа.[5]. За 70 лет в теле человека суммарно осуществляется порядка 1014клеточных делений[6].
Типы митоза
Выработка единой типологии и классификации митозов осложняется целым спектром признаков,[~ 1] которые в различных комбинациях создают разнообразие и неоднородность картин митотического деления. При этом отдельные варианты классификации, разработанные применительно к одним таксонам, являются неприемлемыми в отношении других, поскольку не учитывают специфики их митозов. Например, отдельные варианты классификации митозов, свойственных животным или растительным организмам, оказываются неприемлемыми для водорослей.
Одним из ключевых признаков, лежащих в основе различных типологий и классификаций митотического деления, является поведение ядерной оболочки. Если образование веретена и само митотическое деление протекает внутри ядра без разрушения ядерной оболочки, то такой тип митоза называют закрытым. Митоз с распадом ядерной оболочки, соответственно, называется открытым, а митоз с распадом оболочки только на полюсах веретена, с образованием «полярных окон» —полузакрытым.
Ещё одним характерным признаком является тип симметрии митотического веретена. При плевромитозе веретено деления билатерально симметрично либо асимметрично и состоит, как правило, из двух полуверетён, располагающихся в метафазе-анафазе под углом друг к другу. Для категории ортомитозов характерна биполярная симметрия веретена деления, а в метафазе зачастую наблюдается различимая экваториальная пластинка.
В рамках обозначенных признаков наиболее многочисленным является типичный открытый ортомитоз, на примере которого ниже рассматриваются принципы и стадии митотического деления. Данный тип митоза характерен для животных, высших растений и некоторых простейших.
Варианты классификации митозов
7 типов митоза простейших[18]:
|
6 типов митоза водорослей[17]:
|
Основные типы митоза у водорослей: 1. Закрытый центрический; 2. Закрытый ацентрический; 3. Полузакрытый центрический; 4. Полузакрытый ацентрический; 5. Открытый центрический; 6. Открытый ацентрический |
Происхождение и эволюция митоза
Возможные пути эволюции типов митоза у простейших. В квадратных скобках изображена гипотетическая промежуточная стадия. Условные обозначения: 1. Закрытый эвгленоидный митоз; 2. Закрытый внутриядерный плевромитоз; 3. Закрытый внеядерный плевромитоз; 4. Полузакрытый плевромитоз; 5. Закрытый внутриядерный ортомитоз; 6. Полузакрытый ортомитоз; 7. Открытый ортомитоз;
Предполагается, что сложный митотический процесс высших организмов развивался постепенно из механизмов деления прокариот.[20] Это предположение подтверждается тем, что прокариоты появились приблизительно на миллиард лет раньше первых эукариот. Кроме того, в митозе эукариот и бинарном делении прокариот принимают участие схожие белки.
Возможные промежуточные стадии между бинарным делением и митозом можно проследить у одноклеточных эукариот, у которых в ходе деления не разрушается ядерная оболочка. У большинства же других эукариот, в том числе растений и животных, веретено деления формируется вне ядра, а ядерная оболочка разрушается в течение митоза. Хотя митоз у одноклеточных эукариот ещё недостаточно изучен, можно предположить, что он произошёл от бинарного деления и в конечном счёте достиг того уровня сложности, который имеется у многоклеточных организмов[21].
У многих простейших эукариот митоз также остался процессом, связанным с мембраной, однако теперь уже неплазматической, а ядерной.[22] Возможно, в связи с увеличением размера и числа хромосом, структура типамезосомы разделилась на два элемента: ЦОМТ на ядерной оболочке и кинетохор на хромосоме. Для соединения данных структур между собой в процессе эволюции развилась промежуточная система микротрубочек. В рамках данного представления, наиболее древним и примитивным считается закрытый внутриядерный плевромитоз. Сегрегация хромосом при этом происходит путём расхождения ЦОМТ, к которым хромосомы крепятся посредством микротрубочек. В свою очередь ЦОМТ закреплены на ядерной оболочке и расходятся за счёт роста ядерной мембраны между ними.[23]
От разных вариантов закрытого внутриядерного плевромитоза, вероятно, берут своё начало несколько параллельных эволюционных линий.[23] В качестве эволюционно прогрессивных признаков при этом рассматриваются: распад ядерной оболочки во время митоза; переход ЦОМТ из ядра в цитоплазму; образование биполярного веретена; усиление спирализации хромосом; формирование экваториальной пластинки в метафазе. Таким образом, эволюция митотического деления идёт в направлении от закрытого внутриядерного плевромитоза к открытому ортомитозу.[24]
Регуляция митоза
Основными регулято́рными механизмами митоза являются процессы фосфорилирования и протеолиза[25]. Обратимые реакции фосфорилирования и дефосфорилирования обеспечивают протекание обратимых событий митоза, таких как сборка/распад веретена деления или распад/восстановление ядерной оболочки. Протеолиз лежит в основе необратимых событий митоза, таких как разделение сестринских хроматид в анаф
15)Механизм движения клетки
Вращение мотора вызывает пассивное вращение филамента. Более массивная клетка начинает вращаться примерно со скоростью 1/3 от скорости жгутика и в обратном направлении, а также приобретает поступательное движение.
Подавляющее большинство наделённых жгутиком бактерий имеют палочковидную форму. Из гидродинамических расчётов следует, что для наиболее эффективного движения отношение длины клетки к ширине должно составлять 3,7. Движение кокков крайне неэффективно, поэтому они чаще всего неподвижны.
У ряда бактерий мотор и жгутик могут вращаться только в одном направлении, переориентация происходит при остановке за счёт броуновского движения. Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один пучок. Для совершения кувырка они либо меняют направление вращения, либо изменяют его скорость, из-за чего пучок распадается. При полярном расположении жгутиков один из них может служить и толкающим, и тянущим приспособлением в зависимости от направления вращения.
Скорости движения бактерий варьируют от 20 мкм/с у некоторых Bacillus до 200 мкм/с у Vibrio.
16.
Сперматозо́ид (от др.-греч. σπέρμα (род. п. σπέρματος) — семя, ζωή — «жизнь» и εἴδος — «вид») — мужская половая клетка, мужская гамета, которая служит для оплодотворения женской гаметы, яйцеклетки. Термин используется для обозначения мелких, обычно подвижных гамет у организмов, которым свойственна оогамия. Обычно они значительно меньше яйцеклетки, поскольку не содержат столь значительного количества цитоплазмы и производятся организмом одновременно в значительном количестве. Понятие «сперматозоид» необходимо отличать от понятия «сперма», поскольку последняя состоит из семенной жидкости (в которой содержатся сперматозоиды), а также содержит небольшое количество эпителиальных клетокмочеиспускательного канала. Синонимы: живчик, иногда спермий. Обычно (особенно в ботанике) спермиями называют сперматозоиды, лишенные жгутиков. Строение и функция
Строение сперматозоида
Сперматозоид человека — это специализированная клетка, строение которой позволяет ей выполнить свою функцию: преодолеть половые пути женщины и проникнуть в яйцеклетку, чтобы внести в неё генетический материал мужчины. Сперматозоид, сливаясь с яйцеклеткой, оплодотворяет её.
В организме человека сперматозоид является самой маленькой клеткой тела (если учитывать только саму головку без хвостика). Общая длина сперматозоида у человека равна приблизительно 55 мкм. Головка составляет приблизительно 5,0 мкм в длину, 3,5 мкм в ширину и 2,5 мкм в высоту, средний участок и хвостик — соответственно, приблизительно 4,5 и 45 мкм в длину.[1]
Малые размеры, вероятно, необходимы для быстрого движения сперматозоида. Для уменьшения размера сперматозоида при его созревании происходят специальные преобразования: ядро уплотняется за счет уникального механизма конденсации хроматина (из ядра удаляются гистоны, и ДНК связывается с белками-протаминами), большая часть цитоплазмы выбрасывается из сперматозоида в виде так называемой «цитоплазматической капли», остаются только самые необходимые органеллы.
Сперматозоид мужчины имеет типичное строение и состоит из головки, средней части и хвоста.
Головка сперматозоида человека имеет форму эллипсоида, сжатого с боков, с одной из сторон имеется небольшая ямка, поэтому иногда говорят о «ложковидной» форме головки сперматозоида у человека. В головке сперматозоида располагаются следующие клеточные структуры:
Ядро, несущее одинарный набор хромосом. Такое ядро называют гаплоидным. После слияния сперматозоида и яйцеклетки (ядро которой также гаплоидно) образуется зигота — новый диплоидный организм, несущий материнские и отцовские хромосомы. При сперматогенезе (развитии сперматозоидов) образуются сперматозоиды двух типов: несущие X-хромосому и несущие Y-хромосому. При оплодотворении яйцеклетки X-несущим сперматозоидом формируется эмбрион женского пола. При оплодотворении яйцеклетки Y-несущим сперматозоидом формируется эмбрион мужского пола. Ядро сперматозоида значительно мельче ядер других клеток, это во многом связано с уникальной организацией строения хроматина сперматозоида (см. протамины). В связи с сильной конденсацией хроматин неактивен — в ядре сперматозоида не синтезируется РНК.
Акросома — видоизмененная лизосома — мембранный пузырек, несущий литические ферменты — вещества, растворяющие оболочку яйцеклетки. Акросома занимает около половины объёма головки и по своему размеру приблизительно равна ядру. Она лежит спереди от ядра и покрывает собой половину ядра (поэтому часто акросому сравнивают с шапочкой). При контакте с яйцеклеткой акросома выбрасывает свои ферменты наружу и растворяет небольшой участок оболочки яйцеклетки, благодаря чему образуется небольшой «проход» для проникновения сперматозоида. В акросоме содержится около 15 литических ферментов, основным из который является акрозин.
Центросома — центр организации микротрубочек, обеспечивает движение хвоста сперматозоида, а также предположительно участвует в сближении ядер зиготы и первом клеточном делении зиготы.
Позади головки располагается так называемая «средняя часть» сперматозоида. От головки среднюю часть отделяет небольшое сужение — «шейка». Позади средней части располагается хвост. Через всю среднюю часть сперматозоида проходит цитоскелет жгутика, который состоит из микротрубочек. В средней части вокруг цитоскелета жгутика располагается митохондрион — гигантская митохондрия сперматозоида. Митохондрион имеет спиральную форму и как бы обвивает цитоскелет жгутика. Митохондрион выполняет функцию синтеза АТФ и тем самым обеспечивает движение жгутика.
Хвост, или жгутик, расположен за средней частью. Он тоньше средней части и значительно длиннее её. Хвост — орган движения сперматозоида. Его строение типично для клеточных жгутиков эукариот. 17.
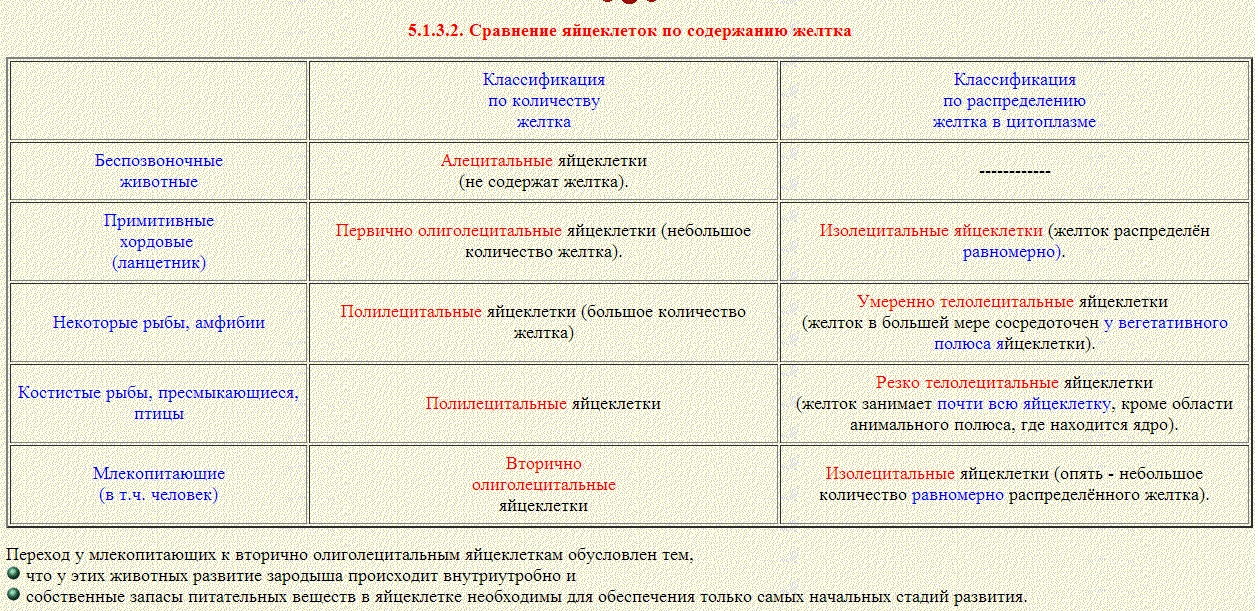
Яйцеклетка (науч. ооцит, реже. овоцит) — женская гамета животных, высших растений, а также многих водорослей и другихпротистов, которым свойственна оогамия. Как правило, яйцеклетки — гаплоидные клетки, но могут иметь другую плоидность уполиплоидных организмов.
Человеческая яйцеклетка имеет диаметр примерно 130 мкм и весит 1.15 нанограмм.
В цитоплазме яйцеклеток (ооплазме) содержатся совокупность питательных веществ — желток.
Яйцеклетки образуются в результате оогенеза. После оплодотворения из оплодотворенной яйцеклетки (зиготы) развиваетсяэмбрион. При партеногенезе эмбрион, а затем новый организм развивается из неоплодотворенной яйцеклетки.
Строение яйцеклетки представлено на рисунке.
Ядро
Цитолемма
Фолликулярный эпителий
Лучистый венец
Кортикальные гранулы
Желточные включения
Блестящая зона
Рецептор во фракции Zp 3-N-ацетилглюкозоамин
Яйцеклетка перемещается в ампуле реснитчатым движением клеток эпителия и ритмичной перистальтикой маточной трубы.
Яйцеклетка окружена прозрачной оболочкой (zona pellucida) и лучистым венцом (corona radiata) несет 23 хромосомы.
Особенности строения половой клетки
Строение яйцеклетки во многом обусловлено содержанием в ней компонентов, необходимых для осуществления биосинтеза белка. К ним относятся рибосомы, ферменты, т-РНК, м-РНК и их предшественники. Особые регуляторные вещества, контролирующие процессы, происходящие с самой яйцеклеткой в разный период ее жизни, играют важнейшую роль в дезинтеграции оболочки ядра. Желток яйцеклетки содержит белки, различные жиры, фосфолипиды, минеральные соли. Он обеспечивает полноценное питание зародыша. Особенности строения яйцеклетки: ее ядро содержит гаплоидный набор хромосом (23), а в ее цитоплазме находится большое количество митохондрий, элементы эндоплазматического ретикулума, свободные рибосомы, желточные включения и РНК. По периферии ядра располагаются кортикальные гранулы.
Строение яйцеклетки человека
Яйцеклетка имеет специфические оболочки, выполняющие защитные функции и препятствующие проникновению в нее нескольких сперматозоидов. Они также облегчают имплантацию зародыша в стенке матки. Яйцеклетка чаще всего имеет шарообразную форму. Она содержит весь набор таких типичных органелл, какие имеет любая клетка. Яйцеклетка окружена плазматической мембраной, а снаружи покрыта блестящей оболочкой, которая состоит из мукополисахаридов. На ней находится так называемый лучистый венец или по-другому – фолликулярная оболочка, представляющая собой микроскопические ворсинки фолликулярной клетки. Она играет важнейшую защитную и питательную функцию. Только воздействие нескольких сперматозоидов способно разрушить внешнюю оболочку яйцеклетки. После такой «атаки» только один или в крайнем случае двое из них оплодотворяют ее. Яйцеклетка не имеет своего аппарата активного движения. На преодоление расстояния в 10 см, которое она проходит по яйцеводу до матки, ей требуется 4-7 дней. Яйцеклетке присуща плазматическая сегрегация, означающая то, что с момента оплодотворения до дробления яйца в ней происходит равномерное распределение цитоплазмы. Благодаря этому процессу в дальнейшем все клетки будущих тканей будут получать ее в необходимом количестве.
18. Сперматогене́з — развитие мужских половых клеток (сперматозоидов), происходящее под регулирующим воздействием гормонов. Одна из форм гаметогенеза.
Сперматозоиды развиваются из клеток-предшественников, которые проходят редукционные деления (деления мейоза) и формируют специализированные структуры (акросома, жгутик и пр.). В разных группах животных сперматогенез различается. У позвоночных животных сперматогенез проходит по следующей схеме: в эмбриогенезе первичные половые клетки — гоноциты мигрируют в зачаток гонады, где формируют популяцию клеток, называемых сперматогониями. С началом полового созреваниясперматогонии начинают активно размножаться, часть из них дифференцируется в другой клеточный тип —сперматоциты I порядка, которые вступают в мейоз и после первого деления мейоза дают популяцию клеток, называемых сперматоцитами II порядка, проходящих впоследствии второе деление мейоза и образующих сперматиды; путём ряда преобразований последние приобретают форму и структуры сперматозоида в ходе спермиогенеза.
Сперматогенез у человека
Схема поперечного сечения семенного канальца
Сперматогенез у человека в норме начинается в пубертатном периоде (около 12 лет) и продолжается до глубокой старости. Продолжительность полного сперматогенеза у мужчин составляет примерно 73—75 дней. Один цикл зародышевого эпителия составляет приблизительно 16 дней
Сперматозоиды образуются в яичках, а именно в извитых семенных канальцах. Стенка семенного канальца делится базальной мембраной на люминальную и адлюминальную стороны. На люминальной стороне расположены клетки Сертоли (сустентоциты) и предшественники половых клеток (сперматогонии, сперматоциты I и II порядков и сперматиды).
Сперматогонии, лежащие непосредственно на базальной мембране извитых семенных канальцев, проходят несколько последовательных стадий митотического деления. Общее количество сперматогоний в яичке мужчины составляет около 1 млрд. Различают две основные категории сперматогоний: А и В. Сперматогонии А, которые делятся митотически, сохраняют способность к делению и поддерживают свою популяцию. Остальные дефференцируются в сперматогоний В, которые «эвакуируются» клеточными контактами сустентоцитов (образуют под основанием половой клетки новый контакт и резорбируют старый). Сперматогония В делится митотически, дифференцируясь в сперматоцит I порядка, вступающий в мейоз.
В результате первого деления мейоза образуются две дочерние клетки сперматоциты второго порядка, каждый из которых содержит гаплоидный набор (23 у человека) d-хромосом . Вторичные сперматоциты расположены ближе к просвету канальца. Во втором делении мейоза образуются две сперматиды. Таким образом, в результате деления одной сперматогонии образуются четыре сперматиды, каждая из которых обладает гаплоидным набором хромосом.
В ходе сложного процесса спермиогенеза сперматиды дифференцируются в зрелые сперматозоиды. Дифференцирующиеся сперматиды лежат в углублениях плазматической мембраны клеток Сертоли. При спермиогенезе комплекс Гольджи формирует акросому, содержащую протеолитические ферменты , которые при контакте с яйцеклеткой растворяют участок её блестящей оболочки (zona pellucida).
Сложный процесс сперматогенеза регулируется гонадотропными гормонами гипофиза и стероидными гормонами яичка. После полового созревания гипоталамус начинает выделять гонадотропный рилизинг-гормон, под влиянием которого гипофиз секретирует фолликулостимулирующий гормон (ФСГ), стимулирующий развитие и функционирование клетки Сертоли и лютеинизирующий гормон (ЛГ), стимулирующий клетки Лейдига к выработке тестостерона. Тестостерон оказывает воздействие на развитие клеток Сертоли, а также на предшественники половых клеток (в ассоциации с андроген-связывающим белком, выделяемым клетками Сертоли).
Секреторная активность гипофиза регулируется клетками Сертоли и клетками Лейтига. Тестостерон, выделяемый клетками Лейдига подавляет активность гипофиза к выработке ЛГ и ФСГ. Ингибин и эстрадиол, образующийся в клетках Сертоли, подавляют гипофиз к выработке ФСГ и клетки Лейдига к выработке тестостерона. Морфофункциональное состояние яичка регулируется гормонами аденогипофиза — ФСГ и ЛГ, причём уровень гормонов постоянный, имеются лишь незначительные колебания.
19. Овогенез — это процесс образования женских половых гамет, идет по той же схеме, что исперматогенез, но с некоторыми существенными отличиями. В результате неравномерного распределения цитоплазмы как при первом, так и при втором делениях мейоза только в одной клетке оказывается большой запас питательных веществ, необходимых для развития будущего зародыша. Следовательно, образуется только одна зрелая яйцеклетка с гаплоидным набором хромосом (n) и три маленькие клеточки, которые впоследствии исчезают. При овогенезе наряду с мейозом происходит так называемоесозревание яйцеклетки, во время которого значительно увеличивается ее объем.
Овогенез
(лат. ovum - яйцо + греч. genesis-зарождение, происхождение, развитие), процесс развития женских половых клеток (гамет), заканчивающийся формированием яйцеклеток. У женщины в течение менструального цикла созревает лишь одна яйцеклетка. Процесс овогенеза имеет принципиальное сходство со сперматогенезом и также проходит через ряд стадий: размножения, роста и созревания. Яйцеклетки образуются в яичнике, развиваясь из незрелых половых клеток -овогониев, содержащих диплоидное число хромосом. Овогонии, подобно сперматогониям, претерпевают последовательные митотические деления, которые завершаются к моменту рождения плода. Затем наступает период роста овогониев, когда их называют овоцитами I порядка. Они окружены одним слоем клеток - гранулёзной оболочкой - и образуют так называемые примордиальные фолликулы. Плод женского пола накануне рождения содержит около 2 млн. этих фолликулов, но лишь примерно 450 из них достигают стадии овоцитов II порядка и выходят из яичника в процессе овуляции. Созревание овоцита сопровождается двумя последовательными делениями, приводящими к уменьшению числа хромосом в клетке вдвое. В результате первого деления, мейоза, образуется крупный овоцит II порядка и первое полярное тельце, а после второго деления - зрелая, способная к оплодотворению и дальнейшему развитию яйцеклетка с гаплоидным набором хромосом и второе полярное тельце. Полярные тельца, представляющие собой мелкие клетки, не играют роли в овогенезе и в конечном счёте разрушаются. В отличие от образования спермиев у мужчин, которое начинается только в период полового созревания, образование яйцеклеток у женщин начинается ещё до их рождения и завершается для каждой данной яйцеклетки только после её оплодотворения. Поэтому любые неблагоприятные факторы внешней среды, начиная со стадии внутриутробного развития девочки, могут повлечь за собой генетические аномалии у её потомства. 20.
Половой процесс, или оплодотворение, или амфимиксис (др.-греч. ἀμφι- — приставка со значением обоюдности, двойственности и μῖξις — смешение) — процесс слияния гаплоидных половых клеток, или гамет, приводящий к образованию диплоидной клетки зиготы. Не следует смешивать это понятие с половым актом (встречей половых партнёров у многоклеточных животных).
Половой процесс закономерно встречается в жизненном цикле всех организмов, у которых отмечен мейоз. Мейоз приводит к уменьшению числа хромосом в два раза (переход от диплоидного состояния к гаплоидному), половой процесс — к восстановлению числа хромосом (переход от гаплоидного состояния к диплоидному).
Различают несколько форм полового процесса:
изогамия — гаметы не отличаются друг от друга по размерам, подвижны, жгутиковые или амебоидные;
анизогамия (гетерогамия) — гаметы отличаются друг от друга по размерам, но оба типа гамет (макрогаметы и микрогаметы) подвижны и имеют жгутики;
оогамия — одна из гамет (яйцеклетка) значительно крупнее другой, неподвижна, деления мейоза, приводящие к её образованию, резко асимметричны (вместо четырёх клеток формируется одна яйцеклетка и два абортивных «полярных тельца»); другая (спермий, или сперматозоид) подвижна, обычно жгутиковая или амебоидная.
Биологическое значение амфимиксиса непосредственно связано с биологической сущностью определенных сторон процесса оплодотворения. Дарвин, открывший «великий закон природы», говорил о прогрессивном значении появления полового процесса в истории органического мира, рассматривая при этом перекрёстное опыление как источник обогащения наследственности. Благодаря бипариентальному наследованию (материнское — от яйцеклетки и отцовское — от спермия) в результате амфимиксиса получаются более жизнеспособные организмы, обладающие более широким спектром изменчивости по сравнению с апомиктичными растениями
Оплодотворение
слияние мужской половой клетки (сперматозоида) с женской (яйцом, яйцеклеткой), приводящее к образованию зиготы - нового одноклеточного организма. Биологический смысл оплодотворения состоит в объединении ядерного материала мужской и женской гамет, что приводит к объединению отцовских и материнских генов, восстановлению диплоидного набора хромосом, а также активации яйцеклетки, то есть стимуляции её к зародышевому развитию. Соединение яйцеклетки со сперматозоидом обычно происходит в воронкообразно расширенной части маточной трубы в течение первых 12 часов после овуляции. Семенная жидкость, попадая во влагалище женщины при половом сношении, обычно содержит от 60 до 150 млн. сперматозоидов, которые, благодаря движениям со скоростью 2-3 мм в минуту, постоянным волнообразным сокращениям матки и труб и щелочной среде, уже спустя 1-2 минуты после полового акта достигают матки, а через 2-3 часа - концевых отделов маточных труб, где обычно и происходит слияние с яйцеклеткой. Различают моноспермное (в яйцеклетку проникает один сперматозоид) и полиспермное (в яйцеклетку проникают два и более сперматозоидов, но с ядром яйцеклетки сливается только одно ядро сперматозоида) оплодотворение. Сохранению активности спермиев во время прохождения их в половых путях женщины способствует слабощелочная среда шеечного канала матки, заполненного слизистой пробкой. Во время оргазма при половом акте слизистая пробка из шеечного канала частично выталкивается, а затем вновь втягивается в него и тем самым способствует более быстрому попаданию сперматозоидов из влагалища (где в норме у здоровой женщины среда слабокислая) в более благоприятную среду шейки и полости матки. Прохождению сперматозоидов через слизистую пробку шеечного канала способствует и резко повышающаяся в дни овуляции проницаемость слизи. В остальные дни менструального цикла слизистая пробка имеет значительно меньшую проницаемость для сперматозоидов. Многие сперматозоиды, находящиеся в половых путях женщины, могут сохранять способность к оплодотворению 48-72 часа (иногда даже до 4-5 суток). Овулировавшая яйцеклетка сохраняет жизнеспособность примерно 24 часа. Учитывая это, наиболее благоприятным временем для оплодотворения считается период разрыва созревшего фолликула с последующим рождением яйцеклетки, а также 2-3-й день после овуляции. Женщинам, применяющим физиологический метод контрацепции, следует помнить о том, что сроки овуляции могут колеб
аться, а жизнеспособность яйцеклетки и сперматозоидов может быть значительно больше. Вскоре после оплодотворения начинается дробление зиготы и образование зародыша. В последние годы для лечения бесплодия применяется метод экстракорпорального оплодотворения - оплодотворение яйцеклетки человека вне организма, культивирование её до определённой стадии и перенос эмбриона (зародыша) в матку. Абсолютным показанием для этого метода оплодотворения является трубное бесплодие, которое развивается вследствие непроходимости или отсутствия маточных труб. В случаях мужского бесплодия применяют инструментальное введение в половые пути женщины спермы мужа или донора с целью обеспечения у неё зачатия (см. Инсеминация искусссвенная).
(Источник: Сексологический словарь)
(сингамия), слияние муж. и жен. половых клеток (гамет) у растений, животных и человека, в результате чего образуется зигота, способная развиваться в новый организм. О. лежит в основе полового размножения и обеспечивает передачу наследственных признаков от родителей потомкам.
21)
 22)
Дробле́ние —
ряд
последовательных митотических делений
оплодотворенного или инициированного
к развитию яйца.
Дробление представляет собой первый
период эмбрионального
развития,
который присутствует в онтогенезе всех
многоклеточных животных.
При этом масса зародыша и
его объём не меняются, оставаясь такими
же, как и в начале дробления.Яйцо разделяется
на все более мелкие клетки — бластомеры.
Характерная особенность дробления —
ведущая регуляторная роль цитоплазмы в развитии.
Характер дробления зависит от
количества желтка и
его расположения в яйце.
Голобластическое
дробление.
тип
равномерного дробления оплодотворенной
яйцеклетки, при котором дочерние клетки
имеют приблизительно одинаковые
размеры.
Голобластическое
дробление
22)
Дробле́ние —
ряд
последовательных митотических делений
оплодотворенного или инициированного
к развитию яйца.
Дробление представляет собой первый
период эмбрионального
развития,
который присутствует в онтогенезе всех
многоклеточных животных.
При этом масса зародыша и
его объём не меняются, оставаясь такими
же, как и в начале дробления.Яйцо разделяется
на все более мелкие клетки — бластомеры.
Характерная особенность дробления —
ведущая регуляторная роль цитоплазмы в развитии.
Характер дробления зависит от
количества желтка и
его расположения в яйце.
Голобластическое
дробление.
тип
равномерного дробления оплодотворенной
яйцеклетки, при котором дочерние клетки
имеют приблизительно одинаковые
размеры.
Голобластическое
дробление
характерно для гомолецитальных яиц. Плоскости дробления разделяют яйцо полностью. Они могут делить его на равные части, как у морской звезды или морского ежа, или же на неравные части, как у брюхоногого моллюска Crepidula. Дробление умеренно телолецитального яйца ланцетника происходит по голобластическому типу, однако неравномерность деления проявляется только после стадии четырех бластомеров. У некоторых клеток после этой стадии дробление становится крайне неравномерным; образующиеся при этом мелкие клетки называют микромерами, а крупные клетки, содержащие желток, – макромерами. У моллюсков плоскости дробления проходят таким образом, что начиная со стадии восьми клеток бластомеры располагаются по спирали; этот процесс регулируется ядром.
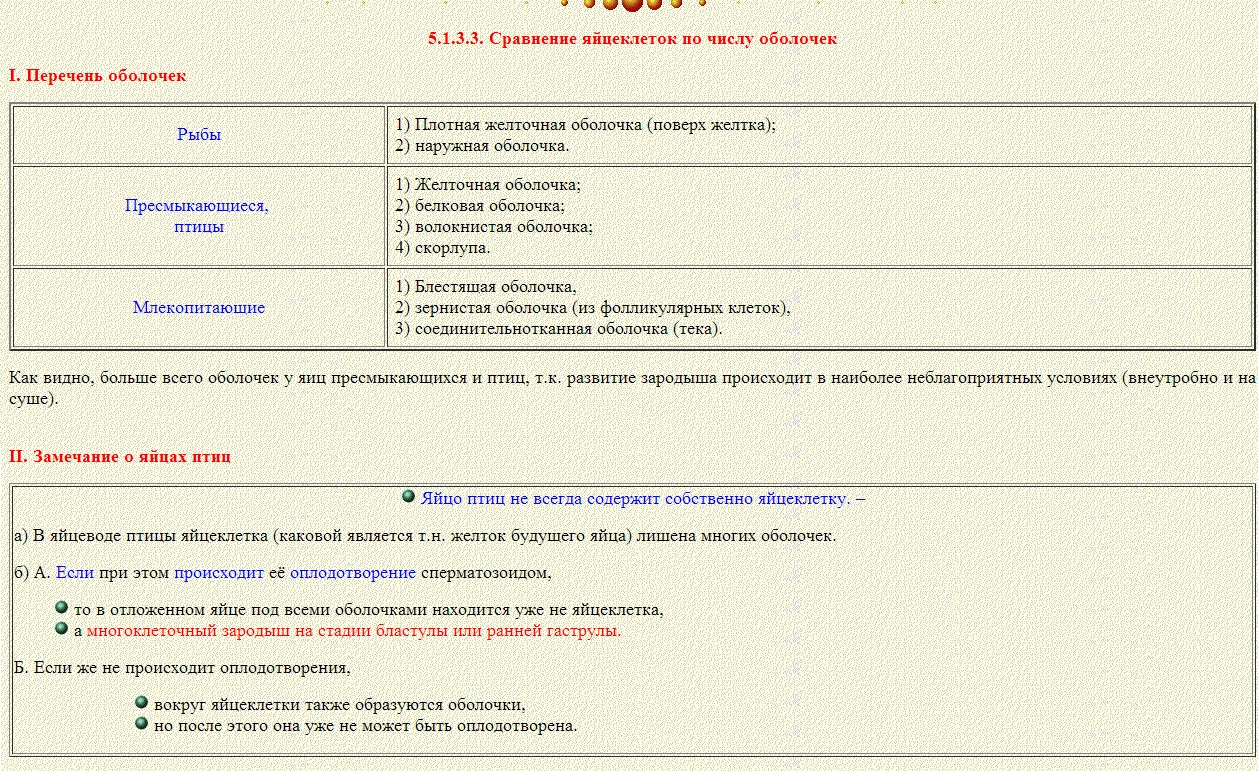
23) Меробластическое дробление
типично для телолецитальных яиц, богатых желтком; оно ограничено относительно небольшим участком у анимального полюса. Плоскости дробления не проходят через все яйцо и не захватывают желток, так что в результате деления на анимальном полюсе образуется небольшой диск клеток (бластодиск). Такое дробление, называемое также дискоидальным, свойственно пресмыкающимся и птицам.
meroblastic cleavage - меробластическое дробление.
Hеполное (частичное) дробление телолецитальных (у головоногих моллюсков, акул, пресмыкающихся, птиц, однопроходных млекопитающих) и центролецитальных яиц.
24)Дискоидальное дробление (от греч. dískos - диск и éidos - вид), один из типов дробления яиц у животных с телолецитальными яйцами (скорпионы, головоногие моллюски, хрящевые и костистые рыбы, пресмыкающиеся и птицы). При Дискоидальное дроблениеделится лишь небольшой диск относительно свободной от желтка и содержащей ядро цитоплазмы. 25) Бластуля́ция (от др.-греч. blastós — «зародыш, росток») — заключительный этап процесса дробления яйца у многоклеточных, в течение которого происходит образование бластулы.
В период бластуляции поверхностные бластомеры образуют эпителиоподобный пласт, возникает центральная полость — бластоцель. Дробление клеток становится асинхронным, продолжительность митотического цикла возрастает за счёт удлинения интерфазы. Происходят изменения в структуре клеток: в интерфазных ядрах появляются ядрышки, усложняется структура митохондрий, образуются эндоплазматическая сеть и специализированные межклеточные контакты. В ядрах активизируется синтез информационной РНК, что обеспечивает переход к гаструляции.
26) Гаструляция — сложный процесс морфогенетических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате чего образуются зародышевые листки (эктодерма,мезодерма и энтодерма) — источники зачатков тканей и органов. Второй после дробления этап онтогенеза. При гаструляции происходит перемещение клеточных масс с образованием из бластулы двухслойного или трёхслойного зародыша — гаструлы.
Тип бластулы определяет способ гаструляции.
Зародыш на этой стадии состоит из явно разделенных пластов клеток — зародышевых листков: наружного (эктодерма) и внутреннего (энтодерма).
У многоклеточных животных, кроме кишечнополостных, параллельно с гаструляцией или, как у ланцетника, вслед за ней возникает и третий зародышевый листок — мезодерма, который представляет собой совокупность клеточных элементов, расположенных между эктодермой и эндодермой. Вследствие появления мезодермы зародыш становится трехслойным.
У многих групп животных именно на стадии гаструляции появляются первые признаки дифференцировки. Дифференцировка(дифференциация) — процесс возникновения и нарастания структурных и функциональных различий между отдельными клетками и частями зародыша.
Из эктодермы образуется нервная система, органы чувств, эпителий кожи, эмаль зубов; из энтодермы — эпителий средней кишки, пищеварительные железы, эпителий жабр и легких; из мезодермы — мышечная ткань, соединительная ткань, кровеносная система, почки, половые железы и др.
У разных групп животных одни и те же зародышевые листки дают начало одним и тем же органам и тканям.
Способы гаструляции
Инвагинация — происходит путем впячивания стенки бластулы в бластоцель; характерна для большинства групп животных.
Деляминация (характерна для кишечнополостных) — клетки, находящиеся снаружи, преобразуются в эпителиальный пласт эктодермы, а из оставшихся клеток формируется энтодерма. Обычно деляминация сопровождается делениями клеток бластулы, плоскость которых проходит «по касательной» к поверхности.
Иммиграция — миграция отдельных клеток стенки бластулы внутрь бластоцеля.
Униполярная — на одном участке стенки бластулы, обычно на вегетативном полюсе;
Мультиполярная — на нескольких участках стенки бластулы.
Эпиболия — обрастание одних клеток быстро делящимися другими клетками или обрастание клетками внутренней массы желтка (при неполном дроблении).
Инволюция — вворачивание внутрь зародыша увеличивающегося в размерах наружного пласта клеток, который распространяется по внутренней поверхности остающихся снаружи клеток.
Типы гаструляции
При гаструляции продолжаются изменения, начавшиеся на стадии бластулы, и поэтому разным типам бластул соответствуют и различныетипы гаструляции. Переход из бластулы в гаструлу может осуществляться 4-мя основными способами: инвагинацией, иммиграцией, деляминацией и эпиболией. Инвагинация или впячивание наблюдается в случае целобластулы. Это наиболее простой способ гаструляции, при котором вегетативная часть впячивается в бластоцель. Вначале появляется небольшое углубление в вегетативном полюсе бластулы. Затем клетки вегетативного полюса все больше и больше впячиваются в полость бластоцеля. В последующем эти клетки доходят до внутренней стороны анимального полюса. Первичная полость, бластоцель, при этом вытесняется и видна только с двух сторон гаструлы в местах изгиба клеток. Зародыш принимает куполообразную форму и становится двухслойным. Его стенка состоит из наружного листка – эктодермы и внутреннего – энтодермы. В результате гаструляции образуется новая полость – гастроцель или полость первичной кишки. Она сообщается с внешней средой с помощью кольцеобразного отверстия – бластопора или первичного рта. Края бластопора называются губами. Различают спинную, брюшную и две боковых губы бластопора. По последующей судьбе бластопора всех животных разделяют на две большие группы: первично- и вторичноротых. К первичноротым относят животных, у которых бластопор остается постоянным или дефинитивным ртом у взрослой особи (черви, моллюски, членистоногие). У других животных (иглокожие, хордовые) бластопор или превращается в заднепроходное отверстие, или зарастает, а ротовое отверстие возникает заново на переднем конце тела зародыша. Таких животных называют вторичноротыми. Иммиграция или вселение является наиболее примитивной формой гаструляции. При этом способе происходит перемещение отдельных клеток или группы клеток из бластодермы в бластоцель с образованием энтодермы. Если вселение клеток в бластоцель происходит лишь со стороны одного полюса бластулы, то такая иммиграция называется униполярной, а с различных участков бластулы – мультиполярной. Униполярная иммиграция свойственна некоторым гидроидным полипам, медузам и гидромедузам. В то время, как мультиполярная иммиграция является более редким явлением и наблюдается у некоторых гидромедуз. При иммиграции внутренний зародышевый листок – энтодерма может образовываться сразу в процессе проникновения клеток в полость бластоцеля. В других случаях клетки могут заполнять полость сплошной массой, а затем выстраиваться упорядоченно возле эктодермы и образовывать энтодерму. В последнем случае гастроцель появляется позднее. Деляминация или расслаивание сводится к расщеплению стенки бластулы. Клетки, которые отделяются внутрь, образуют энтодерму, а наружные – эктодерму. Такой способ гаструляции наблюдается у многих беспозвоночных и высших позвоночных животных. У некоторых животных в связи с увеличением количества желтка в яйцеклетке и уменьшением полости бластоцеля гаструляция только путем инвагинации становится невозможной. Тогда гаструляция происходит способом эпиболии или обрастания. Этот способ состоит в том, что мелкие анимальные клетки усиленно делятся и обрастают вокруг более крупных вегетативных. Мелкие клетки образуют эктодерму, а клетки вегетативного полюса формируют энтодерму. Такой способ гаструляции наблюдается у круглоротых и земноводных. 27)-29) нету вообще 30) Образование осевых органов у ланцетника
После отделения хорды и мезодермы края энтодермы постепенно сближаются в спинной части и в конце концов смыкаются, образуя замкнутую кишечную трубку. В ходе дальнейшего развития мезодерма сегментируется: тяжи разделяются поперечно на первичные сегменты или сомиты. Из них образуются три основные закладки: дерматом формируется из наружной, обращенной к эктодерме стенке сомита, – из его клеток впоследствие возникает соединительная часть кожи, представленная преимущественно фибробластами; склеротом образуется из внутренней части сомита, примыкающей к хорде (низшие позвоночные) или к хорде и нервной трубке (высшие позвоночные) – представляет зачаток осевого скелета; миотом представляет часть сомита, расположенную между дерматомом и склеротомом – является зачатком всей поперечно-полосатой мускулатуры. Дифференцировка сомитов у ланцетника протекает иначе, чем у позвоночных. Это различие выражается в том, что у позвоночных сегментируется только спинная часть мезодермальных тяжей, тогда как у ланцетника они полностью распадаются на сегменты. Последние вскоре разделяются на спинную часть – сомиты, и брюшную – спланхнотом. Сомиты, из которых развивается туловищная мускулатура, остаются обособленными друг от друга, спланхнотомы же сливаются на каждой стороне, образуя левую и правую полости, которые затем объединяются под кишечной трубкой в общую вторичную полость тела (целом). В развитии ланцетника, с одной стороны, ясно представлены черты типичных позвоночных (характерное расположение зачатков при гаструляции, формирование хорды из дорсальной стенки первичной кишки и нервной пластинки из дорсальной эктодермы), а с другой – черты беспозвоночных вторичноротых животных (целобластула, инвагинационная гаструла, трехсегментная стадия, энтероцельная закладка мезодермы и образования целома). В дальнейшем, в связи с образованием хвоста нервно-кишечный канал исчезает. В головной части кишечной трубки прорывается ротовое отверстие, а на заднем конце, под хвостом, образуется анальное – путем вторичного прорыва стенки тела животного на месте закрывшегося бластопора. Зародыш переходит в стадию свободноплавающей личинки. 31) Гаструляция протекает медленнее, чем у ланцетника, поэтому мезодерма не входит в состав первичной энтодермы, а, обособившись в виде самостоятельного зачатка, растет между эктодермой и энтодермой. В связи с этим кишечная энтодерма в начале гаструляции имеет вид незамкнутой трубки (рис. 51). Таким образом, при данном типе гаструляции первичная эктодерма отсутствует. 32)
К концу вторых суток инкубации гаструляция заканчивается и начинается следующая стадия развития – нейруляция или образование осевых органов. На ней зародыш курицы энтодермой прилегает к желтку. Над энтодермой находится хордальный и мезодермальный материал. Выше располагается нервная пластинка и эпидермальная эктодерма. Хордальный материал индуцирует материал эктодермы, лежащий над хордой, заставляет его утолщаться и превращаться в нервную пластинку. В ответ на индукционный стимул эктодермальные клетки, лежащие над хордой (головным отростком), синтезируют мРНК. Их дальнейшее развитие направлено на образование нервной ткани. Клетки, отвечающие на нейрональную индукцию морфологически меняются, превращаясь из кубических в высокие призматические. На стадии нейруляции края нервной пластинки утолщаются и приподнимаются кверху, образуя нервные валики. Вместе с краями нервной пластинки поднимается связанная с ней эпидермальная эктодерма. Постепенно образуется нервный желобок, превращающийся затем в нервную трубку при срастании краев эктодермы. Часть материала нервной пластинки не включается в нервную трубку и называется нервным гребнем или ганглиозной пластинкой. Замыкание нервной трубки и срастание нервных валиков распространяется от переднего конца тела в каудальном направлении. Одновременно с образованием нервной трубки хордальный материал уплотняется и превращается в хорду В мезодермальном материале выделяется дорсальная осевая часть – сомиты и вентральная – спланхнотомы или боковые пластинки, которые делятся на висцеральный и париетальный листки (как в зародышевой, так и внезародышевых частях бластодиска). Сомиты связаны с париетальными листками боковых пластинок нефротомами. Между париетальными и висцеральными листками находится вторичная полость тела – целом. В сомитах различают склеротом, дерматом, миотом. Из склеротома в последующем формируется осевой скелет, все виды соединительной ткани (собственно соединительная, хрящевая, костная). Из дерматома – собственно кожа, а из миотома – скелетная мускулатура. Энтодерма первоначально представлена совершенно плоской пластинкой клеток, распластанной над заполненной жидкостью подзародышевой полостью. Лишь позднее, когда зародыш начинает приподниматься над желтком и обособляться от внезародышевых частей бластодиска, энтодермальная пластинка выгибается кверху, образуя под хордой кишечный желобок. У птиц к моменту закладки осевых органов отсутствует кишечная трубка и нет вентральной стенки туловища. Они возникают в результате образования туловищных складок. |
33)
34)
ВНЕЗАРОДЫШЕВЫЕ ОРГАНЫ. |
развиваются в процессе эмбриогенеза вне тела зародыша и обеспечивают его нормальное развитие и рост. К ним относятся: Амнион (плодный пузырь) - обеспечивает водную среду для развития зародыша. Его стенка состоит из:
Функции амниотической жидкости: - защита плода от механических повреждений; - обеспечение нормального развития конечностей плода; - обеспечение нормального развития дыхательной, пищеварительной и мочевыделительной систем; Желточный мешок - пузырек, связанный с кишечной трубкой. Его стенка состоит из эпителия и соединительной ткани. Функции: - источник образования первых кровеносных сосудов и первых клеток крови; - источник гонобластов - предшественников половых клеток; - до 3-й недели эмбриогенеза - питательная. Аллантоис - пальцевидное выпячивание, которое врастает в амниотическую ножку. Это орган газообмена и выделения, так как по нему к хориону растут сосуды, располагающиеся в пупочном канатике. На 2-м месяце аллантоис редуцируется и как рудимент входит в состав пупочного канатика. Пупочный канатик или пуповина - это упругий тяж, соединяющий зародыш с плацентой. Состоит из:
Все эти компоненты окружены слизистой соединительной тканью (Вартонов студень), который предохраняет сосуды пуповины от сжатия. Хорион развивается из трофобласта. На ранних этапах хорион имеет первичные ворсинки, которые состоят из внутреннего слоя клеток - цитотрофобласта и наружного - симпластотрофобласта, который выделяет протеолитические ферменты, разрушающие слизистую оболочку матки и способствует имплантации. Когда в первичные ворсины врастает соединительная ткань - это уже вторичные ворсины. После врастания в них кровеносных капилляров - этотретичные ворсины. Часть ворсин хориона разрастается и образует ветвистый хорион (область плаценты), вторая большая часть ворсин образует гладкий хорион. Плацента - временный орган, обеспечивающий связь плода с организмом матери. Состоит из: 1) зародышевой части, представленной ветвистым хорионом и приросшей к нему амниотической оболочкой; 2) материнской части - видоизмененной слизистой (децидуальной) оболочкой матки. Содержит децидуальные клетки Структурно-функциональной единицей плаценты является - котиледон. Он образован стволовой ворсиной и ее разветвлениями, находится в лакуне, заполненной материнской кровью и отделен от соседних котиледонов соединительнотканными септами. Типы плацент: - эпителиохориальная (ворсины хориона контактируют с эпителием эндометрия матки) характерна для свиньи, верблюда, лошади. - десмохориальная (ворсины хориона разрушают эпителий и контактируют с подлежащей соединительной тканью эндометрия матки) характерна для жвачных. - эндотелиохориальная (ворсины хориона разрушают эпителий и соединительную ткань и контактируют с эндотелием сосудов эндометрия) характерна для хищников. - гемохориальная (ворсины хориона контактируют с кровью матери) характерна для человека, приматов. Функции плаценты: - дыхательная; - транспорт питательных веществ, воды, электролитов; - выделительная; - эндокринная (синтез гормонов - хорионического гонадотропина (появляется в крови уже на 3-4 сутки беременности), плацентарного лактогена, прогестерона, эстрогена). - защитная (плацентарный барьер). |
35) Плацента (лат. placenta, «лепёшка») — эмбриональный орган у всех самок плацентарных млекопитающих, некоторых сумчатых, рыбы-молот и других живородящиххрящевых рыб, а также живородящих онихофор и ряда других групп животных, позволяющий осуществлять перенос материала между циркуляционными системамиплода и матери;
Также в ботанике плацента — участок плодолистика, к которому прикрепляется семяпочка.
У млекопитающих плацента образуется из зародышевых оболочек плода (ворсинчатой, хориона, и мочевого мешка — аллантоиса (allantois)), которые плотно прилегают к стенке матки, образуют выросты (ворсинки), вдающиеся в слизистую оболочку, и устанавливают, таким образом, тесную связь между зародышем и материнским организмом, служащую для питания и дыхания зародыша[1]. Пуповина связывает эмбрион с плацентой.
Схематичное изображение места расположения плаценты в матке беременной женщины
Плацента вместе с оболочками плода (так называемый послед) у женщины выходит из половых путей через 5 - 60 минут (в зависимости от тактики ведения родов) после появления на свет ребёнка.
Образование плаценты
Строение плаценты
Плацента образуется чаще всего в слизистой оболочке задней стенки матки из эндометрия и цитотрофобласта. Слои плаценты (от матки к плоду — гистологически):
Децидуа — трансформированный эндометрий (с децидуальными клетками, богатыми гликогеном),
Фибриноид Рора (слой Лантганса),
Трофобласт, покрывающий лакуны и врастающий в стенки спиральных артерий, предотвращающий их сокращение,
Лакуны, заполненные кровью,
Синцитиотрофобласт (многоядерный симпласт, покрывающий цитотрофобласт),
Цитотрофобласт (отдельные клетки, образующие синцитий и секретирующие БАВ),
Строма (соединительная ткань, содержащая сосуды, клетки Кащенко-Гофбауэра — макрофаги),
Амнион (на плаценте больше синтезирует околоплодные воды, внеплацентарный — адсорбирует).
Между плодовой и материнской частью плаценты — базальной децидуальной оболочкой — находятся наполненные материнскойкровью углубления. Эта часть плаценты разделена децидуальными септами на 15-20 чашеобразных пространств (котиледонов). Каждый котиледон содержит главную ветвь, состоящую из пупочных кровеносных сосудов плода, которая разветвляется далее в множестве ворсинок хориона, образующих поверхность котиледона (на рисунке обозначена какVillus). Благодаря плацентарному барьеру кровоток матери и плода не сообщаются между собой. Обмен материалами происходит при помощи диффузии, осмоса или активного транспорта. С 3-й недели беременности, когда начинает биться сердце ребёнка, плод снабжаетсякислородом и питательными веществами через «плаценту». До 12 недель беременности это образование не имеет чёткой структуры, до 6 нед. — располагается вокруг всего плодного яйца и называется хорионом, «плацентация» проходит в 3-6 нед.
Функции
Плацента формирует гематоплацентарный барьер, который морфологически представлен слоем клеток эндотелия сосудов плода, их базальной мембраной, слоем рыхлой перикапиллярной соединительной ткани, базальной мембраной трофобласта, слоями цитотрофобласта и синцитиотрофобласта. Сосуды плода, разветвляясь в плаценте до мельчайших капилляров, образуют (вместе с поддерживающими тканями) ворсины хориона, которые погружены в лакуны, наполненные материнской кровью. Он обуславливает следующие функции плаценты.
Газообменная
Кислород из крови матери проникает в кровь плода по простым законам диффузии, в обратном направлении транспортируется углекислый газ.
Трофическая и выделительная
Через плаценту плод получает воду, электролиты, питательные и минеральные вещества, витамины; также плацента участвует в удалении метаболитов (мочевины, креатина, креатинина) посредством активного и пассивного транспорта;
Гормональная
Плацента играет роль эндокринной железы: в ней образуются хорионический гонадотропин, поддерживающий функциональную активность плаценты и стимулирующий выработку больших количеств прогестерона жёлтым телом; плацентарный лактоген, играющий важную роль в созревании и развитии молочных желез во время беременности и в их подготовке к лактации; пролактин, отвечающий за лактацию; прогестерон, стимулирующий рост эндометрия и предотвращающий выход новых яйцеклеток; эстрогены, которые вызывают гипертрофию эндометрия. Кроме того, плацента способна секретировать тестостерон, серотонин, релаксин и другие гормоны.
Защитная
Плацента обладает иммунными свойствами — пропускает к плоду антитела матери, тем самым обеспечивая иммунологическую защиту. Часть антител проходят через плаценту, обеспечивая защиту плода. Плацента играет роль в регуляции и развитии иммунной системы матери и плода. В то же время она предупреждает возникновениеиммунного конфликта между организмами матери и ребёнка — иммунные клетки матери, распознав чужеродный объект, могли бы вызвать отторжение плода. Cинцитий поглощает некоторые вещества, циркулирующие в материнской крови, и препятствует их поступлению в кровь плода. Однако плацента не защищает плод от некоторыхнаркотических веществ, лекарств, алкоголя, никотина и вирусов.
Плацента человека
Человеческая плацента через несколько минут после родов
Плацента человека — placenta discoidalis, плацента гемохориального типа: материнская кровь циркулирует вокруг тонких ворсин, содержащих плодовые капилляры.
В отечественной промышленности с 30-х годов разработаны проф. В. П. Филатовым и выпускаются препараты экстракт плаценты и взвесь плаценты. Препараты плаценты активно используются в фармакологии.
Из пуповинной крови и плаценты можно получать стволовые клетки[2], хранящиеся в Банках пуповинной крови.
Плацентарные экстракты обладают также антибактериальным и противовирусным действиями. Действие препарата из плаценты сочетается с обеспечением организма необходимыми субстратами (витаминами, аминокислотами), что позволяет осуществлять стимуляцию организма без истощения его энергетических, пластических и других ресурсов. Наличие в плаценте аминокислот, ферментов, микроэлементов и уникальных биологически активных веществ, в особенности белков-регуляторов позволяет препаратам из плаценты активировать «спящие» клетки взрослого организма, что приводит к их размножению, обновлению клеточного состава, и в конечном итоге — к омоложению[источник не указан 319 дней].
В некоторых странах плаценту предлагают забрать домой, чтобы закопать её под деревом — этот обычай распространён в самых разных регионах мира[источник не указан 319 дней].
Плацента животных
Послед овцы
Существует несколько типов плаценты у животных. У сумчатых — неполная плацента, что обуславливает столь непродолжительный период беременности (8—40 дней). У парнокопытных — placenta diffusa эпителиохориального типа, placenta zonaria у хищников (эндотелиохорального типа), placenta discoid (гемохориальный тип) у грызунов и человека и placenta cotyledonaria или multiplex у жвачных.
Большинство самок млекопитающих, включая растительноядных (коровы и прочие жвачные), поедают свой послед сразу после облизывания новорождённого. Они делают это не только для того, чтобы уничтожить запах крови, привлекающий хищников, но и с целью обеспечения себя витаминами и питательным веществами, в которых они нуждаются после родов Эпителиохориальная плацента, или полуплацента, имеет наиболее простую структуру. Такой плацентой обладают свиньи, лошади, верблюды и некоторые другие млекопитающие. При образовании ее на поверхности хориона появляются ворсинки в форме небольших бугорков. Они как бы погружаются в соответствующие углубления слизистой оболочки матки, не производя никаких разрушений в ее тканях. При родах ворсинки выходят из своих углублений без всяких нарушений матки. Роды проходят безболезненно и без кровотечений. Десмохориальная плацента, или соединительнотканная, свойственна жвачным. Она характеризуется установлением более тесной связи хориона зародыша со стенкой матки. В месте соприкосновения с ворсинками хориона эпителий слизистой оболочки матки разрушается. Разветвленные ворсинки погружаются в соединительную ткань, приближаясь, таким образом, к кровеносным сосудам матери. Эндотелиохориальная плацента характеризуется разрушениями не только эпителия слизистой матки, но и соединительной ткани. Ворсинки хориона соприкасаются с сосудами и отделены от материнской крови только их тонкой эндотелиальной стенкой. Такой плацентой обладают хищники. Гемохориальная плацента свойственна насекомоядным, грызунам, всем приматам и человеку. При установлении контакта плода с материнским организмом происходят глубокие изменения в матке: частично разрушается соединительная ткань и даже стенки сосудов. На месте разрушенных тканей образуются большие полости, наполненные кровью, которая изливается из сосудов. Ворсинки хориона омываются кровью и всасывают из нее питательные вещества. Прямого сообщения между сосудами ворсинок и матки нет, и весь обмен веществ совершается через сильно утонченную стенку ворсинки. Вследствие тесного контакта между зародышем и материнским организмом роды сопровождаются отторжением значительной части стенки матки и обильными кровотечениями. 36) В настоящее время твердо установлено, что патология новорожденного часто обусловлена неблагоприятными воздействиями в период его внутриутробного развития. Знания о стадиях развития и росте плода необходимы для того, чтобы распознать и избежать возможных осложнений, которые могут возникнуть во время беременности.
РОСТ И СОЗРЕВАНИЕ ПЛОДА
Беременность в среднем продолжается 280 дней, или 10 акушерских месяцев от первого дня последней менструации (продолжительность акушерского месяца 28дней; 10 акушерских месяцев ровняются 40 неделям). В течение этого времени из оплодотворенной яйцеклетки развивается зрелый плод, способный к внеутробному существованию.
Во внутриутробном развитии человека различают (условно) два периода: зародышевый и плодовый.
ЭМБРИОНАЛЬНЫЙ ПЕРИОД
Продолжается от момента оплодотворения до конца второго месяца беременности. В этот период образуются зачатки всех важнейших органов и систем (нервная, кроветворная, сердечно-сосудистая, пищеварительная, выделительная и др.); происходит формирование туловища, головы, лица, зачатков конечностей. Зародыш приобретает черты, характерные для человека. Процессы развития в этот период весьма интенсивны, приспособительные механизмы еще не развиты, поэтому зародыш очень чувствителен к действию повреждающих факторов.
Недостаток кислорода, перегревание, микробы, вирусы, химические вещества, алкоголь, никотин - могут вызвать нарушение развития и гибель эмбриона или появление врожденных уродств.
ПЛОДОВЫЙ ПЕРИОД
Начинается с конца второго - начала третьего месяца беременности и продолжается до момента рождения ребенка. В этот период плод быстро растет, происходит развитие тканей, органов и систем, находящихся в зачаточном состоянии, становление новых функциональных систем, обеспечивающих жизнедеятельность плода в период его внутриутробной жизни и после появления на свет.
Развитие плода происходит в непосредственной взаимосвязи с организмом матери, в котором участвуют биохимические, иммунные, эндокринные, нервные и другие механизмы.
Организм женщины во время беременности находится под воздействием разнообразных факторов окружающей среды, которые, как уже говорилось, влияют на течение беременности и в ряде случаев вызывают развитие той или иной патологии плода. Однако реакция плода на повреждающие факторы зависит от стадии его развития и чувствительности его тканей к их воздействию.
В эмбриональном и плодовом периодах принято выделять стадии, когда зародыш обладает высокой чувствительностью к повреждающим веществам. Эти периоды получили название критических. Выделение этих периодов очень важно. Критические периоды развития характеризуются повышением обменных процессов (в эти периоды создаются все необходимые условия для осуществления одного из этапов развития зародыша в целом, а также отдельных зачатков органов и даже клеток).
Патогенные действия, которые могут привести во время беременности к нарушению развития плода, имеют самый разнообразный характер: переохлаждение, перегревание, недостаток кислорода, ионизирующее излучение, химические вещества, возбудители инфекции.
ПЕРВЫЙ КРИТИЧЕСКИЙ ПЕРИОД
Предшествует имплантации (прикрепление зародыша к стенке матки) или совпадает с ней. Это так называемая стадия предимплантационного развития (начинается с момента оплодотворения и продолжается до момента прикрепления зародыша к стенке матки - 7-8 дней после оплодотворения). Зародыш, на данной стадии, относительно устойчив к действию повреждающих агентов. Повреждающее действие реализуется, обычно, по принципу «все или ничего». Однако в конце предимплантационного периода наблюдается кратковременный период подъема чувствительности зародыша к повреждающим агентам. Именно в это время неблагоприятные факторы оказывают эмбриотоксическое действие, приводя к гибели зародыша.
ВТОРОЙ КРИТИЧЕСКИЙ ПЕРИОД
После завершения имплантации в развитии эмбриона начинается второй критический период – период образования зачатков органов и систем эмбриона, который у человека заканчивается к третьему месяцу внутриутробной жизни. Этот срок является наиболее чувствительным для развития эмбриона (3-7 недель), а также период образования плаценты (9-12 недель). В период образования органов, в результате патогенного действия повреждающих факторов на эмбрион, в первую очередь, поражаются те органы и системы, которые находились в это время в процессе закладки и повышенного обмена веществ. Поэтому для поражения эмбриона в этот период характерным является возникновение уродств (тератогенный эффект). У эмбриона человека наиболее чувствительными к неблагоприятным факторам являются центральная нервная система, органы зрения, железы внутренней секреции и половые железы, поэтому аномалии этих органов встречаются чаще других.
Под воздействием повреждающих факторов во время образования плаценты происходят патологические изменения, в основе которых лежит возникновение плацентарной недостаточности, выражением которой является врожденная гипотрофия плода (дефицит массы тела ребенка).
После завершения процессов формирования органов и формирования плаценты начинается плодовый период (с конца второго - начала третьего месяца беременности до рождения).Эмбриотоксического и тератогенного * эффектов, столь выраженных и типичных для предыдущий стадий, в этот период не наблюдается. Исключение составляют лишь аномалии развития половых органов у плодов женского пола, так как половые органы начинают формироваться в относительно поздний период (12 –14 неделя эмбрионального развития). Это может произойти при приеме гормональных препаратов, в том числе и оральных контрацептивов:
Эстрогены и эстроген + гестогеносодержащие препараты - наиболее уязвимые для срока беременности от 8 до 17 недель. Результатом их применения может быть развитие опухолей гениталий, анатомических и функциональных их дефектов, а также возможно нарушение полового развития у плода мужского пола, - псевдогермофродитизм.
Прогестероносодержащие пероральные контрацептивы - их воздействие возрастает от начала беременности до 4-х месяцев. Результатом их применения может быть нарушение формирования половых органов у плода и псевдогермафродитизма девочек, а также возможны аномалии конечностей, сердечно-сосудистой системы, желудочно-кишечного тракта.
ТРЕТИЙ КРИТИЧЕСКИЙ ПЕРИОД
В 18-22 недели беременности происходят важнейшие процессы, связанные с формированием активности головного мозга плода, кроветворной системы, выработки важнейших гормонов, рефлекторных реакций. Следовательно, этот период является третьим критическим периодом в его развитии.
Во второй половине беременности происходит становление и созревание важнейших органов и структур (нервной, кроветворной, сердечно-сосудистой). В этот период также происходит снижение чувствительности плода к повреждающим агентам.
Повреждающие факторы могут реализовать свое действие, проникая через плаценту или изменяя ее нормальную проницаемость. Плацента – это уникальный фильтр, который обладает ограниченной проницаемостью, защищая организм плода от множества веществ и токсичных продуктов, попадающих в организм матери. Однако плацента проницаема для большинства лекарственных веществ, применяемых в акушерской практике: закись азота, наркотические анальгетики, витамины, большинство гормонов, сердечных препаратов, антибиотиков, противовоспалительных препаратов и др. Лекарственные вещества, принятые беременной женщиной, проникая в организм плода, проходят через плаценту. Количество вещества, поступившего в организм плода, зависит от состояния плаценты, ее кровотока, физико-химических особенностей лекарственного агента (например, его молекулярной массы), а также от осложнений, которые могут возникнуть во время беременности (инфицирование, гестоз, употребление алкоголя, резус-конфликт и др.). При возникновении осложнений беременности барьерная функция плаценты нарушается, и через нее начинают проникать даже те вещества, которые в нормальных физиологических условиях через нее не проходят.
ПЛОД В ОТДЕЛЬНЫЕ МЕСЯЦЫ ВНУТРИУТРОБНОГО РАЗВИТИЯ
В течение первого месяца происходит имплантация, образование зародыша, зачатков важнейших органов, зародышевых оболочек.
В конце второго месяца длина плода - 3 –3,5 см, он имеет сформировавшееся тело, зачатки конечностей, его головка равна длине туловища, на ней видны зачатки глаз, носа, рта. В этот период начинают формироваться половые органы.
В конце третьего месяца длина плода составляет 8-9 см, он имеет массу 20-25 г, его головка крупная, уже становится заметно различие в наружных половых органах, конечности совершают движения, видны пальчики на ручках и ножках.
В конце четвертого месяца беременности плод имеет длину 16 см и массу 110-120 г, в это время формируется лицо, происходит окостенение черепа, заканчивается формирование мышечной системы, движения становятся более активными, но беременной еще не воспринимаются. В этот период пол вашего малыша можно ясно различить.
В конце пятого месяца длина плода – 25-26 см, масса 280-300 г. Плод имеет красную кожу, покрытую пушковыми волосами. Начинают функционировать сальные железы, секрет которых участвует в образовании смазки плода. В кишечнике образуется меконий (первородный кал). Женщина начинает воспринимать движения плода. Первобеременная воспринимает движения с 20 недели, повторнобеременная немного раньше – с 18 недели.
Становится возможным выслушивание сердцебиения плода через брюшную стенку (18-20 недель), их частота значительно больше, чем у взрослого человека - 120-140 ударов в минуту.
В конце шестого месяца беременности длина плода - 30 см, масса 600–680 г, его движения становятся еще более активными. Плод уже может дышать (во внутриутробном периоде развития плода происходят нерегулярные дыхательные движения – так ваш малыш готовится к будущему внеутробному дыханию, но эти дыхательные движения не сопоставимы с внеутробным: легкие плода не расширяются, голосовая щель закрыта).
С шестого-седьмого месяца беременности начинают функционировать почки плода, у него образуется моча, которая выделяется в околоплодные воды.
В конце седьмого месяца (28 недель беременности) длина плода -35 см, масса 1000-1200 г. Подкожный жир плода развит слабо, кожа покрыта смазкой и пушковым волосом. Ушные и носовые хрящи очень мягкие, ногти недоразвиты. Однако плод на этом этапе развития рождается живым.
После окончания седьмого месяца внутриутробного развития плод считается недоношенным, но жизнеспособным; ребенку, родившемуся в эти сроки необходима квалифицированная медицинская помощь и тщательный уход.
В конце восьмого месяца беременности длина плода 40–42 см, масса - 1500 – 1700г.
В конце девятого месяца внутриутробного развития длина плода равна 45 –48 см, масса 2400 – 2500 г, подкожно-жировой слой увеличивается, кожа плода становится гладкой, розовой, уменьшается количество пушковых волос на теле, удлиняются волосы на его головке. Плод, родившийся в этот срок, способен к внеутробной жизни, он громко кричит, открывает глаза, у него выражен сосательный рефлекс.
К концу десятого акушерского месяца признаки недоношенности исчезают, плод рождается зрелым.
При неблагоприятных условиях развития, которые могут сопровождать беременность, у доношенного ребенка могут быть признаки незрелости. Иногда может наблюдаться противоположное явление: ребенок рождается раньше срока, но зрелым.